


Abstract
Human immunodeficiency virus-infected patients on antiretroviral drug therapy frequently experience hepatotoxicity, the underlying mechanism of which is poorly understood. Hepatotoxicity from other compounds such as bosentan and troglitazone has been attributed, in part, to inhibition of hepatocyte bile acid excretion. This work tested the hypothesis that antiretroviral drugs modulate hepatic bile acid transport. Ritonavir (28 μM), saquinavir (15 μM), and efavirenz (32 μM) inhibited [3H]taurocholate transport in bile salt export pump expressing Sf9-derived membrane vesicles by 90, 71, and 33%, respectively. In sandwich-cultured human hepatocytes, the biliary excretion index (BEI) of [3H]taurocholate was maximally decreased 59% by ritonavir, 39% by saquinavir, and 20% by efavirenz. Likewise, in sandwich-cultured rat hepatocytes, the BEI of [3H]taurocholate was decreased 100% by ritonavir and 94% by saquinavir. Sodium-dependent and -independent initial uptake rates of [3H]taurocholate in suspended rat hepatocytes were significantly decreased by ritonavir, saquinavir, and efavirenz. [3H]Taurocholate transport by recombinant NTCP and Ntcp was inhibited by ritonavir (IC50 = 2.1 and 6.4 μM in human and rat, respectively), saquinavir (IC50 = 6.7 and 20 μM, respectively), and efavirenz (IC50 = 43 and 97 μM, respectively). Nevirapine (75 μM) had no effect on bile acid transport in any model system. In conclusion, ritonavir, saquinavir, and efavirenz, but not nevirapine, inhibited both the hepatic uptake and biliary excretion of taurocholate.
Hepatotoxicity is a significant adverse event in human immunodeficiency virus-infected individuals treated with antiretroviral therapy. Incidence rates of 13 to 30% have been reported for severe antiretroviral-associated hepatotoxicity, defined as aspartate aminotransferase and alanine aminotransferase > 5× the upper limit of normal (Rodriguez-Rosado et al., 1998; den Brinker et al., 2000; Sulkowski et al., 2000; Martinez et al., 2001). Most commonly, these liver enzyme elevations force the withdrawal of an otherwise effective therapy in human immunodeficiency virus-infected patients. However, fatalities due to antiretroviral-induced hepatotoxicity have also been reported (Centers for Disease Control and Prevention, 2001).
Retrospective investigations examining the predisposing factors of antiretroviral-associated hepatotoxicity have implicated several drugs. However, there is no consensus on which antiretroviral agents are consistent offenders. One investigation reported that ritonavir-containing regimens increased the risk of hepatotoxicity by 8.6-fold (Sulkowski et al., 2000). Other studies have suggested that nevirapine contributes to hepatotoxicity (Clarke et al., 2000; Piroth et al., 2000; Martinez et al., 2001). One study, however, did not find that any particular antiretroviral agent increased the risk of hepatotoxicity (den Brinker et al., 2000). Additional risk factors have been identified and include length of therapy, coinfection with hepatitis C virus, and higher baseline alanine aminotransferase values. The mechanisms underlying antiretroviral-induced hepatotoxicity are unclear. Current hypotheses focus on the effects of either improved immune function (French et al., 2004) or impaired drug metabolizing enzyme activity (Kaplowitz, 2004). However, these hypotheses are controversial (Reijers et al., 2000; de Maat et al., 2002; Martin-Carbonero et al., 2002; Dailly et al., 2004).
Hepatotoxicity associated with troglitazone, bosentan, and cyclosporin A has been attributed, in part, to alterations in hepatic bile acid transport. In rats, single i.v. doses of troglitazone and bosentan resulted in increased plasma bile acid concentrations (Fattinger et al., 2001; Funk et al., 2001). In one clinical investigation, patients taking bosentan who developed increased aminotransferase enzymes also exhibited concomitant increases in plasma bile acid concentrations (Fattinger et al., 2001). Single i.v. doses of cyclosporin A can also cause dose-dependent elevations in serum bile acid concentrations in rats (Roman et al., 1990).
The bile salt export pump (BSEP; ABCB11, sister of P-glycoprotein) is the hepatic transport protein predominantly responsible for biliary secretion of conjugated and unconjugated bile acids. Since bosentan, cyclosporin A, and troglitazone all inhibit BSEP function, this inhibition has been proposed as a mechanism of drug-induced hepatotoxicity (Stieger et al., 2000; Fattinger et al., 2001; Byrne et al., 2002).
Currently, there are no data on the effects of antiretroviral agents on BSEP function. Therefore, the goal of this work was to examine the effects of selected protease inhibitors (ritonavir and saquinavir) and nonnucleoside reverse transcriptase inhibitors (efavirenz and nevirapine) on hepatic bile acid transport. The effects of these antiretroviral drugs on the transport of taurocholate (a model bile acid) was studied using sandwich-cultured primary human and rat hepatocytes, isolated suspended rat hepatocytes, and transporter expressing cell-based systems.
Materials and Methods
Chemicals. Ritonavir, nevirapine, saquinavir, and efavirenz were obtained through the National Institutes of Health AIDS Research and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases, National Institutes of Health. Collagenase (Type I) was obtained from Worthington Biochemicals (Freehold, NJ). Dulbecco's modified Eagle's medium (DMEM), amphotericin B, and insulin were purchased from Invitrogen (Carlsbad, CA). Insulin/transferrin/selenium culture supplement, Matrigel, and Biocoat plates were purchased from BD Biosciences Discovery Labware (Bedford, MA). Penicillin-streptomycin solution, William's E Medium, fetal bovine serum (FBS), taurocholic acid, dexamethasone, Triton X-100, and soybean trypsin inhibitor were purchased from Sigma-Aldrich (St. Louis, MO). Anti-rat Bsep antibody (K12) cross-reacts with human BSEP and was kindly provided by Dr. Peter Meier (Department of Medicine, University Hospital, Zurich, Switzerland). All other chemicals and reagents were of analytical grade and readily available from commercial sources.
Animals. Male Wistar rats (270–235 g) were obtained from Charles River Laboratories, Inc. (Raleigh, NC) and used as liver donors. Animals had free access to water and food before surgery. All animal procedures complied with the guidelines of the Institutional Animal Care and Use Committee (University of North Carolina, Chapel Hill, NC).
Wild-Type BSEP Plasmid Construction. The full open reading frame of human BSEP cDNA was obtained by PCR using Expand Long Template PCR System (Roche Applied Science, Indianapolis, IN) from a cDNA library synthesized from human liver mRNA using oligonucleotide primers 5′-TGTGGGTTGCAATTACCATGTCTGACTCAG-3′ as the forward and 5′-GCATTGGGTCAACTGATGGGGGATCCAGTG-3′ as the reverse (melting temperature, 94°C × 30 s; annealing temperature, 57°C × 45 s; extension temperature, 68°C × 6 min for 30 cycles). A single PCR product (∼4.1 kb) was visualized on an ethidium bromide-stained 0.8% agarose gel. An aliquot of the PCR product was ligated into the pEF6/V5-His-TOPO plasmid expression vector (Invitrogen). After transformation and growth in Escherichia coli, individual colonies containing the pEF6/V5-His-TOPO/BSEP construct in the sense orientation were identified with an appropriate restriction endonuclease digest. The only full-length clone tolerant of propagation in E. coli was found to have seven nonsynonymous mutations when fully sequenced with an ABI 3700 DNA analyzer (Applied Biosystems Inc., Foster City, CA). It was subsequently found that BSEP-transformed bacteria showed no or minimal growth due to the presence of a cryptic bacterial promoter between positions 59 and 81 downstream of the start codon of BSEP cDNA (Noe et al., 2002). The cryptic promoter was inactivated by introducing synonymous base pair changes at positions 76 to 81 (TATAAT→TACAAC), and each of the seven nonsynonymous mutations was reverted to the wild-type reference allele using QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA) and confirmed by direct DNA sequencing, resulting in a fully functional wild-type BSEP clone with 100% amino acid identity to the BSEP reference sequence (GenBank accession number NM_003742).
[3H]Taurocholate Transport Studies in BSEP-Expressing Sf9 Cell-Derived Vesicles. Recombinant pFASTBAC1 plasmids containing either human BSEP or green fluorescence protein (GFP)-coding sequence were constructed. Recombinant baculovirus was generated with BAC-TO-BAC baculovirus expression system (Invitrogen). In brief, pFASTBAC1 recombinant plasmids were transformed into competent DH10BAC1 cells containing helper and shuttle plasmids. E. coli colonies with recombinant bacmid were selected on Luria Agar plates containing 50 μg/ml kanamycin, 7 μg/ml gentamicin, and 10 μg/ml tetracycline. Purified recombinant bacmid DNA was used to transfect Sf9 insect cells in the presence of Cell-fectin reagent (Invitrogen). Recombinant baculovirus was harvested in the supernatant of cell culture medium after 72 h of culture at 27°C. Recombinant baculovirus stocks were stored at 4°C. Sf9 insect cells were cultured in plastic Erlenmeyer flasks (Nalge Nunc International, Rochester, NY) in a shaking 27°C incubator in Grace's Insect Cell Medium supplemented with 10% FBS and 100 units/ml penicillin G, 100 μg/ml streptomycin sulfate, and 0.25 μg/ml amphotericin B. Log phase cells (1–1.3 × 106 cells/ml) were infected with recombinant baculovirus. Cells were harvested between 70 and 75 h after infection. Plasma membrane vesicles were prepared as described, with modifications (Loe et al., 1996). In brief, cells were homogenized in buffer containing 250 mM sucrose, 50 mM Tris, pH 7.5, 0.24 mM CaCl2, and protease inhibitor mixture tablets (Roche Diagnostics, Indianapolis, IN). Cells were disrupted by N2 cavitation (a 5-min equilibration at 200 p.s.i.) and then released to atmospheric pressure, and EDTA was added to a final concentration of 1 mM. The suspension was centrifuged at 800g at 4°C for 10 min, and the supernatant was layered onto 10 ml of a 35% (w/w) sucrose and 50 mM Tris, pH 7.4 cushion. After centrifugation at 100,00g at 4°C for 1 h, the interface was removed and placed in a 25 mM sucrose and 50 mM Tris, pH 7.4 solution and centrifuged at 100,000g at 4°C for 30 min. The membranes were washed with Tris-sucrose buffer (250 mM sucrose, 50 mM Tris, pH 7.4) and then resuspended by passing repeatedly through a 27-gauge needle.
[3H]Taurocholate (1 μM) uptake into Sf9-derived vesicles was measured by a quick filtration method (Loe et al., 1996). The transport assay buffer used was HEPES (50 mM, pH 7.5), KCl (100 mM), and contained ATP or AMP (4 mM), MgCl2 (10 mM), creatine phosphate (10 mM), creatine kinase (100 μg/ml), glutathione reductase (5 μg/ml), and NADPH (0.25 mM). Preliminary experiments determined uptake was linear through 5 min. Frozen membrane vesicles were thawed quickly in a 37°C water bath. Aliquots of membrane suspensions (15 μg) were preincubated for 5 min at 37°C, and uptake was initiated by addition of 40 μl of prewarmed incubation medium to the membrane suspensions. Vesicle uptake was terminated at 3 min by the addition of 800 μl of ice-cold transport buffer. Vesicle-associated uptake was separated from free [3H]taurocholate by rapid filtration through a 0.45-μm HAWP25 filter (Millipore Corporation, Bedford, MA). Filters were rinsed twice with 3.5 ml of ice-cold membrane suspension buffer. Filters were immersed in 5 ml of liquid scintillation cocktail and assayed for radioactivity. Nonspecific binding of [3H]taurocholate to the filter was determined in the absence of membrane vesicles. Net ATP-dependent [3H]taurocholate uptake was calculated as the difference between uptake in the presence of AMP and ATP.
Western Blot. Sandwich-cultured hepatocytes were lysed with 1% SDS/1 mM EDTA supplemented with Complete Protease Inhibitor (Roche Diagnostics) and stored at –80°C until analysis. Sf9 cell-derived vesicles were suspended in 1% SDS/1 mM EDTA and stored in the same manner. Protein (approximately 30 μg/lane) was resolved with electrophoresis on a NuPage 4 to 12% Bis-Tris gel (Invitrogen) and subsequently transferred onto polyvinylidene difluoride membranes. Membranes were blocked overnight at 4°C with 5% nonfat milk. Thereafter, membranes were incubated with the primary anti-Bsep antibody (1:2000) for 2 h followed by incubation with the secondary anti-Rabbit IgG (1:4000) for 1 h. Membranes were developed with SuperSignal West Dura (Pierce Biotechnology, Rockford, IL). The chemiluminescence signal intensity was quantified with a Versa-Doc 1000 molecular imager (Bio-Rad, Hercules, CA).
Human Hepatocytes. Human hepatocytes were purchased from various commercial sources (Zen-Bio, Research Triangle Park, NC; CellzDirect, Tucson, AZ). Hepatocytes were isolated from two female and four male donors with ages ranging from 11 to 88 years old. Five donors were Caucasian, and one was African American. None of the liver donors had any known hepatic impairment, and none had been taking known hepatotoxic agents previously. Livers were not cirrhotic at the time of harvest.
Hepatocyte Isolation and Culture. Rat hepatocytes were isolated from male Wistar rats using a collagenase perfusion as described previously (LeCluyse et al., 1996). Hepatocyte viability was >85% as determined by trypan blue exclusion. Hepatocyte suspensions were plated in six-well Biocoat dishes at a density of approximately 1.5 million cells/well in supplemented DMEM containing 5% FBS and 0.1 μM dexamethasone. One to 3 h after plating, hepatocyte cultures were lightly agitated, and nonadherent cells were aspirated. After 24 h of incubation at 37°C, cells were overlaid with Matrigel at a final protein concentration of 0.25 mg/ml, and cultures were maintained in serum-free DMEM containing insulin/transferrin/selenium with 0.1 μM dexamethasone for 4 days. Culture medium was replaced every 24 h.
Human hepatocytes were treated in a similar fashion to rat hepatocytes but were kept first in supplemented William's E Medium containing 5% fetal bovine serum and 1.0 μM dexamethasone. Four to 6 h after plating, cultures were changed to serum-free William's E Medium and maintained for up to 6 days.
Bile Acid ([3H]Taurocholate) Transport Studies in Sandwich-Cultured Hepatocytes. Cells were rinsed twice with 2 ml of warm standard HBSS, Hanks' balanced salt solution (HBSS) or Ca2+-free HBSS and preincubated for 10 min at 37°C with 1.5 ml of standard or Ca2+-free HBSS. Ca2+-free HBSS was used to disrupt canalicular networks; accumulation in Ca2+-free HBSS represents accumulation in cells only. Canalicular networks remain intact in standard HBSS and substrate accumulates in both cells and bile canaliculi. To measure [3H]taurocholate accumulation, hepatocytes were incubated with 1 μM[3H]taurocholate in standard HBSS for 10 min. Both preincubation and incubation solutions contained one of four antiretroviral agents in concentrations inclusive of those used clinically: 0 to 28 μM ritonavir (0–20 μg/ml), 0 to 75 μM nevirapine (0–20 μg/ml), 0 to 15 μM saquinavir (0–10 μg/ml), or 0 to 32 μM efavirenz (0–10 μg/ml). Cells were rinsed vigorously three times with ice-cold standard HBSS, and lysed with 1 ml of 0.5% Triton X-100. A blank dish in each group was used to control for any nonspecific binding effects. Cell lysates were analyzed by liquid scintillation spectroscopy (Packard Tricarb; Packard Corp., Meriden, CT). Total protein in each dish was determined by the bicinchoninic acid method (Smith et al., 1985), and total accumulation was normalized to protein content for each sample. The biliary excretion index (BEI; B-CLEAR; Qualyst, Inc., Research Triangle Park, NC) was calculated as the difference in [3H]taurocholate accumulation in the presence and absence of intact canalicular networks (cells + bile canaliculi versus cells) normalized for [3H]taurocholate accumulation in the presence of intact canalicular networks (cells + bile canaliculi). The BEI represents the fraction of accumulated substrate that resides in the bile canalicular compartment.
Suspended Rat Hepatocytes. [3H]Taurocholate uptake was determined in isolated suspended rat hepatocytes as described previously (Vore et al., 1996). In brief, suspended hepatocytes (4 ml; 0.8–1.2 × 106 cells/ml) were allowed to preincubate in a 37°C orbital shaking water bath for 5 min. Uptake was initiated by the addition of 1 μM [3H]taurocholate in the presence or absence of various concentrations of inhibitor (ritonavir, nevirapine, saquinavir, or efavirenz) to the hepatocytes. Hepatocytes were sampled at 15, 30, and 45 s, and uptake was terminated by immediate centrifugation of hepatocytes through a silicone/mineral oil layer into 3 M KOH. Cell pellets were analyzed by liquid scintillation spectroscopy. Adherent fluid volume also was determined by incubating hepatocytes with [14C]inulin. An aliquot of suspended cells was assayed for protein concentration by the bicinchoninic acid method. To determine the relative contributions of antiretroviral effects on sodium-dependent (Ntcp) and sodium-independent (e.g., Oatps) uptake processes in suspended rat hepatocytes, separate experiments were conducted in the presence and absence of sodium. In the presence of sodium, all transport processes, including Ntcp and Oatps, mediate accumulation of [3H]taurocholate. Accumulation of [3H]taurocholate in the absence of sodium is mediated by all non-Ntcp-dependent processes. Choline was used as the replacement for sodium in the sodium-free buffer.
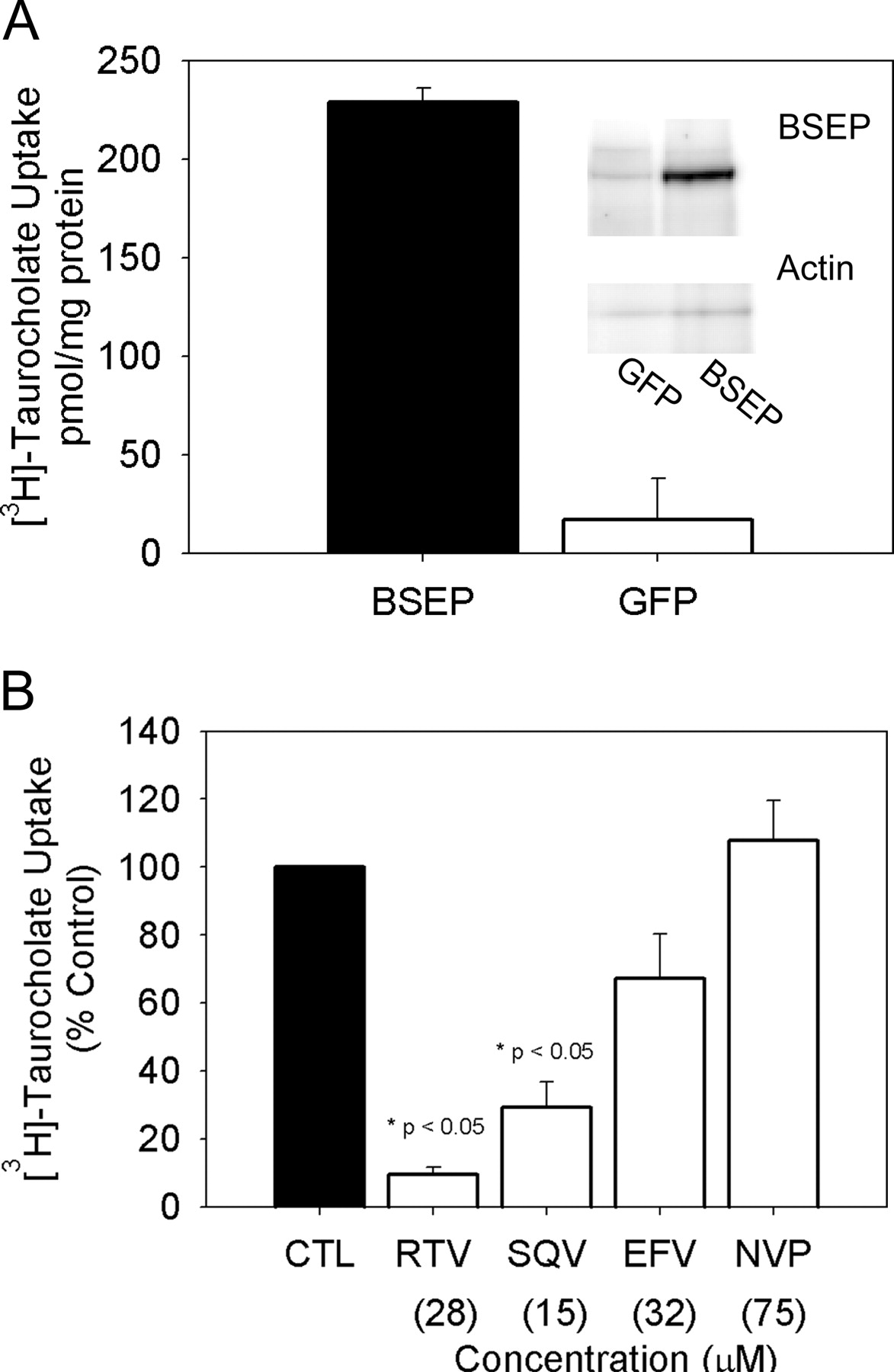
[3H]Taurocholate (1 μM) transport in hBSEP-expressing Sf9 cell-derived membrane vesicles. A, ATP-dependent [3H]taurocholate transport in hBSEP- and GFP-expressing vesicles. Inset, immunoblot of BSEP in hBSEP and GFP-expressing vesicles. B, effects of antiretroviral agents ritonavir (RTV; 28 μM), saquinavir (SQV; 15 μM), efavirenz (EFV; 32 μM), and nevirapine (NVP; 75 μM) on [3H]taurocholate transport expressed as percentage control values. Data were compiled from three separate vesicle preparations and are expressed as mean ± S.E.M.
Oocyte Isolation and Injection. Adult female Xenopus laevis were cooled in an ice bath, anesthetized by immersion in tricaine methanesulfonate, and decapitated. Mature stage V and VI oocytes were isolated and defolliculated by treatment with collagenase A (Roche Diagnostics). Oocytes were maintained at 18°C in oocyte Ringer's buffer (82.5 mM NaCl, 2.5 mM KCl, 1 mM Na2HPO4, 3 mM NaOH, 1 mM CaCl2, 1 mM MgCl2, 1 mM pyruvic acid, and 5 mM HEPES, pH 7.6) supplemented with 0.05 mg/ml gentamicin sulfate, 1.5 mM sodium pyruvate, and 5% heat-inactivated horse serum. Oocytes were injected with 5 ng of human NTCP cRNA 2 days following isolation. Oocytes injected with water (16.1 nl) served as controls. Two days following injection, the uptake of 1 μM [3H]taurocholate at 30 min was evaluated in the presence and absence of ritonavir (0.2–28 μM), saquinavir (15 μM), efavirenz (32 μM), or nevirapine (75 μM). Uptake was terminated by aspiration of the uptake solution followed by three washes in ice-cold oocyte Ringer's buffer. Radioactivity associated with each oocyte was determined by liquid scintillation counting.
Transport Studies Using Recombinant Vaccinia Virus. HeLa cells grown in 12-well plates (∼0.8 × 106 cells/well) were infected with vaccinia (vtf-7) at a multiplicity of infection of 10 plaque-forming units/cell in serum-free Opti-MEM I medium (Invitrogen) and allowed to adsorb for 30 min at 37°C. Cells in each well were transfected with 1 μg of human NTCP or rat Ntcp cDNA packaged into pEF6/V5-His-TOPO vector (Invitrogen), along with Lipofectin (Invitrogen), and incubated at 37°C for 16 h. The parental plasmid lacking any insert was used as control. Transport was evaluated using [3H]taurocholate as outlined previously (Kim et al., 1999). Passive diffusion was determined by carrying out parallel experiments using the parental plasmid DNA lacking the transporter cDNA, and this value was subtracted from the total uptake rate observed in the presence of the transporter cDNA. NTCP or Ntcp-mediated uptake of 5 μM [3H]taurocholate during the initial 3 min of uptake was evaluated in the absence or presence of a drug inhibitor (0.01–100 μM). The extent of inhibition was assessed in terms of an IC50 value (Prism; GraphPad Software, Inc., San Diego, CA). All experiments were carried out in duplicate on at least 2 to 3 experimental days.

Effects of antiretroviral agents on [3H]taurocholate (1 μM) 10-min accumulation in sandwich-cultured human hepatocytes. Ritonavir (A), saquinavir (B), efavirenz (C), and nevirapine (D) data were compiled from two separate human liver preparations; with the exception of nevirapine, the 6.7 and 75 μM concentrations were conducted in only one human liver preparation. These data are expressed as mean ± S.E.M., except for nevirapine as noted above. Black bars, cells + bile; white bars, cells only. BEI values (mean ± S.D.) are expressed across the top of each graph.
Statistics. Parametric testing was performed on data from experiments involving suspended rat hepatocytes and NTCP-injected oocytes. Paired two-tailed Student's t tests were conducted between each treatment group and the appropriate control. Baseline criterion for statistical significance was p < 0.05. For multiple comparisons, the appropriate Bonferroni correction was applied.
Results
BSEP Expressing Sf9-Derived Membrane Vesicles. BSEP-expressing Sf9-derived membrane vesicles were used to determine the effects of antiretroviral agents on BSEP transport protein activity. The average ATP-dependent [3H]taurocholate accumulation in vehicle-treated BSEP-expressing vesicles was 205 ± 59 pmol/mg protein. As shown in Fig. 1B, ritonavir (28 μM) inhibited BSEP-mediated [3H]taurocholate transport by 90%, saquinavir (15 μM) inhibited transport by 71%, and efavirenz (32 μM) inhibited transport by 33%. Nevirapine (75 μM) had no effect on BSEP-mediated [3H]taurocholate transport.

Effects of antiretroviral agents on [3H]taurocholate (1 μM) 10-min accumulation in sandwich-cultured rat hepatocytes. Ritonavir (A), saquinavir (B), efavirenz (C), and nevirapine (D) data were compiled from three separate rat liver preparations and are expressed as mean ± S.E.M. Black bars, cells + bile; white bars, cells only. BEI values (mean ± S.D.) are expressed across the top of each graph.
Sandwich-Cultured Human Hepatocytes. The effect of antiretroviral agents on [3H]taurocholate accumulation in human hepatocytes is shown in Fig. 2. Accumulation of [3H]taurocholate in cells + bile canaliculi decreased in a concentration-dependent manner in ritonavir- and saquinavir-treated groups. At the highest concentrations of each of these drugs (28 μM ritonavir, 15 μM saquinavir), accumulation decreased by 72 and 66%, respectively. Maximal efavirenz (32 μM) exposure resulted in a 37% decrease in accumulation. Nevirapine did not alter accumulation at any concentration examined (0–75 μM). Biliary excretion of [3H]taurocholate was altered by ritonavir and saquinavir and to a lesser extent by efavirenz. The biliary excretion index was decreased by 59% with ritonavir (28 μM), 39% with saquinavir (15 μM), and 20% with efavirenz (32 μM). Nevirapine did not alter the biliary excretion index of [3H]taurocholate.
Sandwich-Cultured Rat Hepatocytes. The effect of antiretroviral agents on [3H]taurocholate disposition in sandwich-cultured rat hepatocytes is shown in Figs. 3 and 4. Ritonavir decreased accumulation in cells + bile canaliculi in a concentration-dependent manner. A 67% decrease in accumulation was observed at the highest concentration of 28 μM. Saquinavir and efavirenz also decreased [3H]taurocholate accumulation over the concentration ranges examined. Saquinavir decreased [3H]taurocholate accumulation by 77% at 15 μM, and efavirenz decreased accumulation by 76% at 32 μM. The biliary excretion index of [3H]taurocholate was completely abolished in rat hepatocytes treated with ritonavir. Saquinavir decreased the biliary excretion index by 96%. However, there was no change in the biliary excretion index with efavirenz (32 μM) or nevirapine (75 μM). Figure 4 demonstrates protein expression of BSEP in cultured human and rat hepatocytes.

Protein expression of BSEP and actin in sandwich-cultured rat and human hepatocytes. Immunoblot analysis of BSEP protein expression was performed in day 4 sandwich-cultured rat hepatocytes (left) and day 6 sandwich-cultured human hepatocytes (right).
Inhibition of Bile Acid Uptake in Isolated Rat Hepatocytes, NTCP-Injected Oocytes, and NTCP-Expressing HeLa Cells. Because the intracellular [3H]taurocholate accumulation data in human and rat sandwich-cultured hepatocytes suggested an inhibition of bile acid uptake processes, further investigations were performed to evaluate antiretroviral effects on [3H]taurocholate uptake by using isolated suspended rat hepatocytes, human NTCP-injected oocytes, and HeLa cells transfected with human and rat NTCP. Figure 5 summarizes the [3H]taurocholate uptake data generated in isolated suspended rat hepatocytes. In the standard Na2+-containing buffer in which Ntcp and Oatp transporters are functional, exposure to ritonavir (28 μM) significantly inhibited initial uptake rates of [3H]taurocholate (0.50 ± 0.13 pmol/s/mg protein) compared with control (1.4 ± 0.26 pmol/s/mg protein; p = 0.0029). At the highest concentrations, saquinavir (0.75 ± 0.12 versus 2.1 ± 1.3 pmol/s/mg protein; p = 0.026) and efavirenz (1.1 ± 0.5 versus 2.3 ± 0.5 pmol/s/mg protein; p = 0.029) also inhibited initial uptake rates, whereas nevirapine (1.8 ± 1.2 versus 2.1 ± 1.2 pmol/s/mg protein; p = 0.394) did not.

Effects of antiretroviral agents on sodium-dependent and -independent [3H]taurocholate (1 μM) uptake in suspended rat hepatocytes. Ritonavir (A), saquinavir (B), efavirenz (C), and nevirapine (D) data were compiled from at least three separate rat liver preparations and are expressed as mean ± S.E.M. Closed circles, control in standard sodium-containing buffer; open circles, control in sodium-free buffer; closed triangles, treatment group in standard sodium-containing buffer; open triangles, treatment group in sodium-free buffer. Statistical comparisons were made for uptake rates between the treatment conditions.
Ritonavir, saquinavir, and efavirenz also significantly inhibited Na2+-independent (Oatp-mediated) initial uptake rates of [3H]taurocholate in suspended hepatocytes. The initial uptake rate of the control group was 0.53 ± 0.10 pmol/s/mg protein. Exposure to ritonavir, saquinavir, and efavirenz decreased the initial uptake rates to 0.13 ± 0.07 (p = 0.002), 0.12 ± 0.09 (p = 0.003), and 0.34 ± 0.03 (p = 0.015) pmol/s/mg protein, respectively. Nevirapine did not alter the [3H]taurocholate uptake rate (0.37 ± 0.17 pmol/s/mg protein; p = 0.120).
In human NTCP-injected oocytes, ritonavir (28 μM) and saquinavir (15 μM) inhibited [3H]taurocholate uptake by 80 and 93%, respectively (Fig. 6). [3H]Taurocholate uptake was diminished by approximately 59% with efavirenz (32 μM). Nevirapine (75 μM) did not alter transport of [3H]taurocholate (Fig. 6).
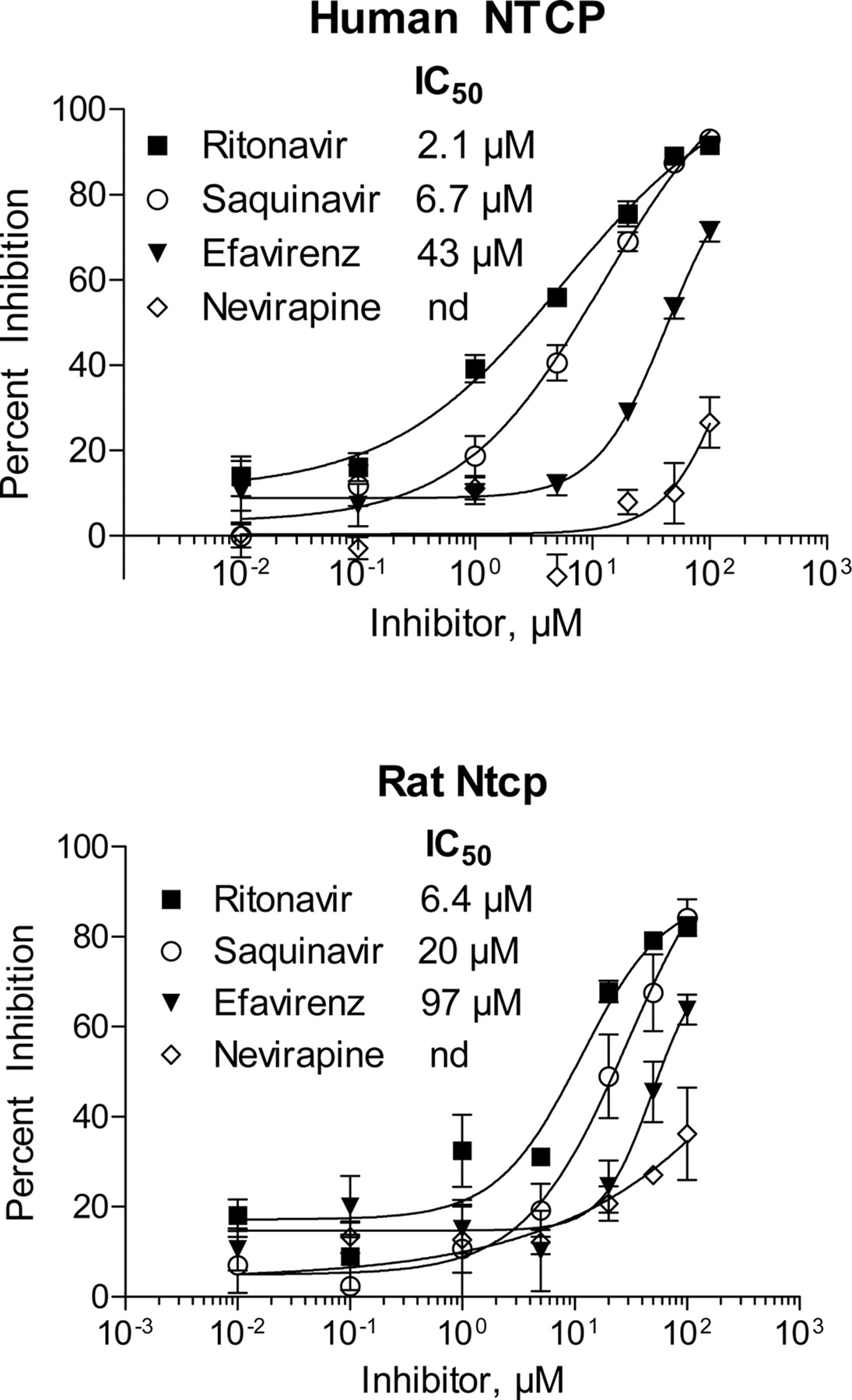
Inhibitory effects of the antiretroviral agents on human NTCP and rat Ntcp-mediated uptake of [3H]taurocholate were further investigated using a recombinant vaccinia expression system in HeLa cells (Fig. 7). IC50 values for human and rat NTCP were calculated for ritonavir (2.1 and 6.4 μM, respectively), saquinavir (6.7 and 20 μM, respectively), and efavirenz (43 and 97 μM, respectively).
Discussion
Strict regulation of intrahepatic bile acid concentrations is considered critical to the maintenance of optimal hepatocyte function. The hepatocyte controls intrahepatic bile acid concentrations by modulating the formation, metabolism, and transport of bile acids in the hepatocyte. In response to elevations in bile acid concentrations, bile acid synthesis and uptake are repressed, and metabolism and excretion are enhanced. Synthesis is repressed by the inhibition of CYP7A1 and CYP8B1 (Hofmann, 1999), and hepatic uptake is repressed by the down-regulation of NTCP and OATP-C (Gartung et al., 1996, 1997). In addition, elevated bile acid concentrations promote bile acid elimination via induction of phase I and II metabolizing enzymes (CYP3A4, UGT2B4, and SULT2A1) and transport proteins (MRP2 and BSEP) (Tu et al., 2000; Eloranta and Kullak-Ublick, 2005). Interference with these homeostatic mechanisms may result in bile-acid induced hepatotoxicity.

Effects of antiretroviral agents on [3H]taurocholate (1 μM) uptake in human NTCP-injected oocytes. Effect of ritonavir (0.28–28 μM), saquinavir (15 μM), efavirenz (32 μM), and nevirapine (75 μM) on [3H]taurocholate transport in human NTCP-injected oocytes. Data represent mean ± S.E.M. (six to eight oocytes) and are expressed as percentage control value for at least three separate experiments.
Drugs that inhibit bile acid transport processes may be associated with cholestasis and/or hepatotoxicity. Meier and colleagues (Stieger et al., 2000; Fattinger et al., 2001) proposed that inhibition of BSEP may increase bile acid concentrations within the hepatocyte. Therefore, assessing the degree of inhibition of BSEP function by xenobiotics may be one approach to identifying drugs with hepatotoxic potential. However, because hepatic bile acid regulation also involves the uptake transporter NTCP, the effects of xenobiotics on the function of this transporter also should be considered. Drugs that inhibit only BSEP may lead to higher intrahepatocyte concentrations of bile acids than drugs that inhibit both NTCP and BSEP.
The aim of the present set of experiments was to examine alterations in bile acid uptake and biliary excretion by selected antiretroviral agents as a potential mechanism of antiretroviral-associated hepatotoxicity. Using multiple model systems, these studies clearly demonstrate inhibition of BSEP-mediated biliary excretion of bile acids and inhibition of NTCP- and OATP-mediated bile acid uptake by ritonavir and saquinavir. Efavirenz modestly inhibited BSEP-mediated biliary excretion of taurocholate in BSEP vesicles and cultured human hepatocytes. Nevirapine did not alter bile acid transport in any experimental systems used.
In the two human systems evaluated (Sf9 cell-derived vesicles and cultured human hepatocytes), the rank order of BSEP inhibition was identical: ritonavir > saquinavir > efavirenz > nevirapine. However, this rank order does not mimic the hepatotoxic potential of these antiretrovirals when used clinically (nevirapine > ritonavir > saquinavir = efavirenz). Nevirapine, although commonly associated with the development of cholestatic (Clarke et al., 2000; Patel et al., 2004) and hepatocellular hepatotoxicity (de Maat et al., 2003; PACTG1022, 2004), did not inhibit the BEI of [3H]taurocholate. Ritonavir-associated hepatotoxicity has been reported in up to 9% of patients (Norvir Product Information, http://www.rxabbott.com/pdf/norpi2a.pdf); however, hepatic toxicity is not generally associated with saquinavir or efavirenz.
Other drugs that are known hepatotoxins (troglitazone and bosentan) have been shown to inhibit biliary excretion of bile acids. These agents also inhibit the hepatic uptake of bile acids (Kostrubsky et al., 2003; Kemp et al., 2005). This was thought to occur as a compensatory mechanism in response to increasing intrahepatic bile acid concentrations resulting from inhibition of biliary excretion. Our in vitro observations also demonstrate inhibition of hepatic uptake of bile acids by ritonavir and saquinavir in human and rat, but once again, the rank order of inhibition of uptake observed does not mimic the clinical hepatotoxic potential of these antiretroviral agents.

Inhibition of human NTCP and rat Ntcp activity in HeLa cells. Inhibitory effects of ritonavir, saquinavir, efavirenz, and nevirapine on NTCP-mediated uptake of [3H]taurocholate (5 μM) were assessed, and IC50 values were determined. Inhibition by nevirapine was not achieved (nd) at the concentration range examined.
The development of hepatotoxicity associated with antiretroviral therapy is complex and likely multifactorial such that inhibition of bile acid transport may be only one of several contributing factors of ritonavir-associated hepatotoxicity. Clearly, the likelihood that inhibition of bile acid transport plays a role in hepatotoxicity associated with nevirapine is minimal. To more clearly understand the consequences of drug-associated inhibition of hepatic bile acid transport, it would be useful to determine whether plasma bile acid concentrations are elevated in patients taking antiretroviral therapy. These investigations are currently being undertaken in our laboratory.
In summary, using four human and three rat models of transport, this work is the first to demonstrate inhibition of BSEP, NTCP, and OATP activity by ritonavir, saquinavir, and efavirenz. Nevirapine did not affect the activities of these transporters in any system. Data generated from this set of experiments suggest that inhibition of BSEP by selected antiretroviral drugs is not the predominant cause of antiretroviral-associated hepatotoxicity. Any inhibition of BSEP appeared to be countered by concurrent inhibition of the uptake transporters NTCP and OATP.
Footnotes
-
This work was supported in part by a 2003 to 2004 developmental grant from the University of North Carolina at Chapel Hill Center for AIDS Research, by a National Institutes of Health-funded program (9P30 AI 50410-04), and by National Institutes of Health Grants GM41935, CA106101, GM54724, GM31304, and AI54980. M.P.M. is the recipient of an American Foundation for Pharmaceutical Education predoctoral fellowship.
-
A portion of this work was previously presented: McRae MP, Brouwer KLR, and Kashuba ADM (2004) Effects of ritonavir and nevirapine on hepatic transport and biliary excretion index of taurocholate in sandwich-cultured primary human and rat hepatocytes. International Society for the Study of Xenobiotics Annual Meeting; 2004 Aug 29–Sept 2; Vancouver, BC, Canada. pp 475, International Society for the Study of Xenobiotics, Washington, DC; and McRae MP, Bourdet DL, Kim RB, Brouwer KLR, and Kashuba ADM (2004) The antiretrovirals ritonavir, saquinavir and efavirenz, but not nevirapine, inhibit taurocholate uptake in NTCP-injected oocytes, PT2337. American Association of Pharmaceutical Scientists Annual Meeting; 2004 Nov 7–11; Baltimore, MD. American Association of Pharmaceutical Scientists, Arlington, VA.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.102657.
-
ABBREVIATIONS: BSEP, bile salt export pump; DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; GFP, green fluorescence protein; HBSS, Hanks' balanced salt solution; BEI, biliary excretion index; NTCP, sodium-dependent taurocholate-cotransporting polypeptide; OATP, organic anion-transporting polypeptide.
- Received February 24, 2006.
- Accepted May 22, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}