


Abstract
Cefadroxil is a cephalosporin antibiotic used in the treatment of infection. However, cerebrospinal fluid (CSF) concentrations of cefadroxil and other aminocephalosporins are not adequate for the treatment of bacterial meningitis. To evaluate the relevance of PEPT2 in affecting the exposure of aminocephalosporins in brain, we investigated the transport properties of cefadroxil at the blood-CSF interface using primary-cultured epithelial cells and isolated whole tissues of choroid plexus. Our results indicated that cefadroxil was preferentially taken up from the apical as opposed to basal side of the monolayer (5-fold), and its apical uptake was stimulated by an inwardly directed proton gradient. The concentration-dependent apical uptake of cefadroxil was characterized by a high-affinity/low-capacity transport system (Km = 39.0 ± 22.7 μM; Vmax = 22.9 ± 6.6 pmol/mg/min) and a nonsaturable component (Kd = 0.15 ± 0.01 μl/mg/min); in contrast, only a nonsaturable component was found for the basal uptake of cefadroxil (Kd = 0.14 ± 0.01 μl/mg/min). The apical-to-basal transepithelial transport of 2 μM cefadroxil was greater than its basal-to-apical transport, but no differences were observed in directionality when 5 mM concentrations of cefadroxil were studied. Moreover, the cellular efflux of cefadroxil was not saturable in either direction (i.e., to apical or basal side). Finally, no differences were observed in the choroid plexus tissue efflux of 2 μM cefadroxil from wild-type and PEPT2 null mice. These findings demonstrate that PEPT2 has an important role in limiting the exposure of cefadroxil in CSF. Located at the apical membrane of choroid plexus epithelium, PEPT2 acts in a unidirectional (as opposed to bidirectional) manner in transporting cefadroxil from CSF into the cell.
β-Lactam antibiotics, first identified with the discovery of penicillin in 1928, have enjoyed tremendous success as an important class of therapeutic agents for the treatment of bacterial infection (Petri, 2001). Although very useful in infections elsewhere in the body, some penicillin and cephalosporin drugs are disappointing in their ability to treat infections of the central nervous system (CNS). Because CNS infections are very common, especially in children, and associated with high mortality and morbidity, they are an increasingly worldwide challenge (Durand et al., 1993; Nau et al., 1998; Lutsar and Friedland, 2000). Bacterial meningitis and other CNS infections are extremely difficult to treat pharmacologically because of either the unavailability of antibacterial agents or an insufficient concentration of the therapeutic agent in brain (Nau et al., 1998; Lutsar and Friedland, 2000). While pursuing a new generation of antibiotics with wider antibacterial spectrum, scientists have also paid more attention to the mechanisms causing subtherapeutic levels of antibiotics in purulent cerebrospinal fluid (CSF). Low concentrations of β-lactam antibiotics in CSF are often presumed to be the result of poor blood-brain barrier (BBB) and blood-CSF barrier (BCSFB) penetration. However, most β-lactam antibiotics show reasonable paracellular penetration across barriers, especially in patients with meningitis (Nau et al., 1998; Lutsar and Friedland, 2000). Therefore, other mechanism(s) need to be identified to explain this phenomenon. Fishman (1966) suggested that a saturable, probenecid-sensitive system transported penicillin from CSF. Spector and Lorenzo (1974) extended these observations and demonstrated that this active efflux pump was located in the choroid plexus and was saturable at high penicillin concentrations. A saturable efflux mechanism was later confirmed by studies using benzylpenicillin and other β-lactam antibiotics (Spector, 1986; Suzuki et al., 1989; Ogawa et al., 1994).
Proton-coupled oligopeptide transporters (POTs) are a family of membrane-bound proteins that transport a variety of dipeptides and tripeptides and structurally related compounds (Daniel and Kottra, 2004; Smith et al., 2004). All POT isoforms use a proton gradient to cotransport substrates and are a part of the solute carrier superfamily of proteins. Four functional POTs have been identified in humans (i.e., PEPT1, PEPT2, PHT1, and PHT2) and range in size from 572 to 729 amino acids. Complementary DNAs for rat POT isoform proteins have also been characterized and show 80 to 90% homology with their human counterparts. The POTs can be divided into two different subgroups based on their ability to recognize l-histidine as a substrate (PEPT1 and PEPT2 versus the peptide/histidine transporters PHT1 and PHT2). Not surprisingly, the various POTs show remarkable differences in their tissue distribution, substrate affinities, and proposed physiological functions.
It seems that PEPT2 may play a role in the elimination of neuropeptides and peptide fragments from CSF. Functional studies using isolated tissue and primary cultured epithelial cells of choroid plexus demonstrated that neuropeptides such as carnosine (Teuscher et al., 2001, 2004) and glycylglutamine (Hu et al., 2005) were transported in a proton-dependent manner and displayed PEPT2-mediated transport kinetics (i.e., high affinity and low capacity). Being present at the apical membrane of choroidal epithelia (Shen et al., 2004), PEPT2 also functions as an efflux pump for the removal of endogenous or exogenous peptide-like chemicals (Novotny et al., 2000; Kuroda et al., 2005). In contrast, neither PEPT2 mRNA (Berger and Hediger, 1999) nor protein (Shen et al., 2004) is expressed in cerebral endothelial cells of the rat BBB. These findings suggest that apical expression (and activity) of PEPT2 at the BCSFB, as opposed to BBB, is the key determinant of peptide/mimetic persistence in the CNS. Recently, PEPT2-deficient mice have been developed (Rubio-Aliaga et al., 2003; Shen et al., 2003) to further probe the physiological and pharmacological significance of PEPT2. Using transgenic animals, PEPT2 null mice had an impaired uptake of dipeptide/mimetic substrates into choroid plexus compared with wild-type controls, and PEPT2 was found to be the primary POT responsible for trafficking of peptides/mimetics at the BCSFB (Shen et al., 2003; Ocheltree et al., 2004a,b; Teuscher et al., 2004). However, those studies did not directly address the sidedness and directionality of uptake, especially as related to the transepithelial transport of aminocephalosporins at the apical (CSF-facing) and basolateral (blood-facing) membranes of choroid plexus epithelium.
With this in mind, we examined the POT-mediated transport characteristics of an aminocephalosporin, cefadroxil, at the blood-CSF interface using rat choroid plexus epithelial cells in primary culture. We also investigated the uptake of cefadroxil in rat choroid plexus whole tissue as well as the efflux of cefadroxil from choroid plexus tissue in PEPT2-deficient and PEPT2-competent mice. Taken as a whole, our findings are unique in demonstrating, for the first time, that PEPT2 acts as an efflux transporter in the removal of cefadroxil from CSF and that the process is unidirectional.
Materials and Methods
Materials. [3H]Cefadroxil (1 Ci/mmol) and [14C]mannitol (53 mCi/mmol) were purchased from Moravek Biochemicals (Brea, CA). Radiochemical purity of each compound was >99.5%, as reported by the manufacturer. Potential inhibitors [i.e., glycylsarcosine (GlySar); glycine; sarcosine; l-histidine; carnosine; glycylglutamine (GlyGln); kyotorphin; cefadroxil; cephalexin; cephalothin; p-aminohippurate (PAH); and tetraethylammonium (TEA)] were obtained from Sigma-Aldrich (St. Louis, MO). All other chemicals were obtained from standard sources. Cell culture reagents including Dulbecco's modified Eagle's medium/Ham's F-12 medium (1:1) and fetal calf serum were purchased from Invitrogen (Carlsbad, CA). Cells were cultured in the 12-well Transwell-Clear filter systems (12 mm in diameter, 0.4-μm pore size, 1.0 cm2) from Fisher Scientific (Chicago, IL). The transepithelial electrical resistance was determined using a Millicell-ERS device (Millipore Corporation, Billerica, MA). Female Sprague-Dawley rats at day 17 gestation and male Sprague-Dawley rats (100–125 g) were purchased from Charles River Laboratories (Portage, MI). PEPT2–/– mice were generated as described previously (Shen et al., 2003) and backcrossed against the C57BL/6 mouse (The Jackson Laboratory, Bar Harbor, ME). Animals were maintained in a temperature-controlled environment with a 12-h light 12-h dark cycle and given ad libitum access to food and water. All studies were carried out in accordance with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the National Institutes of Health.
Preparation and Culture of Choroid Plexus Epithelial Cells. Primary cultures of epithelial cells from 1- or 2-day-old rat choroid plexuses were prepared as described previously by Shu et al. (2002). After a series of isolation and enrichment steps, the cells were seeded on laminin-coated permeable membrane inserts having a diameter of 12 mm and a pore size of 0.4 μm. Cells were grown at 37°C with 5% CO2 in Dulbecco's modified Eagle's medium/Ham's F-12 medium (1:1) containing: 15 mM HEPES, 10% (v/v) fetal calf serum, 2 mM l-glutamine, 5 μg/ml insulin, 5 μg/ml transferrin, 2 μg/ml hydrocortisone, 5 ng/ml selenium, 10 ng/ml epidermal growth factor, 5 ng/ml basic fibroblast growth factor, 100 U/ml penicillin and streptomycin, and 25 μg/ml gentamicin. The medium was changed every other day. After 12 to 14 days, cell growth reached confluence. Only filters with a transepithelial electrical resistance of at least 150 Ω · cm2 were used for further experiments.
Studies in Choroid Plexus Epithelial Cell Monolayers. Cells were treated with normal Na+-containing artificial cerebrospinal fluid (aCSF: 127 mM NaCl, 20 mM NaHCO3, 2.4 mM KCl, 0.5 mM KH2PO4, 1.1 mM CaCl2, 0.85 mM MgCl2, 0.5 mM Na2SO4, and 5.0 mM glucose, pH 7.4) so that the concentrations of ions and glucose mimicked the physiological microenvironment of CSF. The intracellular uptake, transepithelial transport, and cellular efflux of cefadroxil were determined, as described previously by our laboratory (Shu et al., 2002; Teuscher et al., 2004; Hu et al., 2005). Cellular uptake and efflux data were corrected for the extracellular content, as estimated by mannitol. Transepithelial transport data are presented without correction for paracellular flux.
In brief, cell monolayers were washed with aCSF buffer and preincubated apically and basolaterally for 10 min. The buffer was then removed, and fresh uptake buffer containing [3H]cefadroxil and [14C]mannitol, with and without inhibitors, was applied to the apical (0.4 ml) or basolateral side (1.2 ml) of the monolayer; control buffer (no cefadroxil or mannitol) was added to the opposite side. Incubation proceeded for the designated periods of time at 37°C. To measure transepithelial transport, a 100-μl aliquot of buffer was taken from the receiver compartment and the radioactivity was counted. To measure intracellular uptake, the media were removed and the cell monolayers were washed three times with ice-cold buffer to terminate the uptake process. Filters were then excised from the inserts, and the cells were solubilized with 0.5 ml of 0.2 M NaOH and 1% SDS. After the addition of 6 ml of scintillation cocktail [EcoLite(+); MP Biomedicals, Irvine, CA], radioactivity was measured on a dual-channel liquid scintillation counter (Beckman LS 6000; Beckman Coulter, Fullerton, CA). Cellular protein content was determined by the method of Bradford (1976) using a protein assay kit (Bio-Rad, Hercules, CA) with bovine serum albumin as the standard.
To measure cellular efflux, cells were first incubated with [3H]cefadroxil and [14C]mannitol from the apical side of the monolayer for 60 min. Afterward, the buffer was removed and the cells were washed five times with warm buffer. Fresh aCSF buffer was then added to the apical (0.4 ml) and basolateral (1.2 ml) chambers. At the designated times, 100-μl aliquots were taken from both compartments and replaced with control buffer. Radioactive drug was measured in each chamber by liquid scintillation counting. At the final collection time, cell monolayers were washed and removed and then measured for remaining radioactivity. Efflux was expressed as percentage of the initial substrate in cells after loading.
Studies in Choroid Plexus Whole Tissue. Cefadroxil uptake studies were performed in isolated choroid plexus from rat, whereas cefadroxil efflux studies were performed in isolated choroid plexus from transgenic mice. In the first set of experiments, lateral ventricle choroid plexuses were obtained from male Sprague-Dawley rats (30–50 days old) as described previously (Teuscher et al., 2000, 2001). In brief, tissues were weighed and then placed in aCSF buffer that was continuously bubbled with 5% CO2, 95% O2 at 37°C. After a 5-min equilibration period, the plexuses were transferred to 0.95 ml of fresh aCSF buffer for 0.5 min. Uptake was then initiated by adding 50 μl of buffer containing [3H]cefadroxil and [14C]mannitol. Cefadroxil uptake was terminated by transferring the choroid plexuses to ice-cold buffer and filtering under reduced pressure. The filters were washed three times, and the plexuses were soaked in 0.33 ml of 1 M hyamine hydroxide (a tissue solubilizer) for 30 min before the addition of scintillation cocktail (Cytoscint; MP Biomedicals). Radioactivity was then measured by liquid scintillation counting. Cefadroxil uptake data were corrected for filter binding and for its extracellular content as estimated by mannitol.
In the second set of experiments, lateral and fourth ventricle choroid plexuses were obtained from PEPT2+/+ (wild-type) and PEPT2–/– (null) mice (6–8-week-old littermates) as described previously (Shen et al., 2003; Ocheltree et al., 2004a,b; Teuscher et al., 2004). The mouse choroid plexuses were treated in a manner similar to that described above for choroid plexuses obtained from rat. However, in these experiments, plexuses were incubated with aCSF buffer containing [3H]cefadroxil and [14C]mannitol for 120 min. The tissues were washed five times with warm buffer and then placed into 1 ml of fresh aCSF buffer. At designated times, 100-μl aliquots of buffer were taken and replaced with control buffer. Radioactive drug was measured in the buffer by liquid scintillation counting. At the final collection time, tissues were washed and removed and then measured for remaining radioactivity. Efflux was expressed as percentage of the initial substrate in choroid plexuses after loading.
Stability Studies. Choroid plexus cell monolayers were incubated apically with aCSF buffer containing 1 mM [3H]cefadroxil for 5, 15, 30, and 60 min. At the end of incubation, media from both the apical and basolateral compartments were collected for analysis. Cell monolayers were washed four times with ice-cold buffer, the filters were detached, and 1 ml of ice-cold Milli-Q water (Millipore Corporation) was added before sonication for 10 min. Cell lysates were then mixed with an equal volume of acetonitrile followed by centrifugation at 14,000 rpm for 10 min at 4°C. Choroid plexus tissues (5 and 60 min) were prepared in a similar manner, except that smaller volumes (approximately 0.2 ml each) of water and acetonitrile were added to the tissues. Samples were kept on ice and analyzed within a few hours by high-performance liquid chromatography (Model 510 Pump; Waters, Milford, MA) with radiochromatographic detection (FLO-ONE 500TR; PerkinElmer Life and Analytical Sciences, Boston, MA). [3H]Cefadroxil was analyzed using a reversed-phase column (Hypersil ODS, C-18, 5 μm, 250 × 4.6 mm; Alltech Associates, Deerfield, IL) and a mobile phase of 6% acetonitrile in water, pumped isocratically at 1 ml/min. Under ambient conditions, cefadroxil had a retention time of 4.2 min.
Data Analysis. The concentration-dependent uptake of cefadroxil was best fit to an equation containing a Michaelis-Menten term along with a nonsaturable component: cefadroxil uptake = Vmax × S/(Km + S) + Kd × S, where Vmax is the maximal rate of saturable uptake, Km is the Michaelis-Menten constant, Kd is the clearance for nonsaturable processes, and S is the substrate concentration. To confirm the presence of a single transport system, a Woolf-Augustinsson-Hofstee analysis was performed: V = Vmax – Km × V/S, where V is the saturable uptake component (i.e., V = cefadroxil uptake – Kd × S).
Data are reported as mean ± S.E. using a minimum of three independent experiments. In cell culture studies, each experiment was run in triplicate. In isolated choroid plexus studies, each experiment consisted of two plexuses from one rat or six plexuses from two mice. Statistical comparisons were performed using one-way analysis of variance, and post hoc pairwise comparisons were made using Dunnett's test (Prism; GraphPad Software Inc., San Diego, CA). A probability of p ≤ 0.05 was considered statistically significant. Linear and nonlinear regression analyses were performed with a weighting factor of unity (SigmaPlot 8.0; SPSS Inc., Chicago, IL). The quality of fit was determined by evaluating the coefficient of determination (r2) and standard error of parameter estimates and by visual inspection of the residuals.
Results
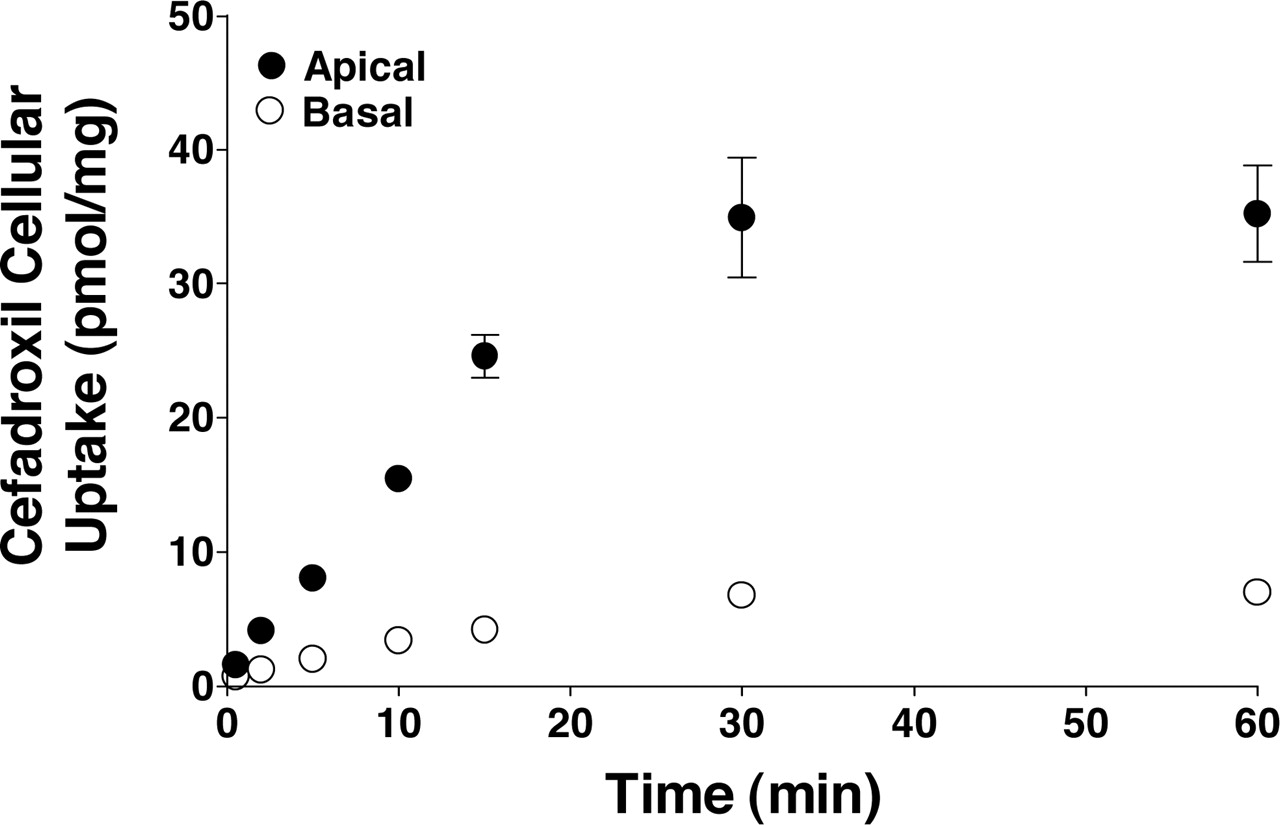
Time Course of Cefadroxil Uptake in Rat Choroid Plexus Epithelial Cell Monolayers. As shown in Fig. 1, the uptake of cefadroxil was substantially greater when introduced from the apical than from the basolateral surface of cell monolayers (∼5-fold). A linear uptake rate was observed over 15 min and, as a result, initial rates were determined at 5 min for subsequent cell culture experiments. When introduced apically, cefadroxil reached a plateau value of 35 pmol/mg protein at 60 min. Given its medium concentration of 2 μM and assuming a cellular volume of 3.66 μl/mg protein (Blais et al., 1987), the intracellular to extracellular concentration ratio of cefadroxil was 4.8, suggesting the presence of an active uptake process. In contrast, the intracellular to extracellular concentration ratio of cefadroxil was 1.0 when introduced basolaterally.
Concentration-Dependent Uptake of Cefadroxil in Rat Choroid Plexus Epithelial Cell Monolayers. To probe the concentration dependence of uptake for cefadroxil from the apical and basal sides of the cell monolayer, initial rates were measured over a wide range of buffer concentrations (i.e., apical, 1.25–500 μM; basolateral, 5–10,000 μM). As observed in Fig. 2A, the apical uptake of cefadroxil occurred by both saturable and nonsaturable processes, in which Vmax = 22.9 ± 6.6 pmol/mg protein/min, Km = 39.0 ± 22.7 μM, and Kd = 0.15 ± 0.01 μl/mg protein/min (r2 = 0.974). Moreover, a single transport system was involved for the saturable component, as suggested by a linear slope of the Woolf-Augustinsson-Hofstee plot (r2 = 0.998). In contrast, the basolateral uptake of cefadroxil occurred only by a nonsaturable component, in which the slope of the line was linear out to 10 mM (r2 = 0.917; Fig. 2B). Here, the nonsaturable uptake clearance (Kd) was 0.14 ± 0.01 μl/mg protein/min, a value very close to the one observed after cefadroxil uptake from the apical side.

Time course of 2 μM cefadroxil uptake in rat choroid plexus cell monolayers (pH 7.4, both chambers). Uptake was linear for 15 min from both the apical and basolateral sides. All data were mannitol-corrected and expressed as mean ± S.E. (n = 4–5).

Concentration-dependent uptake of cefadroxil in rat choroid plexus cell monolayers (pH 7.4, both chambers). Initial-rate uptakes were determined at 5 min for cefadroxil concentrations ranging from 1.25 to 500 μM (apical) and 5 to 10,000 μM (basolateral). Apical uptake data were fitted by nonlinear regression in which Vmax = 22.9 pmol/mg protein/min, Km = 39.0 μM, and Kd = 0.15 μl/mg protein/min (r2 = 0.974; A). The inset of A shows a Woolf-Augustinsson-Hofstee plot (V, picomoles per milligram per minute versus V/[cefadroxil], microliters per milligram per minute), which is linear (r2 = 0.998). Basolateral uptake data were fitted by linear regression in which Kd = 0.14 μl/mg protein/min (r2 = 0.917; B). All data were mannitol-corrected and expressed as mean ± S.E. (n = 4–6).
Time- and Concentration-Dependent Uptake of Cefadroxil in Rat Choroid Plexus Whole Tissue. As shown in Fig. 3A, the tissue uptake of cefadroxil was linear for up to 2 min of incubation and reached a plateau value of 2.2 pmol/mg tissue at 60 min. Given its medium concentration of 0.2 μM and assuming a water content of 0.787 μl/mg tissue (Ennis and Keep, 2005), the tissue water-to-extracellular concentration ratio of cefadroxil was 14.0. Concentration-dependent analyses (0.5–500 μM), based on 1-min uptakes, revealed that cefadroxil accumulated in choroid plexus tissue by both saturable (Vmax = 34.0 ± 3.8 pmol/mg tissue/min; Km = 17.1 ± 5.4 μM) and nonsaturable (Kd = 0.13 ± 0.01 μl/mg tissue/min) processes (r2 = 0.974; Fig. 3B). As suggested by the Woolf-Augustinsson-Hofstee plot, the saturable component involved a single transport system (r2 = 0.960). Thus, the experiments in choroid plexus whole tissue were similar to those in choroid plexus epithelial cells in primary culture with respect to saturability and transporter affinity and the ability of cefadroxil to accumulate in choroid plexus against a concentration gradient.

Time course of 0.2 μM cefadroxil uptake in rat choroid plexus whole tissue (buffer pH 7.4), where uptake was linear for up to 2 min (A). The concentration-dependent uptake of cefadroxil was then evaluated using initial-rate uptakes of 1 min and cefadroxil buffer concentrations of 0.5 to 500 μM (B). Saturable and nonsaturable components of the curve were determined by nonlinear regression in which Vmax = 34.0 pmol/mg tissue/min, Km = 17.1 μM, and Kd = 0.13 μl/mg tissue/min (r2 = 0.974; B). The inset of B shows a Woolf-Augustinsson-Hofstee plot (V, picomoles per milligram per minute versus V/[cefadroxil], microliters per milligram per minute), which is linear (r2 = 0.960). All data were mannitol-corrected and expressed as mean ± S.E. (n = 5–6).
Effect of pH and Inhibitors on Apical Uptake of Cefadroxil in Rat Choroid Plexus Epithelial Cell Monolayers. To determine whether the apical uptake of cefadroxil was stimulated by an inwardly directed proton gradient, we investigated cellular accumulation of the drug at pH 7.4 and 6.5 (using aCSF buffer, adjusted with dropwise addition of HCl). As shown in Fig. 4A, the uptake of cefadroxil was increased 2-fold at the lower pH, a finding consistent with PEPT2 being functional active on the apical membrane of our cell culture preparations. Specificity of the PEPT2-mediated uptake of cefadroxil was examined using a wide range of potential inhibitors. As shown in Fig. 4B, the apical uptake of cefadroxil was reduced by 75 to 85% when studied in the presence of a model dipeptide (i.e., GlySar), several neuropeptides (i.e., carnosine, GlyGln, and kyotorphin), and two aminocephalosporins (i.e., cefadroxil and cephalexin). In contrast, cefadroxil uptake was unaffected by amino acids (i.e., glycine, sarcosine, and l-histidine, the latter being a substrate of the peptide/histidine transporters PHT1 and PHT2), a cephalosporin lacking an α-amino group (i.e., cephalothin), and prototypical organic anionic and cationic substrates (i.e., PAH and TEA).

Effect of pH on the 5-min apical uptake of 2 μM cefadroxil in rat choroid plexus cell monolayers (apical chamber at pH 7.4 or 6.5 and basolateral chamber at pH 7.4; A). ★ ★ ★, p < 0.001 compared with pH 7.4 values. Effect of potential inhibitors (5 mM) on the 5-min apical uptake of 2 μM cefadroxil in rat choroid plexus cell monolayers (both chambers at pH 7.4; B). ★ ★, p < 0.01 compared with control values. All data were mannitol-corrected and expressed as mean ± S.E. (n = 7–8).
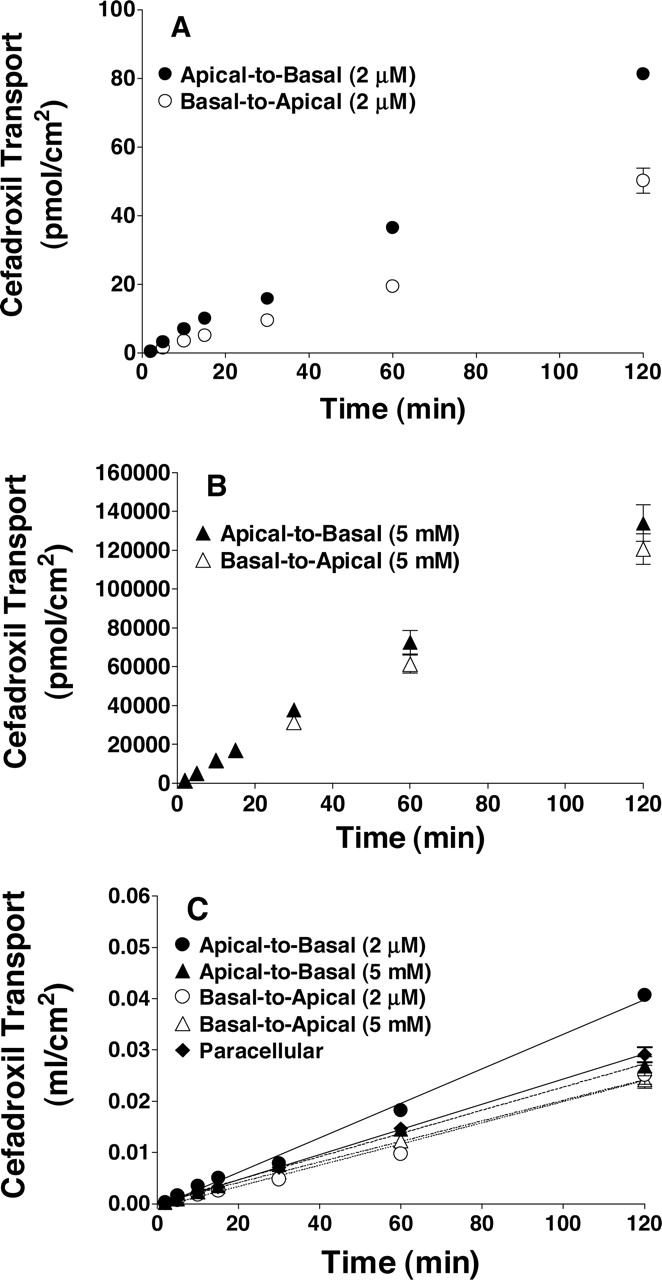
Transepithelial Transport of Cefadroxil in Rat Choroid Plexus Epithelial Cell Monolayers. As shown in Fig. 5A, the apical-to-basal transepithelial transport of 2 μM cefadroxil was greater than its basal-to-apical transport. However, no difference in directionality was observed when cefadroxil was studied at 5 mM concentrations (Fig. 5B). When cefadroxil was adjusted for differences in the buffer concentration of the drug (i.e., expressed as a transport volume in Fig. 5C; milliliters per square centimeter), the slopes of the lines (which represent permeability) were essentially the same for the 5 mM concentrations of cefadroxil in either direction as well as the basal-to-apical transport of 2 μM cefadroxil. Moreover, these three slopes were not distinguishable from that of mannitol. Thus, when saturation occurs at the apical surface, consistent with PEPT2 being present on this membrane, diffusional permeability is the same in both the apical-to-basal and basal-to-apical directions and indistinguishable from that of paracellular flux.

Time course for the transepithelial transport of cefadroxil across rat choroid plexus cell monolayers (pH 7.4, both chambers). Studies were performed using either 2 μM cefadroxil (A) or 5 mM cefadroxil (B). Transepithelial data (in A and B) were also transformed by dividing the transported amount of cefadroxil by its concentration in medium and plotted in panel C (milliliters per square centimeter), where the slopes of the lines represent the permeabilities (centimeters per minute) of the monolayers. Linear regression analyses indicated that the apical-to-basal permeability of 2 μM cefadroxil was significantly different from the other curves (p < 0.001). The other three slopes for cefadroxil permeability were not significantly different from one another, nor distinguishable from that of paracellular flux, as estimated by mannitol (p > 0.50). The apical-to-basal and basal-to-apical slopes of mannitol (5 mM) were combined, because polarity was not evident (p > 0.50). All data were expressed as mean ± S.E. (n = 4–6).
Cellular Efflux of Cefadroxil in Rat Choroid Plexus Epithelial Cell Monolayers and Choroid Plexus Whole Tissue from Transgenic Mice. Having established that choroid plexus epithelial cells preferentially accumulated and transported cefadroxil in the apical-to-basal direction, the cellular efflux kinetics of cefadroxil were examined along with the role of PEPT2 in this process. As shown in Fig. 6, A and B, the initial efflux rate of cefadroxil was significantly faster to the apical as opposed to basal side of the cell monolayer (at all time points up to 60 min), yet both sides reached the same equilibrium value within 90 to 120 min of incubation. In addition, the cellular efflux pattern of cefadroxil was similar at 2 μM and 1 mM concentrations of drug. This relationship is best displayed in Fig. 6C, where cefadroxil was rapidly effluxed from the cells with no difference in slope (i.e., first-order half-lives of 18.3 and 19.3 min at 2 μM and 1 mM, respectively). These results suggest that the cellular efflux of cefadroxil from choroid plexus was not saturable and that PEPT2 was not functioning in a bidirectional manner. We subsequently evaluated the efflux of 2 μM cefadroxil from choroid plexus whole tissue of PEPT2+/+ and PEPT2–/– mice. As shown in Fig. 7A, no differences were observed between genotypes in the time course of efflux from tissue to buffer. Moreover, the tissue efflux rate was the same (i.e., first-order half-lives of 31.5 and 25.2 min for wild-type and PEPT2 null mice, respectively; Fig. 7B), confirming that PEPT2 was not involved in the efflux of cefadroxil from choroid plexus.
Stability of Cefadroxil. Greater than 97% cefadroxil was recovered intact from samples collected from either the donor, receiver, or intracellular compartment of the cell culture experiments and from tissue samples of choroid plexus. Therefore, cefadroxil instability was not a confounding issue in these studies, and no further correction of the data was performed.
Discussion
In the present study, several new findings were revealed with respect to the trafficking of cefadroxil at the BCSFB. First, cefadroxil had a preferential apical uptake and apical-to-basal transepithelial transport across choroid plexus epithelial cells. Second, the apical uptake of cefadroxil was saturable and of high affinity, whereas the basolateral uptake of cefadroxil was a nonsaturable process. Third, the choroid plexus efflux of cefadroxil was neither saturable nor mediated by PEPT2. Given its apical expression in choroid plexus epithelium, the results demonstrate that PEPT2 acts in a unidirectional (as opposed to bidirectional) manner in transporting cefadroxil into the cell.
A major challenge in the treatment of CNS infections, including meningitis, is the effective delivery of antibacterial compounds across the BBB and BCSFB into brain (Nau et al., 1998). Even though some β-lactam antibiotics can enter the brain parenchyma via “leaky” barriers produced by meningeal inflammation (Norrby, 1978), various transporters expressed in the BBB and BCSFB can minimize the effective penetration of antibiotics by eliminating them from brain and/or CSF into the blood (Sun et al., 2003; Kusuhara and Sugiyama, 2004). In particular, it has been demonstrated in transgenic mice that PEPT2 accounts for 80 to 85% of cefadroxil uptake in isolated choroid plexus, with a minor contribution by organic anion transporters (OATs) (Ocheltree et al., 2004a). Although the apical uptake and apical-to-basal transport of GlySar (Shu et al., 2002) and GlyGln (Hu et al., 2005) were clearly preferred in choroid plexus epithelial cells in primary culture, no such information existed regarding the sidedness and directionality of cefadroxil at the BCSFB (Ocheltree et al., 2004a). Results from our study demonstrate that, like GlySar and GlyGln, cefadroxil has a preferred transcellular efflux from CSF to blood. If this preference is sustained in vivo, then PEPT2 would serve to reduce cefadroxil exposure in CSF, thereby limiting its effectiveness (and perhaps that of other β-lactam antibiotics) in the treatment of bacterial infections of the brain.
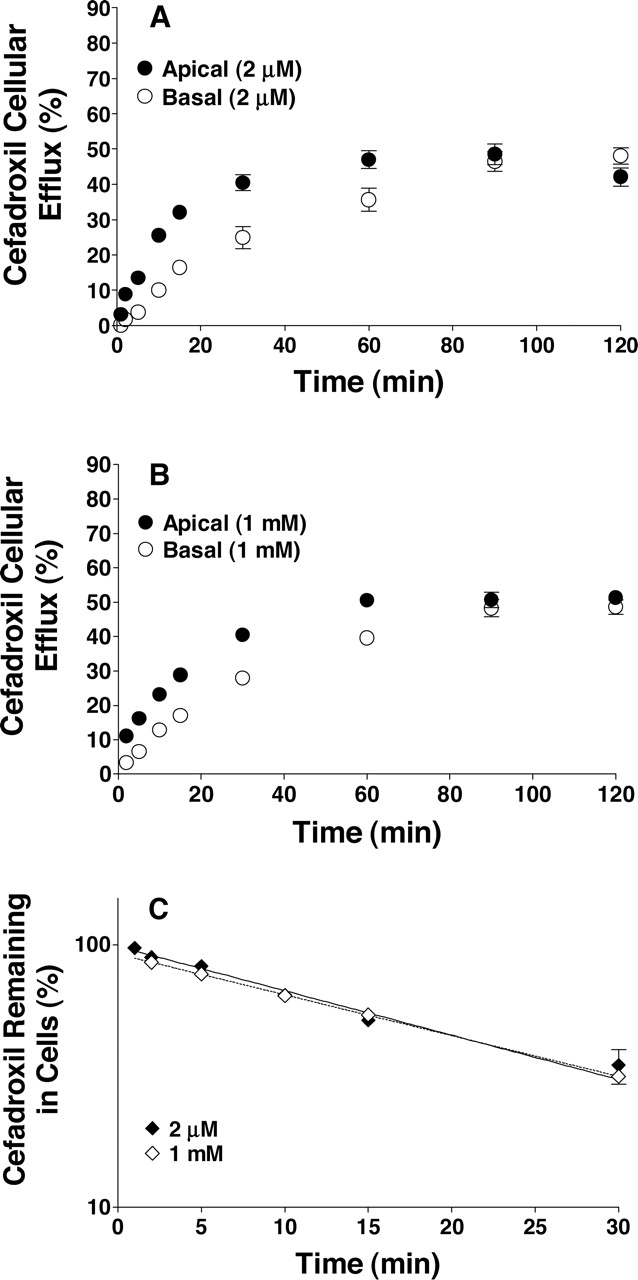
Time course of cefadroxil efflux from rat choroid plexus cell monolayers (pH 7.4, both chambers), where cells were preloaded for 60 min with either 2 μM cefadroxil (A) or 1 mM cefadroxil (B). Efflux into the apical and basolateral chambers was reported as percentage of the initial cefadroxil loaded in cells. The percentage of cefadroxil remaining in cells as a function of time was also analyzed (C). Logarithmic-linear regression indicated that the two lines were not significantly different (p = 0.5917 for slope; p = 0.3858 for y-intercept). All data were mannitol-corrected and expressed as mean ± S.E. (n = 6).

Time course of cefadroxil efflux from choroid plexus whole tissue of transgenic mice, where plexuses from wild-type and PEPT2 null mice were preloaded for 120 min with 2 μM cefadroxil (A). The percent of cefadroxil remaining in tissues as a function of time was also analyzed (B). Logarithmic-linear regression indicated that the two lines were not significantly different (p = 0.1648 for slope; p = 0.1556 for y-intercept). All data were mannitol-corrected and expressed as mean ± S.E. (n = 3–4).
Kinetic analyses support the PEPT2-mediated apical uptake of cefadroxil in choroid plexus. High-affinity interactions observed for cefadroxil in these rat cell culture and whole tissue studies (Km values of 39 and 17 μM, respectively) were comparable with values reported for cefadroxil uptake in rat renal brush-border membrane vesicles (Km = 9 μM; Ries et al., 1994), PEPT2 cRNA-transfected Xenopus laevis oocytes (Km = 32 μM; Boll et al., 1996), and isolated choroid plexus from PEPT2+/+ mice (Km = 27 μM; Ocheltree et al., 2004a). Previous immunoblot and immunolocalization studies further demonstrated that, although PEPT2 is localized to the apical membrane of choroid plexus epithelium, PEPT1 was not present in the choroid plexus (Novotny et al., 2000; Shu et al., 2002; Shen et al., 2004). In addition, even though PHT1 and PHT2 transcripts have been detected in rat brain (Yamashita et al., 1997; Sakata et al., 2001), including PHT1 expression in choroid plexus (Yamashita et al., 1997), the lack of inhibition of cefadroxil uptake by l-histidine in this study argues against either PHT1 or PHT2 being expressed on the apical membrane. Surprisingly, the present studies showed only a 7% inhibition of cefadroxil uptake by PAH (not statistically significant) in rat choroid plexus cell cultures (Fig. 4B) or isolated whole tissue (data not shown). In contrast, our previous studies in transgenic mice showed a 10 to 15% inhibition of cefadroxil uptake by PAH in choroid plexus tissue (Ocheltree et al., 2004a). Regardless of the species (i.e., mouse versus rat) or preparation used (i.e., primary cell cultures versus whole tissue), the choroid plexus uptake of cefadroxil by OATs was a minor process.
By comparing the Vmax/Km and Kd values for the apical uptake of cefadroxil in choroid plexus cell cultures, one can estimate that 80% of the total uptake occurred by PEPT2 and 20% of the total uptake occurred by nonsaturable processes. This finding agrees with the inhibition studies, where a residual activity of ∼20% for cefadroxil would indicate complete saturation of the PEPT2 transporter. In comparing the Vmax values for cefadroxil uptake in rat cell cultures (23 pmol/mg protein/min) versus whole tissue (equivalent to 227 pmol/mg protein/min; assuming 6.7 mg of tissue/mg protein), it is also evident that apical PEPT2 is down-regulated by approximately 10-fold in choroid plexus epithelial cells in primary culture compared with freshly isolated tissue. Interestingly, the nonsaturable uptake of cefadroxil was very similar across the apical and basolateral membranes (i.e., 0.14–0.15 μl/mg of protein/min) as was its nonsaturable transepithelial permeability (i.e., apical-to-basal direction at 5 mM concentrations and basal-to-apical direction at 2 μMor 5 mM concentrations, as shown in Fig. 5C), indicating that cefadroxil has equal access into and across the cells in the absence of PEPT2 activity. These results suggest a nonspecific mechanism for cefadroxil uptake, such as simple diffusion or transport by a channel across the choroid plexus epithelial membranes, along with paracellular transport between the cells. However, in the presence of PEPT2 protein, cefadroxil exposure in CSF would be restricted by the transporter's unidirectional uptake of drug at the apical membrane of choroid plexus epithelium, the rate-determining step in its transcellular efflux. A model that accounts for these observations is shown in Fig. 8.

Proposed model for cefadroxil trafficking at the BCSFB, where CP is the choroid plexus epithelium, NS is the nonspecific uptake/efflux, PC is the paracellular transport, PEPT2 is the specific oligopeptide transporter (Slc15a2), and TJ is the tight junction.
One must use caution in extrapolating these results with cefadroxil to other aminocephalosporin antibiotic drugs. As reported by Kuroda et al. (2005), cefaclor was rapidly eliminated from rat CSF by a benzylpenicillin-sensitive mechanism distinct from OAT3, whereas cefalexin was removed by a GlySar-sensitive transporter, presumably PEPT2. Thus, cefaclor and cefalexin do not share the same uptake mechanism in choroid plexus, even though both drugs are aminocephalosporins differing only by a single functional group in the dihydrothiazine ring (3-chloro for cefaclor; 3-methyl for cefalexin). This example highlights the importance of defining the key substrate recognition sites for specific membrane transporters in the brain. In this regard, the minimal structural and chemical prerequisites for PEPT2 recognition and transport include a free N terminus, a correctly positioned backbone carbonyl group, and a carboxylic group that is suitably distanced from the intramolecular carbonyl function and the amino terminal head group (Theis et al., 2002). Understanding the structural and chemical requirements of peptidomimetic substrates for PEPT2 will be critical for future drug development and targeting strategies.
Although there is an increasing number of studies examining the mechanisms of how peptides/peptidomimetics are taken up by cells, there is much less known about how such compounds are effluxed from cells. The current study indicates that, in cultured choroid plexus epithelial cells, such efflux occurs via a nonsaturable mechanism (e.g., possibly diffusion). There is some evidence from giant patch-clamp experiments that oligopeptide transporters might act as bidirectional transporters (Kottra and Daniel, 2001), but in our study, we found no difference in dipeptide efflux between PEPT2+/+ and PEPT2–/– mice.
In conclusion, our results are unique in demonstrating that PEPT2 functions in the unidirectional transport of an aminocephalosporin drug, cefadroxil, from CSF into choroid plexus epithelium. This finding provides the mechanistic basis for low levels of cefadroxil in CSF (Ocheltree et al., 2004a) and furthers our understanding of drug efflux transporters and their role in affecting the CNS exposure and pharmacological response to drugs. Future studies will be directed at studying the in vivo pharmacokinetics and tissue distribution of cefadroxil in PEPT2-competent and PEPT2-deficient mice.
Footnotes
-
This study was supported in part by Grants R01 GM035498 (to D.E.S.) and R01 NS034709 and P01 HL018575 (to R.F.K.) from the National Institutes of Health.
-
doi:10.1124/jpet.105.090654.
-
ABBREVIATIONS: CNS, central nervous system; CSF, cerebrospinal fluid; BBB, blood-brain barrier; BCSFB, blood-cerebrospinal fluid barrier; POT, proton-coupled oligopeptide transporter; GlySar, glycylsarcosine; GlyGln, glycylglutamine; PAH, p-aminohippurate; aCSF, artificial cerebrospinal fluid; OAT, organic anion transporter; TEA, tetraethylammonium.
- Received June 6, 2005.
- Accepted August 15, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}