


Abstract
The impact of age and sex on the expression of hepatic cytochrome P450 3A4 (CYP3A4) was recently determined in a transgenic mouse line carrying the human CYP3A4 gene. To further investigate the physiological regulation of human CYP3A genes, a novel transgenic mouse line was generated using a bacterial artificial chromosome clone containing both CYP3A4 and CYP3A7 genes. CYP3A7 expression was observed in transgenic mouse fetal livers, whereas CYP3A4 exhibited developmental expression characterized by sexual dimorphism in postpubertal livers. Hepatic CYP3A4 protein and RNA were expressed in immature transgenic male mice and became undetectable after 6 weeks of age, whereas CYP3A4 was expressed in both immature and adult females. CYP3A4 was markedly elevated by the xenobiotic receptor activator phenobarbital in both male and female livers, demonstrating drug induction of the CYP3A4 transgene in this mouse model. Furthermore, continuous infusion of recombinant growth hormone (GH) in transgenic male mice, overriding the pulsatile male plasma GH profile, increased hepatic CYP3A4 mRNA and protein to normal female levels. Continuous GH treatment also feminized the expression of endogenous murine Cyp2b and Cyp3a44 genes. Thus, human CYP3A4 contains all of the gene regulatory sequences required for it to respond to endogenous hormonal regulators of developmental expression and sexual dimorphism, in particular GH. These findings may help elucidate the role of GH in determining the sex-dependent expression of CYP3A4 in human liver and suggest that GH therapy may alter the pharmacokinetic and pharmacodynamic properties of CYP3A4 substrates, leading to enhanced metabolism and disposition of drugs in men.
The human cytochrome P450 3A (CYP3A) gene subfamily consist of four members, CYP3A4, CYP3A5, CYP3A7, and CYP3A43, which are all located on chromosome 7q22.1 (Gellner et al., 2001). The encoded CYP3A7 protein is a fetal-specific form (Komori et al., 1990) that accounts for approximately 50% of the total P450 protein present in fetal liver, whereas CYP3A4 is the most abundant P450 enzyme in mature hepatocytes and small intestinal epithelial cells. CYP3A4 is responsible for the metabolism of more than 50% of marketed drugs, including cholesterol-lowering statins, benzodiazepines, calcium channel blockers, human immunodeficiency virus antivirals, macrolide antibiotics, and physiologically important substrates, such as steroids and bile acids (Guengerich, 1999).
CYP3A4 can be regulated by many endogenous and exogenous factors, leading to considerable interindividual variations in the metabolism of CYP3A4 drug substrates. Nuclear receptor regulation of the CYP3A4 gene accounts in part for some of this diversity (Willson and Kliewer, 2002). Xenobiotic drugs that activate pregnane X receptor (PXR), constitutive androstane receptor (CAR), or vitamin D receptor are known to induce CYP3A4 expression, potentially leading to altered pharmacokinetic profiles of drugs metabolized by this P450 enzyme. In addition, hepatocyte nuclear factor-4α plays an important role in CYP3A4 transcriptional regulation (Tirona et al., 2003).
CYP3A4 expression increases during the development from childhood to adulthood as suggested by a recent comprehensive study (Stevens et al., 2003). Although Cyp3a genes are expressed in a sexually dimorphic manner in mice and rats (Waxman et al., 1988; Kawai et al., 2000; Sakuma et al., 2002), it has not been firmly established whether CYP3A4 expression is sex-dependent in humans (Hines and McCarver, 2002; Gandhi et al., 2004; Parkinson et al., 2004), which has been studied in vitro using human liver preparations and in vivo using CYP3A4 probe drugs. Several lines of evidence indicate that the metabolism of several clinical drugs, mainly oxidized by CYP3A4, is more efficient in women than in men (Greenblatt et al., 1980; Watkins et al., 1989; Gorski et al., 1998). Recent studies show that women have higher hepatic CYP3A4 content and drug-metabolizing capacity than men (Wolbold et al., 2003); however, this conclusion is opposed by other observations (Schmucker et al., 1990; George et al., 1995).
Investigation of the sex specificity of CYP3A4 in humans is complicated by interindividual differences in exposure to dietary supplements, drinking, smoking, and medication history, all of which are known to affect CYP3A4 expression or enzymatic activity. These factors can be controlled in the case of animals maintained under defined dietary and environmental conditions, suggesting that animal models, including transgenic mice, may be useful for investigation of the sexual dimorphism and developmental expression of human P450 enzymes. Extensive investigations carried out in rats and mice, noted above, have demonstrated that Cyp3a expression is sex-dependent in these species, although the sexual dimorphism of certain P450 enzymes in rats may be opposite to that in humans (Rich and Boobis, 1997). In addition, a recent study with a CYP3A4 transgenic mouse line (Granvil et al., 2003) revealed a sexual dimorphism in the developmental expression of the CYP3A4 transgene (Yu et al., 2005).
To further investigate the sexual dimorphism and developmental regulation of human CYP3A genes, a novel transgenic mouse line carrying human CYP3A4 and CYP3A7 was produced with a bacterial artificial chromosome (BAC) clone. Sexual dimorphism of CYP3A4 expression was observed in transgenic mouse livers, whereas CYP3A7 was expressed in fetal livers. Phenobarbital, an activator of the xenobiotic receptors CAR and PXR, markedly induced the expression of the CYP3A4 transgene and of the endogenous murine Cyp3a genes. Because sexually dimorphic growth hormone (GH) secretion is an important determinant of gender-specific patterns of growth and metabolism (Jaffe et al., 1998) and GH treatment markedly alters Cyp3a expression as demonstrated by extensive studies with rats and mice (Waxman et al., 1988; Robertson et al., 1998; Kawai et al., 2000; Sakuma et al., 2002), transgenic adult mice were treated with GH to determine the hormone responsiveness of the CYP3A4 transgene. Our findings show that continuous GH treatment stimulated hepatic CYP3A4 expression in transgenic male mice. These results may provide insight into the physiological regulation of CYP3A4 expression in humans and its potential significance in drug therapy.
Materials and Methods
Chemicals, Enzymes, and Other Reagents. Phenobarbital, 4-amidinophenylmethanesulfonyl fluoride, and disodium ethylenediaminetetraacetate were purchased from Sigma (St. Louis, MO). Recombinant human cytochrome P450 Supersomes and pooled human liver microsomes (coded H161) were purchased from BD Discovery Labware, Inc. (Woburn, MA). Restriction enzymes were purchased from Roche Applied Science, Life Technologies (Gaithersburg, MD), or New England Biolabs (Ipswich, MA) and were used in buffer systems provided by the manufacturers. Mouse antiglyceraldehyde-3-phosphate dehydrogenase (GAPDH) monoclonal antibody was purchased from Chemicon International Inc. (Temecula, CA). Cytochrome P450 immunoblot analysis was carried out using antibodies to human CYP3A7 (Sim et al., 2005) and CYP3A4 (mAb 275-1-2) and rat CYP3A1 (mAb 2-13-1) (Gelboin et al., 1995), CYP1A2 (mAb 22-341) (Park et al., 1982), CYP2A3 (Nagata et al., 1987), and CYP2B1 (Yamano et al., 1989). Highly purified recombinant mouse GH was purchased from the National Hormone and Peptide Program-UCLA (Torrance, CA).
Animals and Treatments. Mice were maintained under controlled temperature (23 ± 1°C) and lighting (lights on 6:00 AM–6:00 PM) with chow and water provided ad libitum. Animal handling was in accordance with the National Institutes of Health guidelines for the care and use of laboratory animals and study protocols approved by the Institutional Animal Care and Use Committee at National Cancer Institute. Phenobarbital sodium salt was dissolved in saline at a concentration of 8 mg/ml. Mice (male or female, 6–7 weeks old, two to three in each group) were administered i.p. with phenobarbital (80 mg/kg) or saline (as control) everyday for 4 days, after which the mice were killed and livers were collected and stored at –80°C. For the GH treatment study, 7-week-old male and female mice were implanted s.c. with an Alzet miniosmotic pump (Durect Corporation, Cupertino, CA) filled with 0.8 μg/μl recombinant mouse GH that was released at a rate of 0.5 μl/h. Mice with surgery but no pump implantation (sham treatment) were used as a control. At least four mice were included in each treatment group. All mice were killed 1 week later, at which point the livers were collected and snap-frozen in liquid nitrogen and then stored at –80°C.
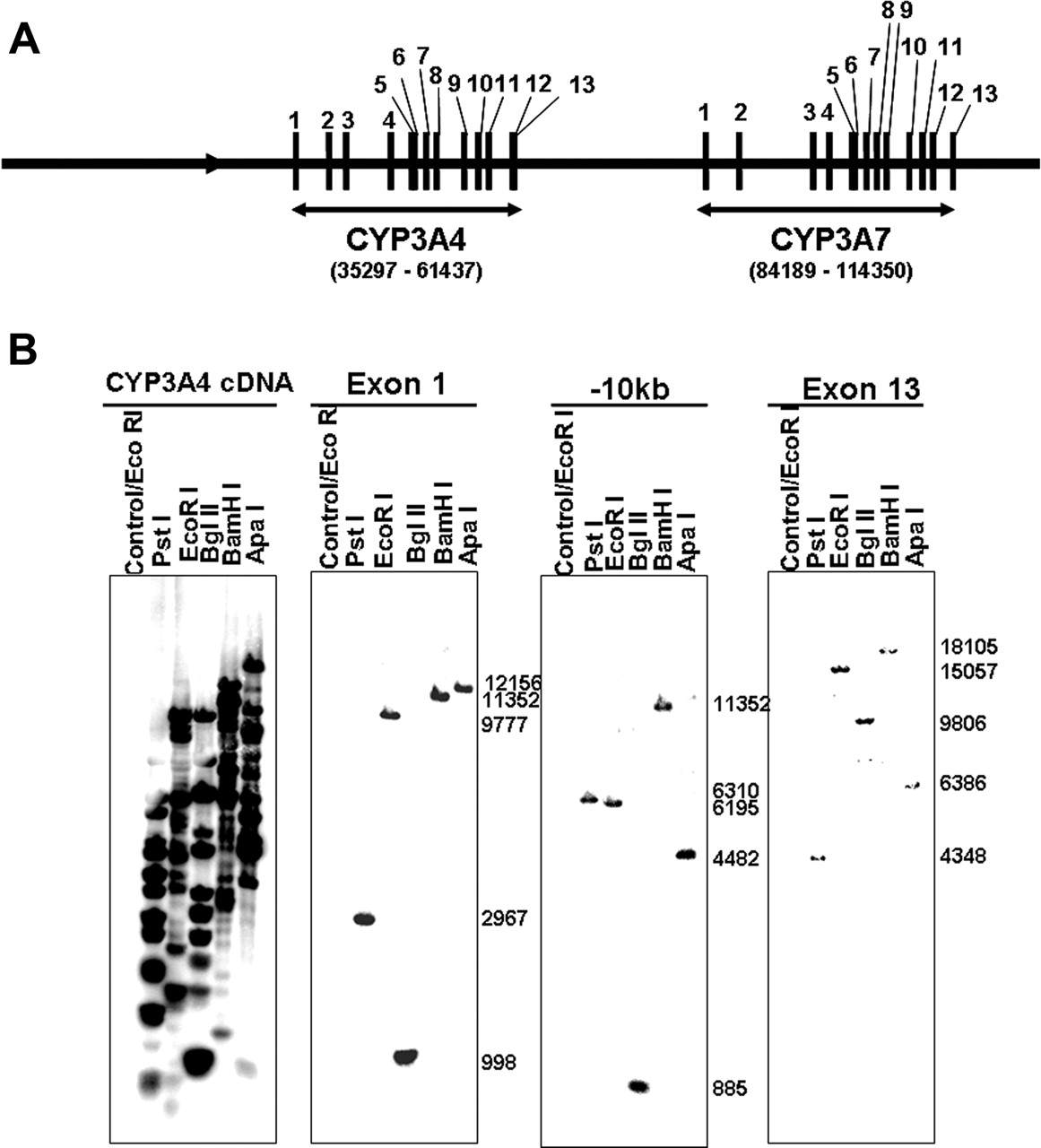
Generation ofCYP3A4-CYP3A7Transgenic Mice. The BAC clone RP11-757A13 (Resgen/Invitrogen Corporation, Huntsville, AL) containing the complete CYP3A4 and CYP3A7 gene sequences, including 5′- and 3′-flanking sequence (Fig. 1A), was identified. The BAC clone was purified by using a maxiprep kit (QIAGEN, Valencia, CA) and verified by Southern blot analysis with 32P-end-labeled CYP3A4 cDNA and DNA oligonucleotide probes recognizing specific regions (exons 1 and 13, –10 kb upstream) of the human CYP3A4 gene (Fig. 1B). The BAC clone was linearized by restriction enzyme digestion (P1-Sce) and purified before microinjection into fertilized FVB/N mouse eggs. Several transgenic founders were identified by Southern blot analysis and bred with C57BL/6 mice. Mice resulting from this breeding step that were positive for the human CYP3A4 transgene by Southern blot analysis were tested for CYP3A4 protein expression by Western blotting. Mice derived from the C1 and C2 founder lines expressed CYP3A4 protein (Fig. 2A). The C2 founder line was selected and further mated to C57BL/6 mice for at least five generations. Homozygous mice were then generated by breeding mice positive for the human CYP3A4 transgene and were designated as CYP3A4 and CYP3A7 transgenic mice. The CYP3A4-negative litter mates obtained from the more than five-generation backcross into the C57BL/6 strain were bred with each other and were used as wild-type controls for subsequent studies.
Southern Blot Analysis. DNA was digested with PstI, EcoRI, BglII, BamHI, or ApaI for the analysis of the BAC clone. Genomic DNA was isolated from mouse tail and digested with BamHI for the determination of individual mouse genotype. Electrophoresis and Southern hybridization conditions described previously were followed (Granvil et al., 2003). Random-primer 32P-labeled DNA oligonucleotide probes recognizing specific regions of the human CYP3A4 gene were used for analysis of the BAC clone. A random-primer 32P-labeled CYP3A4 cDNA (Gonzalez et al., 1988) probe was used for screening mice containing human CYP3A4 and CYP3A7 transgenes.

Verification of the BAC clone used for the production of CYP3A4 and CYP3A7 transgenic mouse line. A, diagram of the wild-type CYP3A4 and CYP3A7 genes in BAC clone (123,778 bp in length) used for microinjection. Vertical black bars, CYP3A4 and CYP3A7 exons. B, Southern blot analyses of the BAC clone after purification and digestion with PstI, EcoRI, BglII, BamHI, or ApaI. Hybridizations were performed by using 32P-end-labeled CYP3A4 cDNA and DNA oligonucleotide probes that recognize exons 1 and 13 and –10 kb upstream sequence of human CYP3A4 gene, respectively. A BAC clone that does not contain the CYP3A4 gene was digested with EcoRI and serves as a negative control.
Preparation of Microsomes. Tissues were homogenized in ice-cold buffer [50 mM Tris-HCl, 150 mM KCl, 20% (v/v) glycerol, 1 mM disodium ethylenediaminetetraacetate, and 0.5 mM 4-amidinophenylmethanesulfonyl fluoride, pH 7.4], and microsomes were isolated by differential centrifugation as described previously (Granvil et al., 2003; Cheung et al., 2005). Protein concentrations were determined using a BCA Protein Assay Kit (Pierce Chemical Co., Rockford, IL), following the manufacturer's instructions.
Western Blot Analysis. Microsomal proteins (5–30 μg) from each sample were separated by SDS-PAGE through 10% polyacrylamide gels and transferred onto nitrocellulose membranes (Schleicher and Schuell, Keene, NH). After probing with antibodies against human CYP3A4 and CYP3A7 and rat CYP3A1, CYP1A2, CYP2A3, CYP2B1, CYP2E1, or mouse GAPDH, the membranes were incubated with the appropriate secondary antibodies (anti-rabbit or anti-mouse IgG horseradish peroxidase purchased from Sigma) followed by ECL detection (Pierce Chemical Co.) as described previously (Granvil et al., 2003; Cheung et al., 2005; Yu et al., 2005). Blots were scanned and the relative intensity of each band was analyzed using Kodak 1D (version 3.6.3) Scientific Imaging Systems software (New Haven, CT).
RNA Analysis. Total RNA was isolated from liver tissues with TRIzol reagent (Invitrogen, Carlsbad, CA) by following the manufacturer's instructions, and the concentrations were determined by optical densitometry at 260 nm. SYBR green-based quantitative real-time polymerase chain reaction (QPCR) analysis of the specific P450 and 18S mRNAs in liver samples was carried out using an Applied Biosystems 7900HT instrument (Applied Biosystems, Foster City, CA) and gene-specific primers under conditions reported previously (Wiwi et al., 2004). Primers used for analysis of CYP3A4 were 5′-ATG AAA GAA AGT CGC CTC GAA-3′ and 5′-AAG GAA ATC CAC TCG GTG CTT-3′.

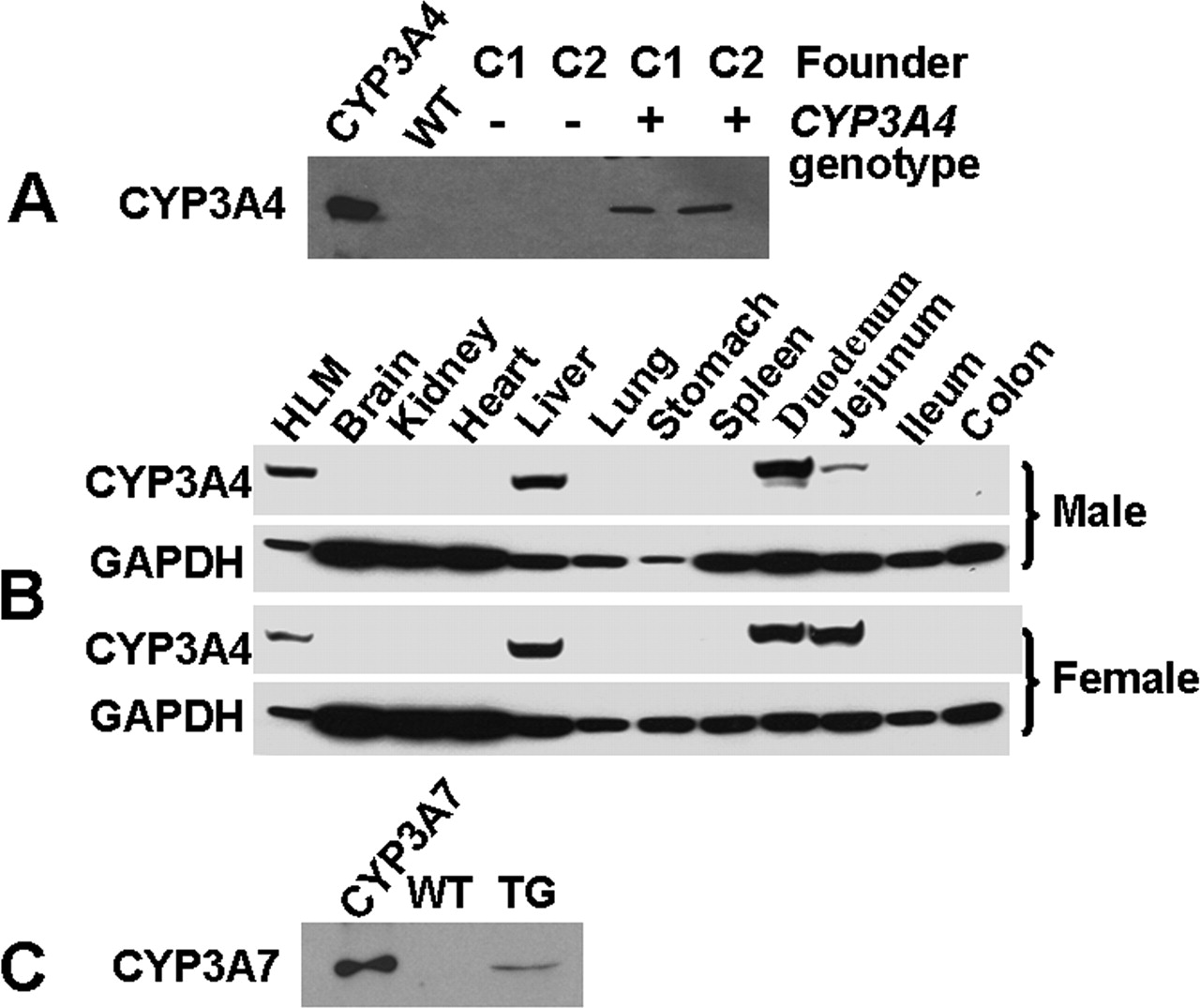
Expression of CYP3A4 and CYP3A7 in transgenic mice. Western blot analyses show that: A, CYP3A4 is expressed in the liver of CYP3A4 genotype-positive 3-week-old mice from founder lines C1 (male liver sample) and C2 (female liver sample); B, expression of CYP3A4 is detected in the liver, duodenum, and jejunum of 3-week-old male and female mice (three in each group, pooled samples were used in the study) of the C2 founder line; and C, the human fetal form CYP3A7 is expressed in the transgenic fetal liver between embryonic days 16 and 17 (pooled sample from >10 individuals). Microsomal proteins (20 μg of each well) were separated by SDS-PAGE, transferred onto a membrane, and CYP3A4, GAPDH, and CYP3A7 were detected using the Enhanced Chemiluminescence Kit (Pierce Chemical Co.). The monoclonal antibody against CYP3A4 (mAb 275-1-2) (Gelboin et al., 1995) used in these experiments reacts with CYP3A4 but does not recognize murine Cyp3a or other liver proteins. This antibody was used to screen positive founder lines and to assay the tissue distribution of CYP3A4. A specific polyclonal antibody that only recognizes CYP3A7 (Sim et al., 2005) was used for the evaluation of CYP3A7 expression in fetal livers. Recombinant CYP3A4 and CYP3A7 protein and pooled human liver microsomes (HLMs) were used as positive controls.
Statistical Analysis. All values are expressed as the means ± S.D. unless indicated otherwise. All data were analyzed by paired or unpaired Student's t test (Prism, version 3.02; GraphPad Software Inc., San Diego, CA), and the difference was considered significant if p was <5%.
Results
Generation of CYP3A4-CYP3A7 Transgenic Mice. A transgenic mouse line was created using the BAC clone RP11-757A13, containing the complete CYP3A4 and CYP3A7 genes together with over 35 kb of 5′-flanking DNA and 9 kb of 3′-flanking DNA (Fig. 1A). Hybridization analyses were performed with 32P-labeled probes including CYP3A4 cDNA and oligonucleotides recognizing specific regions of exons 1 and 13 and sequences –10 kb upstream of the CYP3A4 gene were used to verify the BAC clone. The size of each hybridized band corresponded well with the size predicted from the map of the CYP3A4 gene sequence (Fig. 1B). Transgenic mice containing the human CYP3A4 and CYP3A7 genes were generated (see Materials and Methods), and the presence of the CYP3A4 transgene was confirmed by Southern blotting at each breeding step. Two positive founders, designated C1 and C2, were shown to carry the CYP3A4 transgene. Western blot analysis with a monoclonal antibody that recognizes CYP3A4 but not any endogenous murine Cyp3a proteins revealed that CYP3A4 was expressed in the livers of 3-week-old CYP3A4-positive mice from both founders (Fig. 2A). The C2 founder was selected without any bias to generate a homozygous transgenic mouse line for the following studies.
Tissue Distribution of CYP3A4 and CYP3A7 Proteins in Transgenic Mice. Western blots were performed to assess the expression of CYP3A4 and CYP3A7 proteins in transgenic mouse tissues. CYP3A4 protein was detected in the liver but was not detectable in brain, kidney, heart, lung, stomach, spleen, or colon of 3-week-old mice (Fig. 2B). To determine the expression of CYP3A4 in the gut, small intestine was segmented into duodenum, jejunum, and ileum. Western blots of microsomal proteins indicated that CYP3A4 was expressed in the duodenum and jejunum but not in the ileum. Interestingly, CYP3A4 levels are comparable between duodenum and jejunum for females, whereas it is lower in jejunum for males (Fig. 2B). Analysis of GAPDH protein as a loading control verifies that the intestinal protein was of similar quality and quantity between male and female jejunum samples, thus indicating that low expression in males was not caused by degradation. Furthermore, CYP3A7 protein was expressed in transgenic fetal liver (Fig. 2C) as shown using a specific antipeptide antibody. Although the CYP3A4 was not detected in kidney from 3-week-old transgenic mice by Western blotting, we cannot conclude that the protein is absent since this tissue, unlike liver and gut, is complex with many different cell types. Further studies using immunohistochemistry and tissues from mice at different age will be required to address whether CYP3A4 is expressed in kidney and other complex tissues like the brain.
Sexual Dimorphism of CYP3A4 Expression in Transgenic Mice. The impact of sex and age on the expression of CYP3A4 in the transgenic mouse livers was investigated since both factors can affect hepatic CYP3A4 expression (Yu et al., 2005). CYP3A4 protein was expressed in the livers of male mice from 1 to 4 weeks of age, decreased in 5-week-old mice, and was undetectable from age 6 to 24 weeks of age. In contrast, hepatic CYP3A4 expression remained at similar levels in the 5- to 24-week-old female mice of this transgenic line (Fig. 3). In contrast to the differential expression of CYP3A4 between adult males and females, the developmental expression patterns for murine Cyp1a2 and GAPDH protein showed no difference between the sexes from ages 3 to 24 weeks (Fig. 3). Further studies revealed similar age- and sex-dependent profiles for hepatic CYP3A4 mRNA, namely, suppression in males but not females after the age of 5 weeks. Moreover, similar profiles were apparent for the endogenous mouse liver gene Cyp3a44, whose expression was also female-specific in adults (Table 1). In contrast, expression of a related mouse gene, Cyp3a25, was not age- or sex-dependent.
Sex-dependent developmental regulation of human CYP3A4 and mouse Cyp3a genes in transgenic mice
Hepatic RNA levels were determined for each CYP3A gene by QPCR using transgenic liver tissue isolated from mice at each age, as indicated. Data shown are mean ± S.E. values for n = 4 to 5 livers/group, normalized to 18S RNA levels, with the 8-week male liver level of each RNA set to 1.

CYP3A4 developmental regulation in transgenic male and female mouse livers. Liver tissues were collected from transgenic male and female mice of different ages, and microsomes were prepared by differential centrifugation. Pooled (four to five livers in each group) samples were used for Western blot analysis of CYP3A4, murine Cyp1a2, and GAPDH protein levels. CYP3A4 protein is essentially absent in the male livers beginning at week 6, whereas it is consistently expressed in female liver at a level that is similar at all ages after 2 weeks.
Induction of CYP3A4 and Murine Cyp3a by Phenobarbital. Treatment of the transgenic mice with phenobarbital induced the CYP3A4 transgene (Fig. 4). CYP3A4 was not detectable in the livers of 6- to 7-week-old transgenic male mice, in agreement with the results shown in Fig. 3, but was detected in females treated with saline. Phenobarbital, a known activator of the nuclear receptors PXR and CAR, induced CYP3A4 expression ∼11-fold in the transgenic female livers. Moreover, phenobarbital induced CYP3A4 in the male transgenic livers to a level comparable with that induced in the females. Murine Cyp3a protein levels were also elevated, approximately 3- to 4-fold, compared with saline-treated controls (Fig. 4). Induction of CYP3A4 and murine Cyp3a was associated with significantly elevated mRNA levels and resulted in sharply increased Cyp3a enzymatic activity (data not shown), indicating the potential to alter the metabolism and pharmacokinetics of Cyp3a drugs.
Growth Hormone Stimulates Expression of Human CYP3A4 and Murine Cyp3a44 in Transgenic Male Mice. Because plasma GH profiles are an important physiological regulator of sex-dependent CYP3A gene expression, we evaluated the effect of GH treatment on CYP3A4 expression in transgenic mouse livers. Recombinant mouse GH was administered as a continuous infusion over 7 days, which overrides the male, pulsatile plasma GH pattern, and in the case of the rat feminizes liver gene expression (Mode et al., 1981; Ahluwalia et al., 2004; Holloway et al., 2006). Western blot analysis revealed that liver CYP3A4 expression was strongly induced in the transgenic males treated with continuous GH. In contrast, GH treatment did not alter hepatic CYP3A4 expression in the females (Fig. 5). Continuous GH infusion also stimulated the expression of a female-specific murine Cyp2b form in the male mice. Murine Cyp1a2, Cyp2a, and Cyp2e1 levels were lower in both male and female mice treated with GH, compared with sham-operated controls (Fig. 5). As a control, the content of GAPDH protein was shown to be unchanged in both males and females after GH treatment. In addition, CYP3A7 protein was undetectable in both the control and GH-treated male and female mouse livers (data not shown).
To further investigate whether the effects of GH are due to transcriptional regulation and to provide more quantitative analysis, mRNA levels were determined by QPCR analysis. CYP3A4 mRNA was sharply increased in the GH-treated male mice (approximately 30-fold higher than in sham-treated control males), whereas there was no change in the GH-treated females (Fig. 6A), a result that is consistent with Western blot analysis (Fig. 5). Continuous GH treatment also induced the endogenous murine Cyp3a44 gene in the transgenic male livers, to a level ∼130-fold higher than sham control males and comparable with that of correspondingly treated females (Fig. 6B). In contrast, GH had no significant effect on Cyp3a11 and Cyp3a25 mRNA levels in males or females (Fig. 6, C and D), which serve as examples of sex-independent and GH-unresponsive hepatic Cyp3a genes. Together, these findings demonstrate that the plasma GH profile is an important factor in modulating sex-dependent expression of the CYP3A4 transgene at the transcriptional level, as it is in modulating the sex-dependent expression of the endogenous mouse Cyp3a44 gene.

Induction of hepatic CYP3A4 and murine Cyp3a proteins by phenobarbital (PB). Transgenic mice (6–7 weeks old) were treated daily for 4 days with saline or phenobarbital as detailed under Materials and Methods. Liver microsomes from individual mice were prepared and analyzed by SDS-PAGE. CYP3A4 and murine Cyp3a proteins were detected with mAb 275-1-2 and mAb 2-13-1 (Gelboin et al., 1995), respectively, by using the Enhanced Chemiluminescence Kit. mAb 2-13-1 reacts strongly with murine Cyp3a and very weakly with human CYP3A4 (Yu et al., 2005). Although CYP3A4 is not detectable in control male livers, phenobarbital induces CYP3A4 expression in the males, resulting in a level comparable with that of phenobarbital-treated females.

Stimulation of CYP3A4 protein expression in transgenic male mouse livers by continuous GH treatment. Mice (7 weeks old, four to five in each group) were treated with recombinant mouse GH by continuous infusion for 7 days using Alzet osmotic minipumps. Sham-treated mice were used as controls. Tissue microsomes, prepared from individual mouse liver by differential centrifugation, were separated by SDS-PAGE and subjected to Western blot analysis with enhanced chemiluminescence detection of CYP3A4 and murine Cyp2b, Cyp1a2, Cyp2a, Cyp2e1, and GAPDH. GH treatment induced CYP3A4 protein expression in transgenic male mice. Induction of a female-specific Cyp2b protein band was also seen in the GH-treated males. Partial decreases in several of the other P450 proteins, most notably Cyp1a2, were seen in the GH-treated males and females.

QPCR analysis of hepatic CYP3A4 and murine Cyp3a mRNA levels in mice treated with GH. Total mRNAs were prepared from mouse livers by using TRIzol reagent, and cDNA was synthesized by reverse transcription. QPCR analysis was carried out with SYBR Green I polymerase chain reaction master mix (Applied Biosystems). Data shown are relative expression levels after normalized to the level of 18S RNA and represent mean ± S.E. values for four to five individual livers per group, with the sham-treated male RNA level set to 1. Hepatic CYP3A4 and Cyp3a41 mRNA levels were significantly higher (p < 0.05) in the GH-treated males (M) than in sham-treated males. GH treatment of female (F) mice had no significant impact on P450 mRNA levels.
Discussion
This study characterizes a new transgenic mouse line carrying human CYP3A4 and CYP3A7 transgenes and its use for investigation of CYP3A gene regulation. The results reveal that hepatic CYP3A4 is subject to developmental expression that is sexually dimorphic, as manifest by the low to undetectable level of CYP3A4 protein and mRNA in the postpubertal male transgenic mice. Moreover, CYP3A4 expression in adult male livers is shown to be induced by GH administered as a 7-day continuous infusion.
Sexual dimorphism in the expression of several hepatic P450 drug-metabolizing enzymes has been demonstrated and studied extensively in rats (Waxman et al., 1991; Shimada et al., 1995; Robertson et al., 1998; Kawai et al., 2000). In particular, rat CYP3A2 and CYP3A18 genes are expressed in a male-specific manner, whereas CYP3A9 is female-specific. Murine Cyp3a41 and Cyp3a44 correspond to female-specific forms (Sakuma et al., 2002). Human CYP3A4 seems to be expressed at higher levels in women than men (Greenblatt et al., 1980; Watkins et al., 1989; Gorski et al., 1998; Wolbold et al., 2003). However, other studies show no sex dependence for CYP3A4 (Schmucker et al., 1990; George et al., 1995). A recent study using a large collection (n = 94) of well characterized surgical liver samples revealed, however, a 2-fold higher level of CYP3A4 protein and RNA in women compared with men (Wolbold et al., 2003). Previous investigations with a CYP3A4 transgenic mouse line (Yu et al., 2005) and in the present study using a mouse line made with a larger transgene containing both the CYP3A4 and CYP3A7 genes and their 5′- and 3′-flanking DNA revealed that hepatic CYP3A4 expression is sexually dimorphic in transgenic adult mice but not in prepubertal mice. Given the dominant role of CYP3A4 in human hepatic drug metabolism and given the finding that the drug efflux pump P-glycoprotein does not show significant sexual dimorphism in human liver (Wolbold et al., 2003), CYP3A4 and its sexually dimorphic expression could be a major factor contributing to sex-dependent drug clearance in humans. Nevertheless, the question is still open whether CYP3A5 can compensate for lower CYP3A4 levels since CYP3A4 expression is known to be highly variable in humans (Stevens et al., 2003).
The polypeptide hormone GH, which is secreted by the pituitary gland in a sexually dimorphic manner, regulates expression of sex-specific or -dominant P450 genes. As an essential hormone for normal linear growth, GH is secreted in pulses, but with differentiated levels and frequencies in men and women (van den Berg et al., 1996; Jaffe et al., 1998). Treatment with exogenous GH has been shown to alter CYP3A expression in rats (Robertson et al., 1998; Kawai et al., 2000), elevate Cyp3a41 and Cyp3a44 expression in mice (Sakuma et al., 2002), and increase CYP3A4 activity in humans (Jaffe et al., 2002). In agreement with these findings, the present study demonstrates that GH plays a key role in the sexually dimorphic expression of the CYP3A4 transgene in mice through a pretranslational mechanism. Signal transducer and activator of transcription (STAT) 5b has been identified as an essential determinant of GH-mediated sexual expression of P450 enzymes (Park et al., 1999; Holloway et al., 2006); however, the precise intracellular regulatory cascade that STAT5b activates leading to the regulation of hepatic P450 expression is only partially understood and awaits more extensive investigation. Furthermore, the absence of CYP3A4 protein in transgenic adult male mice is in stark contrast to the stable, albeit variable expression of CYP3A4 in humans, suggesting potential influences of the hormonal environment, inducers, or other regulatory mechanisms in human subjects that are absent in the mouse model. Rather, the present finding establishes that the human CYP3A4 gene contains all of the DNA sequence elements required to respond to the endogenous mouse hormonal environment, leading to a pattern of postnatal developmental regulation, adult sexual dimorphism, and plasma GH responsiveness that is very similar to that of the endogenous mouse Cyp3a44 gene. This is an important observation that may facilitate further studies on these regulatory events and the extent to which they are conserved between mice and humans.
In clinic pharmacotherapy, GH has been used to treat GH-deficient growth-retarded children. GH treatment stimulates normal linear growth and can be used to improve bone density in prepubertal patients (Lanes et al., 1996; Powell et al., 1997). The elderly and GH-deficient adults can also benefit from GH replacement therapy, as indicated by increased lean mass, declined adiposity, and obviously restored skin thickness (Rudman et al., 1990; Hana et al., 2004), although the balance of benefits and risks and GH dosage have not been defined (Marcus and Hoffman, 1998; Cummings and Merriam, 2003). In one placebo-controlled clinical study, where the effects of GH treatment on P450 activity were investigated in elderly men, a reduction in CYP1A2 activity was observed with no significant change in CYP3A4 activity (Jurgens et al., 2002). The lack of effect on CYP3A4 activity could be due to the utilization of endogenous cortisol metabolism as the marker of CYP3A4 activity, which may not be a reliable indicator of CYP3A4-dependent metabolism in vivo (Galteau and Shamsa, 2003). The present CYP3A4 transgenic model, in agreement with earlier studies using human hepatocyte cultures (Liddle et al., 1998) and human subjects (Jaffe et al., 2002), suggests that continuous GH treatment not only suppresses CYP1A2 expression but also increases CYP3A4 expression and activity. These GH-dependent changes in hepatic P450 expression may significantly alter the pharmacokinetics and pharmacodynamics of drugs metabolized by CYP1A2 or CYP3A4. Changes in drug efficacy may result, perhaps leading to adverse effects as a consequence of the interactions between GH and P450-metabolized drugs.
The development of P450-humanized mouse models (Gonzalez, 2003) provides a means to overcome species differences in drug metabolism, which are, in large part, associated with differences in the genes encoding P450 drug-metabolizing enzymes. These humanized mouse models have provided valuable tools for studying the function and regulation of cytochromes P450 in a whole animal system, where the functional significance of human P450 expression can be evaluated under controlled conditions, and the regulatory networks and mechanistic basis for responses to xenobiotics can be delineated in preclinical studies. These models could also have implications in the understanding of hormone homeostasis and human disease. However, although the physiological environments are obviously different between mice and humans, particularly in body weight, blood flow rate, and hormone profiles, this gap may be filled by using an allometric scaling approach to provide quantitative assessments.
In conclusion, the human CYP3A transgenic mouse line described here provides a means for studying the regulation and function of human CYP3A4 and CYP3A7 genes in a whole animal model. Sexually dimorphic expression of CYP3A4 was observed and could lead to significant differences in drug metabolism and disposition, pharmacokinetics, and pharmacodynamics between men and women. These findings, taken in the context of earlier reports, suggest that caution should be advised regarding the potential for altered drug metabolism when prescribing CYP3A4 drugs to patients receiving GH replacement therapy.
Acknowledgments
We thank John R. Buckley for technical assistance.
Footnotes
- Received August 17, 2005.
- Accepted November 14, 2005.
-
This work was supported by the Intramural Research Program of the National Cancer Institute (to F.J.G.), by National Institutes of Health Grant DK33765 (to D.J.W.), and by Wellcome Trust Postdoctoral Research Fellowship 064866 (to C.C.).
-
C.C. and A.-M.Y. contributed equally to this work.
-
doi:10.1124/jpet.105.094367.
-
ABBREVIATIONS: P450, cytochrome P450; PXR, pregnane X receptor; CAR, constitutive androstane receptor; BAC, bacterial artificial chromosome; mAb, monoclonal antibody; GH, growth hormone; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PAGE, polyacrylamide gel electrophoresis; QPCR, quantitative real-time polymerase chain reaction.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}