


Abstract
Aryl- (SULT1A1), estrogen- (SULT1E1), and hydroxysteroid- (SULT2A1) sulfotransferases (SULTs) are active determinants of xenobiotic detoxication and hormone metabolism in the adult human liver. To investigate the role of these conjugating enzymes in the developing human liver, the ontogeny of immunoreactive SULT1A1, SULT1E1, and SULT2A1 expression was characterized in a series of 235 pre- and postnatal human liver cytosols ranging in age from early gestation to a postnatal age of 18 years. Interindividual variability in expression levels was apparent for all three SULTs in pre- and postnatal liver samples. Expression of the three SULTs displayed distinctly different developmental profiles. Semiquantitative Western blot analyses indicated that SULT1A1 and SULT2A1 immunoreactive protein levels were readily detectable in the majority of developmental human liver cytosols throughout the prenatal period. Whereas SULT1A1 expression did not differ significantly among the various developmental stages, SULT2A1 expression increased during the third trimester of gestation and continued to increase during postnatal life. By contrast, SULT1E1, a cardinal estrogen-inactivating enzyme, achieved the highest levels of expression during the earliest periods of gestation in prenatal male livers, indicating a requisite role for estrogen inactivation in the developing male. The present analysis suggests that divergent regulatory mechanisms are responsible for the differential patterns of hepatic SULT1A1, SULT1E1, and SULT2A1 immunoreactive protein levels that occur during pre- and postnatal human development, and implicates a major role for sulfotransferase expression in the developing fetus.
In the adult human liver, aryl- (SULT1A1), estrogen- (SULT1E1), and hydroxysteroid- (SULT2A1) sulfotransferases are particularly important determinants of drug and hormone metabolism. The human hepatic sulfotransferase-conjugating enzymes catalyze the transfer of a sulfonate moiety from the physiological sulfonate donor, 3′-phosphoadenosine-5′-phosphosulfate, to an appropriate xenobiotic substrate or endogenous metabolic intermediate (Jakoby et al., 1980). In most cases, xenobiotic sulfonation is a detoxication mechanism that reduces the lipophilicity of target substrates and results in the formation of polar end products that are more amenable to excretion.
Human SULT1A1 is expressed in hepatic and extrahepatic adult tissues and metabolizes a broad spectrum of phenolic substrates, including pharmaceuticals such as troglitazone (Honma et al., 2002) and minoxidil (Falany and Kerl, 1990), carcinogenic dietary heterocyclic amines such as 2-amino-3-methyl-9H-pyrido [2,3-b]indole (Glatt et al., 2004), and estradiol at supraphysiological concentrations (Falany, 1997). SULT2A1 is robustly expressed in adult human liver and catalyzes the sulfonation of the sex hormone precursor, DHEA, as well as monohydroxy bile acids and other alcohol and hydroxysteroid substrates (Radominska et al., 1990; Falany et al., 1994, 1995a). SULT1E1 is primarily responsible for catalyzing the sulfonation of estrogens at nanomolar concentrations and is considered to be the principal sulfotransferase involved in the inactivation of estrogens under physiological conditions (Falany et al., 1995b).
SULT1A1 (Vietri et al., 2001), SULT2A1 (Barker et al., 1994), and SULT1E1 (Miki et al., 2002) enzyme activities have been detected previously in a limited number of fetal liver samples. Several sulfotransferases, including SULT1A1, catecholamine sulfotransferase, SULT2A1, and SULT1E1, are also expressed in the growing placenta (Kerry et al., 2001). In the developing human liver, SULT1A1 has been reported to be comparatively more sensitive than its adult counterpart to the inhibitory effects of salicylic acid (Vietri et al., 2001), suggesting that significant disparities between fetal and adult sulfotransferase activities may have an impact on transplacental drug detoxication and hormone metabolism.
In the maternal-fetal unit, the sulfotransferases are poised to work in concert to deflect transplacental drug toxicity and to modulate bioactive hormone levels. For example, hepatic SULT2A1 is strategically positioned to cooperate with SULT1E1 in the control of estrogen homeostasis in the developing fetus. In this regard, formation of the abundant circulating steroid, DHEA sulfate, is catalyzed by SULT2A1 in fetal liver and adrenal tissues. Then, through the action of placental sulfatases, the DHEA that is released provides the placenta with the primary source for estrogen biosynthesis (Barker et al., 1994).
During human embryogenesis, a well integrated network of sulfotransferase expression has crucial implications for the health of the maternal-fetal unit. To improve our understanding of this important metabolic dynamic, the present study examines the age- and gender-related alterations in SULT1A1, SULT2A1, and SULT1E1 immunoreactive protein content in a spectrum of pre- and postnatal human liver cytosols.
Materials and Methods
Materials. A panel of 235 pre- and postnatal human liver cytosol samples was obtained from several sources, including the Brain and Tissue Bank for Developmental Disorders, University of Maryland at Baltimore and the University of Miami (National Institute for Child Health and Human Development, N01-HD-8-3283 and N01-HD-8-3284, respectively), and the Central Laboratory for Human Embryology, University of Washington, Seattle, WA (National Institute of Child Health and Human Development, HD-00836). Donor characteristics and a description of the preparation of subcellular fractions have been published previously (Koukouritaki et al., 2002). Supernatant fractions from the first high-speed differential centrifugation step were used in the current study. Polyclonal rabbit anti-human SULT1A1 antibody was developed and characterized previously (Falany et al., 1990). Polyclonal rabbit anti-human SULT1E1 and rabbit anti-human SULT2A1 antibodies were purchased from Pan-Vera Corp. (Madison, WI), as were standards consisting of insect cell cytosols containing baculovirus-expressed SULT1A1, SULT1E1, or SULT2A1. Horseradish peroxidase-conjugated goat anti-rabbit IgG was obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Prestained BenchMark protein standards were purchased from Invitrogen (Carlsbad, CA). ECL Plus Western Blotting Detection Reagents and Hybond-P membranes were purchased from GE Healthcare (Piscataway, NJ). The Criterion Western blotting system was purchased from Bio-Rad Laboratories (Hercules, CA).
Western Blot Analysis of Human SULT1A1, SULT1E1, and SULT2A1 in Pre- and Postnatal Liver Cytosols. Enhanced chemiluminescence Western blotting procedures were performed as described previously (Duanmu et al., 2002), with the indicated modifications. Human liver cytosolic proteins (7.5–20 μg of protein per lane) were fractionated on precast 10% Tris-HCl Criterion electrophoresis gels (Bio-Rad) and transferred onto Hybond-P membranes (GE Healthcare), using the Criterion Western blotting system (Bio-Rad), according to manufacturer's instructions. Each Western blot included four to five decremental amounts of insect cell cytosolic protein containing baculovirus-expressed human SULT1A1 (1.98–4950 ng of cytosolic protein), SULT2A1 (4.50–1350 ng of cytosolic protein), or SULT1E1 (14.3–1710 ng of cytosolic protein) for the purpose of constructing a reference standard curve. Uniform protein loading and transfer were verified for each Western blot using Ponceau S staining, as described previously (Fang et al., 2005a). For the detection of immunoreactive SULT1A1, primary antibody was used at a dilution of 1:20,000, and secondary antibody (horseradish peroxidase-conjugated goat anti-rabbit IgG) was used at 1:30,000. Both SULT1E1 and SULT2A1 primary antibodies were used at a dilution of 1:5000, and secondary antibody was used at 1:20,000. Primary antibody incubations were performed overnight at 4°C, whereas secondary antibody incubations were conducted at room temperature for 3 h. Western blots were developed using ECL Plus Western Blotting Detection Reagents, according to the manufacturer's instructions (Amersham). Chemiluminescence signals were digitally captured for image quantification using an Image Station 440CF and 1D Image Analysis Software (Eastman Kodak Co., Rochester, NY).
Sulfotransferase Enzyme Activities. After Western blot analysis, 10 human liver cytosols from the developmental panel (three prenatal, four early postnatal, and three postnatal samples) were analyzed for correlative form-selective SULT activities, using prototypical substrates. SULT1A1 activity was assayed using p-nitrophenol as substrate in a thin-layer chromatography-based assay, as described previously (Falany, 2005). Dehydroepiandrosterone sulfonation was used to determine SULT2A1 activity (Falany et al., 1989), and nanomolar concentrations of β-estradiol were used to assay SULT1E1 activity selectively (Falany et al., 1995b). All assays were performed in triplicate, and product formation did not exceed 15% of the substrate in the reaction.
Data Analysis. After data capture from an individual blot, a standard curve relating band intensity to amount of protein standard (i.e., amount of insect cytosolic protein containing one of the baculovirus-expressed sulfotransferases) was prepared by fitting a least-squares line to the data corresponding to the standard samples. A standard curve r2 value of 0.9 was set as the minimal limit for acceptance of a data set. The intensity of each enhanced chemiluminescence band representing SULT1A1, SULT1E1, or SULT2A1 in the human liver cytosol samples was then expressed as nanograms of SULT standard (i.e., nanograms of insect cell cytosolic protein containing baculovirus-expressed human SULT) per microgram of human liver cytosolic protein. As indicated in the figure legends, the data were grouped according to age and, in selected cases, gender. Scatter and box and whisker plots of the data were prepared using Prism, version 4.02 (GraphPad Software, San Diego, CA). Statistical comparisons among three groups were performed using the Kruskal-Wallis nonparametric ANOVA and Dunn's multiple comparison test, and two-group comparisons were performed using the Mann-Whitney U test (Prism). Correlation analyses between SULT immunoreactive protein levels and enzyme activities were also performed using Prism. In all cases, p < 0.05 was accepted as significant.
Results
Western blot analyses of immunoreactive SULT1A1, SULT2A1, and SULT1E1 protein levels in human pre- and postnatal liver cytosols demonstrated the presence of all three sulfotransferases throughout the developmental periods examined. For analysis, the protein expression data were initially separated according to subject age into three groups: prenatal (8–42 weeks), early postnatal (birth to 12 months), and postnatal (>1–18 years). SULT2A1 expression in the individual samples during this progression is shown in Fig. 1 (A and B). The data indicated the presence of considerable interindividual variability in the amount of SULT2A1 among samples. For example, in the postnatal (>1–18 years) samples, the 25th and 75th percentile values of SULT2A1 protein content differed by more than 40-fold (7.6 and 312.2 ng SULT2A1/μg cytosolic protein, respectively). Nevertheless, SULT2A1 expression was found to increase significantly after the birth transition (Fig. 1C), with the median SULT2A1 protein contents in the prenatal and early postnatal samples determined to be 7.4 and 178.4 ng SULT2A1/μg cytosolic protein, respectively. Also, relative to the prenatal interval, significantly greater levels of SULT2A1 expression were maintained in the postnatal (>1–18 years) human liver (median 109.5 ng SULT2A1/μg cytosolic protein). When the prenatal samples were evaluated according to trimester, the increase in SULT2A1 immunoreactive protein levels was observed to begin during the third trimester (Fig. 1D). In contrast to the rat counterpart of SULT2A1, which demonstrates both an age- and a gender-dependent pattern of expression (Runge-Morris and Wilusz, 1991), the levels of human hepatic SULT2A1 during development were not differentially expressed with respect to gender (data not shown).

Developmental expression of human hepatic SULT2A1 immunoreactive protein. A, representative Western blot showing the relative amounts of SULT2A1 immunoreactive protein in a random subset of human liver cytosols from the developmental panel. B, SULT2A1 immunoreactive protein levels in individual human liver cytosol samples are shown plotted against subject age, divided into three age groups: prenatal (a), early postnatal (birth to 12 months; b), and postnatal (>1–18 years; c). The number of samples in each group is indicated, and the data are presented as nanograms of SULT2A1 standard per microgram of human liver cytosolic protein. EGA, estimated gestational age; PNA, postnatal age. C, summary statistics are shown for SULT2A1 protein content in the prenatal and postnatal samples. D, summary statistics are shown for SULT2A1 protein content in the prenatal samples according to trimester. For each box and whisker plot, the line bisecting the box is the median value, the upper and lower limits of the box are the 25th and 75th percentile values, and the error bars (whiskers) extend to the lowest and highest values (note the broken y-axis). Statistical comparisons, performed using the Kruskal-Wallis nonparametric ANOVA followed by Dunn's multiple comparison test, are indicated by the lines above the plots. **, p < 0.01; ***, p < 0.001; ns, not significant.
Like SULT2A1, hepatic immunoreactive SULT1A1 protein was detected from early gestation, remained present throughout the pre- and postnatal periods, and displayed considerable interindividual variability in levels (Fig. 2, A and B). For example, one subject in the postnatal age group (>1–18 years) displayed a hepatic SULT1A1 immunoreactive protein level that was >10-fold higher than those measured in the majority of pre- and postnatal liver samples (Fig. 2B). Also like human hepatic SULT2A1, the developmental expression of human hepatic SULT1A1 did not display gender-related differences (data not shown). In contrast to SULT2A1, SULT1A1 protein expression did not differ significantly among the prenatal and postnatal sample groups (Fig. 2C).
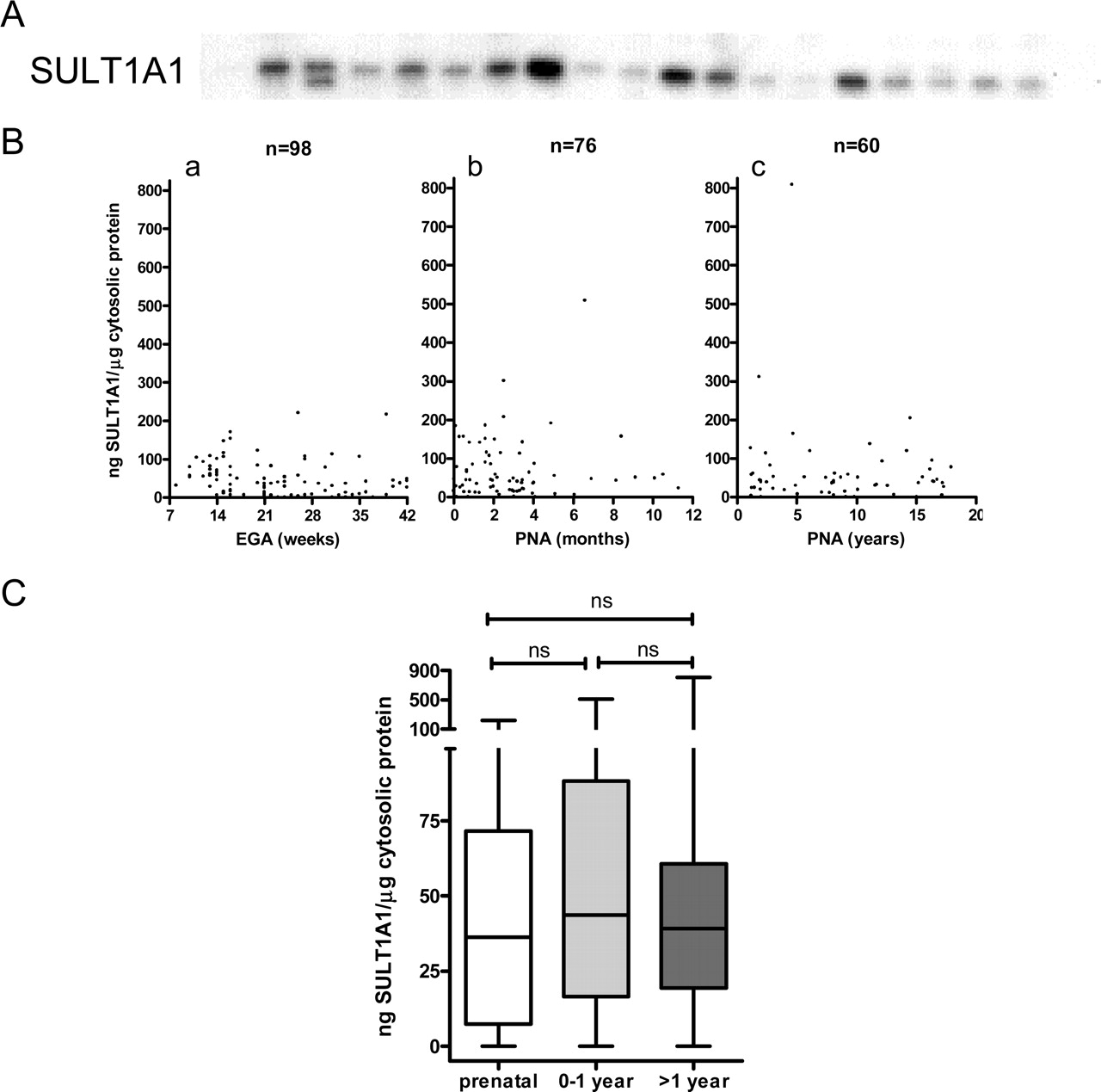
Developmental expression of human hepatic SULT1A1 immunoreactive protein. A, representative Western blot showing the relative amounts of SULT1A1 immunoreactive protein in a random subset of human liver cytosols from the developmental panel. B, SULT1A1 immunoreactive protein levels in individual human liver cytosol samples are shown plotted against subject age, divided into three age groups: prenatal (a), early postnatal (birth to 12 months; b) and postnatal (>1–18 years; c). The number of samples in each group is indicated, and the data are presented as nanograms of SULT1A1 standard per microgram of human liver cytosolic protein. EGA, estimated gestational age; PNA, postnatal age. C, summary statistics are shown for SULT1A1 protein content in the samples of each age group. For each box and whisker plot, the line bisecting the box is the median value, the upper and lower limits of the box are the 25th and 75th percentile values, and the error bars (whiskers) extend to the lowest and highest values (note the broken y-axis). Statistical comparisons, performed using the Kruskal-Wallis nonparametric ANOVA followed by Dunn's multiple comparison test, are indicated by the lines above the plots. ns, not significant.
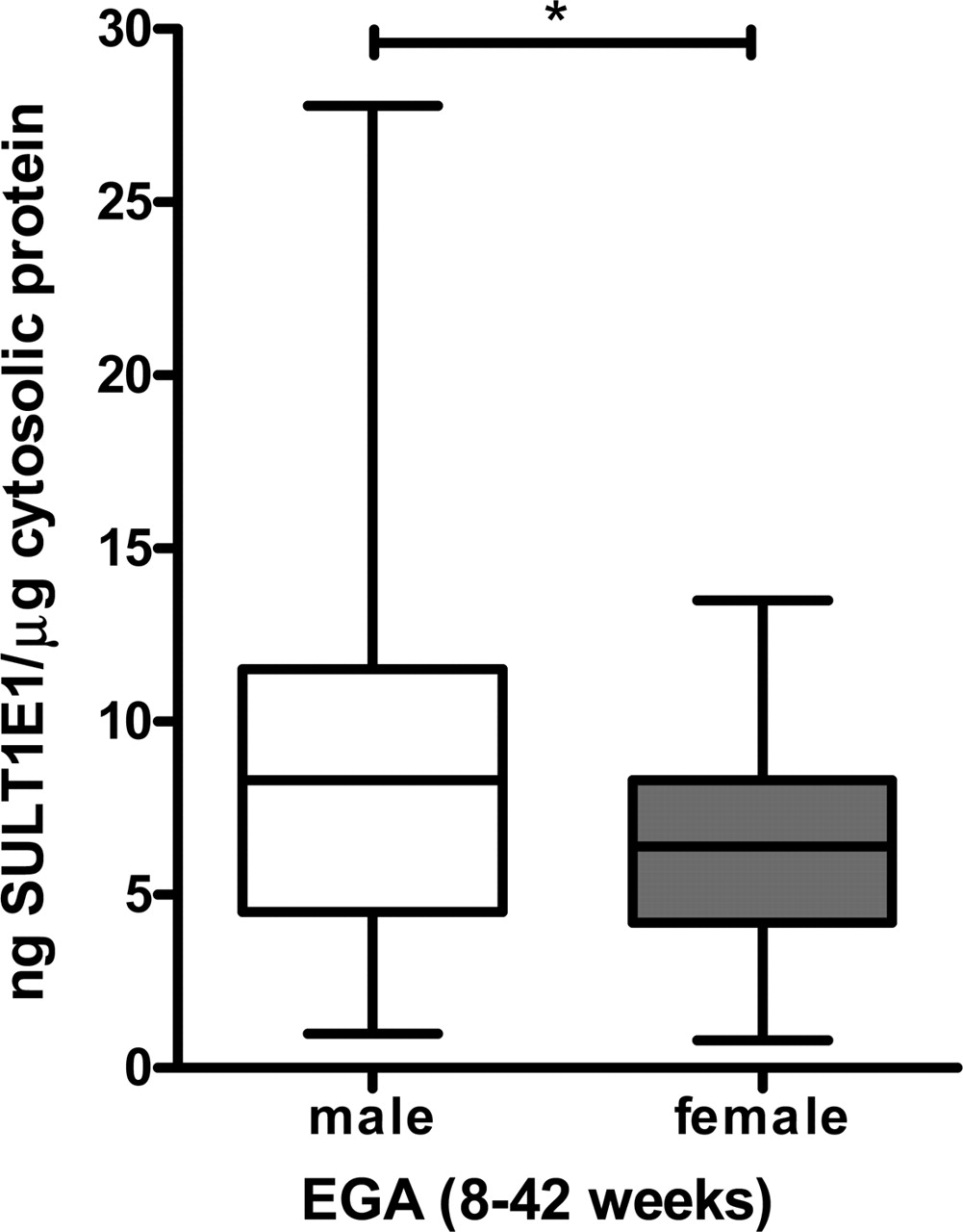
In sharp distinction to the patterns of SULT2A1 and SULT1A1 developmental expression, human hepatic SULT1E1 levels were greatest during the prenatal period and declined sharply thereafter (Fig. 3B). A significant decline in the immunoreactive expression of human hepatic SULT1E1 occurred from the prenatal period (8–42 weeks, median 7.8 ng SULT1E1/μg cytosolic protein) to the first year of life (median 4.2 ng SULT1E1/μg cytosolic protein), and again from the first postnatal year to childhood (>1–18 years, median 2.3 ng SULT1E1/μg cytosolic protein) (Fig. 3C). Upon closer scrutiny, the strongest prenatal expression of human hepatic SULT1E1 was observed to occur during the first and second trimesters, and declined significantly thereafter (Fig. 3D). Further analysis revealed that the trend toward high levels of prenatal hepatic SULT1E1 expression segregated most consistently with males (median of 8.3 ng SULT1E1/μg cytosolic protein in males versus median of 6.4 ng SULT1E1/μg cytosolic protein in females) (Fig. 4).

Developmental expression of human hepatic SULT1E1 immunoreactive protein. A, representative Western blot showing the relative amounts of SULT1E1 immunoreactive protein in a random subset of human liver cytosols from the developmental panel. B, SULT1E1 immunoreactive protein levels in individual human liver cytosol samples are shown plotted against subject age, divided into three age groups: prenatal (a), early postnatal (birth to 12 months; b) and postnatal (>1–18 years; c). The number of samples in each group is indicated, and the data are presented as nanograms of SULT1E1 standard per microgram of human liver cytosolic protein. EGA, estimated gestational age; PNA, postnatal age. C, summary statistics are shown for SULT1E1 protein content in the samples of each age group. D, summary statistics are shown for SULT2A1 protein content in the prenatal samples according to trimester. For each box and whisker plot, the line bisecting the box is the median value, the upper and lower limits of the box are the 25th and 75th percentile values, and the error bars (whiskers) extend to the lowest and highest values (note the broken y-axis). Statistical comparisons, performed using the Kruskal-Wallis nonparametric ANOVA followed by Dunn's multiple comparison test, are indicated by the lines above the plots. *, p < 0.05; ***, p < 0.001; ns, not significant.
To determine the extent to which SULT expression, as determined by Western blot analysis, was indicative of concomitant alterations in SULT enzyme activity, 10 human liver cytosols from the developmental panel (three prenatal, four early postnatal, and three postnatal samples) were assessed for SULT enzyme activities using substrates that have been commonly regarded as selective probes for SULT1A1 (p-nitrophenol), SULT2A1 (dehydroepiandrosterone), and SULT1E1 (β-estradiol) (Falany et al., 1989, 1995b; Falany, 2005). For SULT2A1 and SULT1E1, enzyme activities significantly correlated positively with immunoreactive protein contents (Fig. 5). Although p-nitrophenol sulfonation activity did not correlate positively with SULT1A1 protein level (Fig. 5), this finding is consistent with a recent report indicating that p-nitrophenol is actually not a suitable probe substrate for the determination of SULT1A1 activity in human liver cytosols, because p-nitrophenol is substantially metabolized by additional SULTs that are present in human liver, such as SULT1B1 (Tabrett and Coughtrie, 2003). In that report, there was no correlation between SULT1A1 protein content and maximal p-nitrophenol sulfonation activity in a panel of 68 liver cytosols (Tabrett and Coughtrie, 2003).
Discussion
The cellular composition of fetal liver is a dynamic mixture of hematopoietic and hepatobiliary precursors (Barker et al., 1994). Hepatocytes predominate in the adult human liver, and toxic substrates such as bile acids may be detoxified by either sulfonation or UDP glucuronidation pathways (Hofmann, 1990). In fetal life, however, UDP-glucuronosyltransferase transcripts are not readily detected (Strassburg et al., 2002), suggesting that the hepatic sulfotransferases play a prevailing role in fetal biotransformation and in the precise control of hormone metabolism during embryogenesis.

Comparison of SULT1E1 immunoreactive protein levels in prenatal male and female samples. Summary statistics are shown for SULT1E1 protein content in the samples from the prenatal male and female subjects [(EGA 8–42 weeks) (EGA, estimated gestational age)]. For each box and whisker plot, the line bisecting the box is the median value, the upper and lower limits of the box are the 25th and 75th percentile values, and the error bars (whiskers) extend to the lowest and highest values. *, p < 0.05 (Mann-Whitney U test).

Correlation analyses of SULT immunoreactive protein levels with enzymatic activities toward probe substrates in human liver cytosols. Ten human hepatic cytosol samples from the developmental panel (three prenatal, four early postnatal, and three postnatal), which had been analyzed for SULT1A1, SULT2A1, and SULT1E1 immunoreactive protein contents, were analyzed for p-nitrophenol (PNP; for SULT1A1; A), dehydroepiandrosterone (DHEA; for SULT2A1; B) and β-estradiol (E2; for SULT1E1; C) sulfonation activities. Each scatter plot shows one of the sets of enzyme activities plotted against the corresponding immunoreactive protein contents, with the least-squares line also shown. The r2 values from the correlation analysis are indicated; *, p < 0.05, **, p < 0.01.
To dissect the roles of hepatic xenobiotic-metabolizing enzymes as fetal defenses against toxic insult, the set of pre- and postnatal human liver samples that provided the foundation for this study was previously characterized with respect to the ontogeny of CYP2C, CYP3A, CYP1E1, and flavin-containing monooxygenase expression. Analogous to the present investigation, interindividual differences and differential patterns of CYP2C9 and CYP2C19 expression were observed in pre- and postnatal human liver samples, suggesting that opposing gene-regulatory factors are temporally recruited during liver development (Koukouritaki et al., 2004). CYP2E1 levels were diminished in prenatal liver samples but were noted to increase steadily with progressive hepatic maturation (Johnsrud et al., 2003). By contrast, the fetal and adult forms of CYP3A (Stevens et al., 2003) and flavin-containing monooxygenase (Koukouritaki et al., 2002) displayed inverse patterns of gene expression, indicating that developmentally sensitive transcription factors signal a switch in isoform-specific gene expression at particular phases of liver development.
In the present study, human hepatic SULT2A1 immunoreactive protein was expressed from early gestation, through the birth transition, and during the postnatal period up to 18 years of age. SULT2A1 levels in the developing human liver were subject to interindividual variations in expression and, consistent with their robust levels of expression in the adult human liver, increased significantly during the perinatal and postnatal periods. SULT2A1 sulfonates the gonadal hormone precursor DHEA and provides the starting material for estrogen biosynthesis in the placenta (Barker et al., 1994). In addition, since SULT2A1 catalyzes the sulfonation of toxic monohydroxy bile acids (Kitada et al., 2003), its continuous expression during fetal and postnatal life may be essential to avoid bile acid stress and the emergence of intrahepatic cholestasis, a life-threatening event in the developing human (Howard and Murphy, 2003).
Sulfotransferases are important determinants of hormone metabolism and bioavailability. Although rat models have been used extensively in the past to investigate thyroid hormone metabolism in developing fetal tissues, iodothyronine sulfotransferases display important species-, tissue-, and gender-dependent differences that are not directly recapitulated in the human. For example, SULT1B1 and SULT1C1 play a major role in iodothyronine sulfonation in rat liver, and the catalytic contribution of rat hepatic SULT1C1 is greater in male relative to female rat liver (Kester et al., 2003). A number of human hepatic SULT enzymes, including the recently cloned human hepatic SULT1B2 (trivial name hST1B2), are capable of sulfonating iodothyronines (Wang et al., 1998; Li and Anderson, 1999; Kester et al., 1999). Among the human sulfotransferases that are catalytically active toward iodothyronines, SULT1A1 displayed a robust catalytic efficiency toward iodothyronine sulfonation in human liver and kidney cytosols (Kester et al., 1999; Richard et al., 2001) and is considered to play a central role in the regulation of receptor-active levels of 3,5,3′-triiodothyronine (T3) in humans (Kester et al., 1999).
The current analysis demonstrated that human hepatic SULT1A1 is expressed at comparable levels in fetal and postnatal human liver. These observations are in accord with previous reports defining SULT1A1 expression in the developing human (Richard et al., 2001). Maintaining a tight harness on the temporal release of the biologically active form of thyroid hormone T3, also is essential to ensure normal brain and nervous system development in the differentiating fetus (Zoeller, 2004). In the adult human liver, the sulfonation of thyroid hormone accelerates its deiodination and consequent irreversible inactivation by D1 deiodothyroninase (Kester et al., 1999). During embryogenesis, D1 levels are relatively low and iodothyronine concentrations are high (Kester et al., 1999). Under these special conditions, the sulfonation of T3 is strategically positioned to serve as a reversible reservoir for the targeted release of bioactive T3 by tissue sulfatases (Kester et al., 1999).
Consistent with a previous study, which detected the presence of hepatic SULT1E1 enzyme activity between 17 and 21 weeks of gestation (Miki et al., 2002), the current investigation demonstrated that the expression of human hepatic SULT1E1 during development occurs early and correlates inversely with maturation. SULT1E1 immunoreactive protein levels were greatest during the initial embryonic period of liver development (first and second trimester, e.g., 8–28 weeks gestational age), and expression was highest in fetuses of the male gender. Endocrine disruption of the male androgen axis during development has been described as one of the multifactorial risk factors in the pathogenesis of hypospadias and testicular dysgenesis (Silver, 2004). It is plausible that the striking levels of hepatic SULT1E1 expression that are evident during early male development are necessary to inactivate biologically available estrogens and to maximize androgen intensity at critical stages of gonadal organogenesis.
Correlative analysis revealed that SULT immunoreactive protein levels were significantly associated with corresponding form-selective catalytic activities in the cases of SULT2A1 and SULT1E1. For SULT1A1, the lack of a positive correlation of immunoreactive protein content with the cytosolic metabolism of p-nitrophenol, a well known SULT1A1 substrate, is consistent with the recent finding of Tabrett and Coughtrie (2003) and is probably attributable to p-nitrophenol metabolism by additional SULT isoforms that are expressed in human liver.
Our results agree closely with an independent investigation delineating the tissue-specific expression of several SULT enzymes in prenatal donors ranging in age from 14 to 18 weeks of gestation (Stanley et al., 2005). In this recent study, SULT1E1 content was detectable in fetal liver, whereas robust levels of SULT1A1 were expressed in fetal liver and a wide variety of extrahepatic tissues (Stanley et al., 2005). SULT2A1 was most abundantly expressed in the fetal adrenal gland, and the levels of SULT2A1 in fetal liver were reduced in comparison to adult liver (Stanley et al., 2005).
Foundation studies in rodent and adult human hepatocytes indicate that nuclear receptor and liver-enriched transcription factors are critical regulators of hepatic SULT gene expression (Runge-Morris et al., 1999; Sonoda et al., 2002; Assem et al., 2004). However, the transcription factor controls that govern sulfotransferase expression in the developing human liver are unknown. Candidates include liver-enriched transcription factors, including those of the hepatocyte nuclear factor family. Hepatocyte nuclear factor transcription factors are instrumental to the process of tissue differentiation during embryogenesis (Landry et al., 1997) and have also been implicated in the regulation of human hepatic CYP2C gene expression (Ibeanu and Goldstein, 1995). Information on the molecular regulation of the human hepatic sulfotransferases is also emerging. Our laboratory reported a role for the peroxisome proliferator-activated receptor α (PPARα) nuclear receptor in the transactivation of human hepatic SULT2A1 transcription (Fang et al., 2005b). Since the PPARα transcription factor was recently demonstrated to regulate human SULT2A1 expression in primary adult human hepatocytes and in HepG2 cells (Fang et al., 2005b), it is reasonable to expect that endogenous activators of PPARα, such as lipid mediators generated during hepatic biogenesis, may influence the temporal shift in SULT2A1 expression that occurs with progressive liver maturation. Similarly, a recent study demonstrated that the human SULT1A1 promoter is synergistically transactivated by Sp1 and Ets family transcription factors, mediators that also figure prominently in cell cycle control and developmental gene regulation (Hempel et al., 2004). Overall, aberrations in the regulatory signals that govern the highly orchestrated pattern of SULT1E1, SULT1A1, and SULT2A1 expression that occurs during pre- and postnatal liver development would be predicted to have serious implications for development. Future investigations in the field will be faced with the challenge of dissecting the complex transcription factor networks that determine the course of metabolic gene expression with advancing liver differentiation during fetal life. The ultimate goal of this research will be to predict the capacity of the pre- and postnatal liver to achieve a correct balance of xenobiotic detoxication, biotransformation of harmful endogenous intermediates, and hormone homeostasis in the developing human.
Footnotes
- Received August 1, 2005.
- Accepted December 7, 2005.
-
This work was conducted with support from National Institutes of Health Grants ES05823 (to M.R.-M.) and GM38953 (to C.N.F.), and by services provided by the Imaging and Cytometry Facility Core of Environmental Health Sciences Center Grant ES06639.
-
doi:10.1124/jpet.105.093633.
-
ABBREVIATIONS: DHEA, dehydroepiandrosterone; SULT, sulfotransferase; T3, 3,5,3′-triiodothyronine; ANOVA, analysis of variance; PPARα, peroxisome proliferator-activated receptor α.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}