


Abstract
Grapefruit juice elevates blood levels of some drugs taken orally, primarily by inhibiting intestinal CYP3A4-mediated first-pass metabolism. Two prominent furanocoumarins in the juice, 6′,7′-dihydroxybergamottin (DHB) and bergamottin (BG), have been demonstrated as important contributors to grapefruit juice-drug interactions. Using CYP3A4-expressing Caco-2 cells and representative probes from distinct CYP3A4 substrate subgroups (midazolam, testosterone), we compared the time-dependent inhibitory properties of DHB and BG. DHB rapidly inhibited CYP3A4 activity in a substrate-independent fashion with maximal inhibition (≥85%) generally occurring within 30 min. In contrast, BG had a slower onset and exhibited substrate-dependent inhibition. Whereas testosterone 6β-hydroxylation was inhibited by >50% with all exposure times (0.5-3 h), midazolam 1′-hydroxylation was unaffected, or even activated, with short exposure times (<1 h). After a 3-h exposure, however, BG had begun to “catch up” with DHB, causing ≥70% inhibition, independent of substrate. Likewise, loss of CYP3A4 protein, believed to reflect rapid intracellular degradation of the enzyme following mechanism-based inactivation, was comparable between the furanocoumarins (40-50%). The time course of BG-mediated inhibition was similar after just a 30-min exposure, indicating that the short exposure presumed to occur after juice ingestion is sufficient to initiate the events required to cause substantial inhibition (≥50%). These results suggest that after ingestion of a glass of grapefruit juice, CYP3A4 is maximally inhibited by DHB before BG has the opportunity to act. However, foods containing BG but not DHB (e.g., lime juice) could produce a substrate-dependent interaction with drugs consumed concomitantly, but a substrate-independent interaction with drugs taken several hours after food consumption.
Although the primary site of inhibition-based metabolic drug-drug interactions is generally the liver, several studies have shown, for drugs taken orally, that the small intestine can serve as a second, presystemic site. In a recent review of the 24 inhibition-based interactions in which intestinal metabolism was implicated as an important contributor to the potential clinical significance (Doherty and Charman, 2002), all involve the prominent drug metabolizing enzyme CYP3A4. This is not surprising, as CYP3A4 is the major cytochrome P450 expressed in epithelial cells of the human small intestinal mucosa (enterocytes), at least in most Caucasians, and it has been implicated in the metabolism of >50% of all drugs in clinical use (Thummel and Wilkinson, 1998). Although a concomitant medication frequently represents the CYP3A4 inhibitory agent, a number of nondrug (rather, nonregulated) agents are also known to inhibit CYP3A4 in vivo and include herbal supplements and dietary substances (Huang and Lesko, 2004). Of these nonregulated agents, grapefruit juice has become one of the most widely recognized. Indeed, grapefruit juice accounted for nearly half (10) of the aforementioned inhibition-based interactions (Doherty and Charman, 2002).
Interactions between drugs and grapefruit juice are unique, in that, when the juice is consumed in usual volumes, inhibition occurs exclusively in the small intestine (Lown et al., 1997; Bailey et al., 1998a). Modes of intestinal CYP3A4 inhibition by grapefruit juice include both reversible and mechanism-based (Bailey et al., 1998a; Guo et al., 2000), as well as destruction of the CYP3A4 protein (Lown et al., 1997). The loss of protein without a corresponding decrease in mRNA is believed to reflect accelerated intracellular degradation following mechanism-based inactivation (Lown et al., 1997). Of the numerous candidate inhibitors identified in grapefruit juice, a group of compounds termed furanocoumarins have been shown to potently inhibit CYP3A4 by at least one of these modes. The two most abundant furanocoumarins, bergamottin (BG) and 6′7′-dihydroxybergamottin (DHB), inhibit CYP3A4 activity in both a reversible and irreversible manner (Fukuda et al., 1997; Schmiedlin-Ren et al., 1997a; He et al., 1998; Guo et al., 2000; Tassaneeyakul et al., 2000; Paine et al., 2004). Human volunteer studies with lime juice (which contained BG but undetectable DHB) (Bailey et al., 2003) and an aqueous extract of grapefruit juice (which contained DHB and extremely low concentrations BG or other furanocoumarins) (Kakar et al., 2004) indicated that both furanocoumarins contribute to the “grapefruit juice effect” in vivo.
In a recent systematic comparison of BG and DHB as inhibitors of intestinal CYP3A4 activity, we demonstrated, using human intestinal microsomes, that the two furanocoumarins differ markedly in inhibitory kinetic and binding properties (Paine et al., 2004). As reversible inhibitors, whereas DHB effectively inhibited the hydroxylation of both testosterone and midazolam (apparent Ki ∼0.8 μM), which are representative of two distinct CYP3A4 substrate subgroups (Kenworthy et al., 1999; Schrag and Wienkers, 2001; Galetin et al., 2003), BG was eight times more effective in inhibiting testosterone than midazolam hydroxylation (ap-parent Ki of 1.6 and 13 μM, respectively). In contrast to their behavior as reversible inhibitors, both DHB and BG were found to be substrate-independent mechanism-based inhibitors with comparable potency (KI ∼25 μM; kinact ∼0.35 min-1).
If mechanism-based inhibition by each furanocoumarin occurs rapidly in vivo, these findings suggest that the substrate-dependent reversible inhibition by BG is clinically irrelevant, and the effects of the two furanocoumarins in vivo should be similar whether the interacting drug belongs to the testosterone or midazolam subgroup. Extrapolating these in vitro data to the in vivo situation is not straightforward, however, as BG is 1000 times more lipophilic than DHB (log P values of 5.6 and 2.6, respectively) (SciFinder Scholar, v2002; American Chemical Society, Columbus, OH). This marked disparity in lipid solubility infers that these furanocoumarins may differ in their rates of entry into enterocytes following ingestion of grapefruit juice. In addition, the two furanocoumarins would be expected to differ in their rates of exit from the basolateral (vascular) side of enterocytes, where serum proteins reside. These factors, combined with a difference in the unbound fraction within enterocytes, could lead to time-dependent differences in the concentration of each furanocoumarin at the enzyme active site, which could in turn lead to a difference in time to onset of inhibition. An additional issue in extrapolating data obtained from micro-somes or recombinant enzymes is that furanocoumarin-mediated loss of enterocyte CYP3A4 protein does not occur in isolated enzyme systems (Edwards et al., 1999).
These observations led us to systematically compare the disposition and CYP3A4 inhibitory properties of BG and DHB using CYP3A4-expressing Caco-2 cell monolayers. In addition to CYP3A4, this modified human intestinal cell line expresses potentially relevant transporters indicative of enterocytes (e.g., P-glycoprotein and multidrug resistance associated proteins 1-5) (Pfrunder et al., 2003). Not only have these cells been shown to serve as a useful model to study the intestinal first-pass extraction of drugs (midazolam, indinavir, sirolimus, and saquinavir) (Fisher et al., 1999b; Hochman et al., 2000; Paine et al., 2002; Mouly et al., 2004), but also drug interactions at the level of the intestine (DHBmidazolam and ketoconazole-midazolam) (Schmiedlin-Ren et al., 1997a; Gibbs et al., 2000). We report that BG and DHB differ considerably in rates of entry and time to onset of inhibitory effect and that substrate-dependent inhibition by BG can be demonstrated in this intact model of enterocytes.
Materials and Methods
Materials and Chemicals
Cell culture materials and media ingredients [culture inserts, murine laminin, Dulbecco's modified Eagle's medium (DMEM), non-essential amino acids, fetal bovine serum, dl-α-tocopherol, zinc sul-fate, sodium selenite, gentamicin, 1α,25-dihydroxyvitamin D3 (1α,25-(OH)2-D3), and all-trans-retinoic acid] were purchased from sources as previously described (Mouly et al., 2004). Testosterone, 6β-hydroxytestosterone (6β-OH TST), cortisol, midazolam maleate, alprazolam, NADPH, and human serum albumin (fraction V) were purchased from Sigma-Aldrich (St. Louis, MO). DHB was a gift from the Florida Department of Citrus (Lakeland, FL). BG and psoralen were purchased from Indofine Chemical Co. (Hillsborough, NJ). 1′-Hydroxymidazolam (1′-OH MDZ) was purchased from BD Gentest (Woburn, MA). Complete Mini protease inhibitor was purchased from F. Hoffmann-La Roche (Nutley, NJ). Jejunal microsomes were previously prepared from mucosal scrapings from four human donor small intestines (Paine et al., 1997); none of the preparations contained readily detectable CYP3A5 immunoreactive protein using a selective antibody to CYP3A5 (BD Gentest). Western blotting materials and reagents [sodium dodecyl sulfate, acrylamide/bis (37.5:1), ammonium persulfate, TEMED, polyvinylidene difluoride membrane, and enhanced chemiluminescence reagents] were purchased from sources as previously described (Paine et al., 2002). A murine monoclonal antibody that recognizes all human CYP3A isoforms, termed 13-7-10, was a gift from Dr. Pierre Kremers (Universite de Liege, Liege, Belgium). A murine monoclonal antibody raised against human villin was purchased from Chemicon International (Temecula, CA). The secondary antibody, rabbit anti-mouse horseradish peroxidase-conjugated IgG, was purchased from Zymed Laboratories (South San Francisco, CA). All other chemicals and reagents were of tissue culture, electrophoresis, or HPLC grade where appropriate.
Caco-2 Cell Experiments
Cell Culture Conditions. The Caco-2 cell clone P27.7 (Schmiedlin-Ren et al., 1997b) (passage 24-33) was seeded onto laminin-coated culture inserts (4.2-cm2 growth area), at a density of ∼5 × 105 cell/cm2, as previously described (Mouly et al., 2004). After reaching confluence (∼5 days), the cell cultures were maintained in differentiation medium (Mouly et al., 2004) for 3 weeks. Except for the disposition and apparent permeability coefficient (Papp) determination studies with BG and DHB (preliminary studies with 1α,25-(OH)2-D3-treated cells produced nearly identical results with untreated cells), differentiation medium was supplemented with 1α,25-(OH)2-D3 (a CYP3A4-inducing agent) and all-trans-retinoic acid as previously described (Mouly et al., 2004). Cell cultures for a given experiment were maintained throughout the ∼4-week period in a 37°C, humidified incubator with a 5% carbon dioxide air atmosphere.
Time Course Studies with DHB and BG. Warm incubation medium (differentiation medium devoid of fetal bovine serum and 1α, 25-,(OH)2-D3 and was spiked with DHB or BG to yield a final concentration of 30 μM (0.3% methanol). For apical-to-basolateral (A→ B) translocation, DHB- or BG-containing incubation medium (1.5 ml) was added to the apical compartment, followed by plain incubation medium (1.5 ml), supplemented with or without human serum albumin (HSA) (4%) to the basolateral compartment of culture inserts. For basolateral-to-apical (B→ A) translocation, plain incubation medium was added to the apical compartment, followed by DHB- or BG-containing incubation medium to the basolateral compartment. After 0, 0.25, 0.5, 1, 2, 3, or 4 h at 37°C, apical and basolateral media were collected and placed on dry ice. The remaining cell monolayers were quickly washed with cold DMEM, then scraped into cold incubation medium (0.4 ml), and placed on dry ice. All collections were stored at -80°C pending analysis for DHB or BG. After thawing each sample, internal standard (psoralen) was added to a volume of 0.4 to 0.5 ml, which was brought up to 1 ml with DMEM. A standard curve was prepared similarly using known amounts of DHB or BG (0.25-15 nmol). The unknowns and standards were further processed and quantified for DHB or BG by HPLC as previously described (Paine et al., 2004).
Apparent Permeability Coefficient of DHB and BG. To determine the Papp of each furanocoumarin in the A→ B direction, DHB or BG (30 μM) was added to the apical compartment, followed by plain incubation medium (with or without HSA) to the basolateral compartment of at least triplicate culture inserts. For the B→ A direction, plain incubation medium was added to the apical compartment, followed by DHB or BG (without HSA) to the basolateral compartment. After 10 or 30 min (at which <10% of the initial concentration of DHB or BG, respectively, had translocated to the receiver compartment), apical and basolateral media and remaining cell monolayers were collected and analyzed for DHB or BG as described for the time course studies.
Inhibition of CYP3A4 Catalytic Activity by DHB and BG.Concentration-dependent inhibition (IC50 determination). Incubation medium containing DHB or BG and CYP3A4 substrate (midazolam or testosterone) was added to the apical compartment, followed by plain incubation medium, supplemented with or without HSA (4%), to the basolateral compartment of culture inserts. Final concentrations of DHB and BG ranged from 0 to 10 and 0 to 30 μM, respectively (1% methanol). Final concentrations of midazolam and testosterone were 8 μM (0.2% dimethyl sulfoxide) and 250 μM (1% methanol), respectively. After 30 min at 37°C, apical and basolateral media were collected, placed on dry ice, then stored at -80°C pending analysis for 1′-OH MDZ or 6β-OH TST. After thawing, internal standard (alprazolam or cortisol) was added to 0.4 to 0.5 ml of sample, which was brought up to 1 ml with DMEM. A standard curve was prepared similarly using known amounts of 1′-OH MDZ (1-150 pmol) or 6β-OH TST (0.1-5.0 nmol). The unknowns and standards were further processed and quantified for 1′-OH MDZ and 6β-OH TST by liquid chromatography/mass spectrometry and HPLC, respectively, as previously described (Paine et al., 2004).
Time-dependent inhibition. Incubation medium containing DHB or BG (30 μM) was added to the apical compartment, followed by plain incubation medium, supplemented with or without HSA, to the basolateral compartment of culture inserts. After 0, 0.5, 1, or 3 h, apical and basolateral media were aspirated. After washing the cells with warm DMEM, midazolam (8 μM) or testosterone (250 μM) was added to the apical compartment, followed by plain incubation medium to the basolateral compartment. After 1 h, apical and basolateral media were collected and processed as described for the preceding experiment. The remaining cell monolayers were washed with cold DMEM and scraped into 0.5 ml of cold lysis buffer [0.5% Triton X-100, 0.5 mM EDTA, 0.15 M NaCl, and 8 mM N-tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid] containing protease inhibitors. The cell lysates were placed on ice for 1 h, centrifuged at 16,000g for 10 min, and the supernatant was collected and stored at -80°C pending analysis for CYP3A4 and villin immunoreactive protein by Western blot (described below).
Time-Dependent Inhibition of CYP3A4 Catalytic Activity following a Short Exposure to DHB and BG. Incubation medium containing DHB or BG (30 μM) was added to the apical compartment, followed by plain incubation medium (with or without HSA) to the basolateral compartment. After 30 min, apical and basolateral media were aspirated, the cells were washed, and plain incubation medium was added to both compartments (with or without HSA in the basolateral compartment). After 0, 1, 2.5, or 4 h, the media were aspirated, and substrate (8 μM midazolam or 250 μM testosterone) was added to the apical compartment, followed by plain incubation medium (with or without HSA) to the basolateral compartment. After 1 h, apical and basolateral media and remaining cell monolayers were collected in the same manner as described for the previous experiments.
Western Blot Analysis of Cell Lysates for CYP3A4. Total protein concentration of the cell lysates was determined by the method of Lowry et al. (1951) using bovine serum albumin as the reference standard. Cell lysates were diluted in sample buffer, then 20 μg of protein were electrophoretically separated on 0.1% SDS/9% polyacrylamide gels, and transferred to polyvinylidene difluoride membranes as previously described (Paine et al., 2002). The membranes were probed for CYP3A4 and the control protein villin using the appropriate primary and secondary antibodies as described (Paine et al., 2002). Although the primary antibody used has been reported to detect all human CYP3A isoforms, it was previously shown not to detect CYP3A5 in these modified Caco-2 cells; this was likely due to the very low level of expression of CYP3A5 relative to CYP3A4 (Schmiedlin-Ren et al., 1997b). CYP3A4 and villin immunoreactive protein were visualized by enhanced chemiluminescence using the Chemi-Doc imaging system (Bio-Rad, Hercules, CA). Densitometry was performed with the Bio-Rad software program Quantity One (v4.1). CYP3A4 protein content was expressed as the ratio of the integrated optical density (IOD) of CYP3A4 to that of villin.
Human Jejunal Microsomal Incubations
BG was dissolved as 100-fold concentrated solutions and midazolam as a 1000-fold concentrated solutions in methanol. Incubation mixtures consisted of 0.1 mg/ml microsomal protein, BG, midazolam, and potassium phosphate buffer (0.1 M, pH 7.4). The mixtures were equilibrated in a shaking water bath at 37°C for 5 min before initiating the reactions with NADPH (final concentration, 1 mM) to yield a final volume of 0.5 ml. Final concentrations of BG, midazolam, and methanol (v/v) were 0 to 10 μM, 4 μM, and 1.1%, respectively. Reactions were terminated after 4 min with 1-ml ice-cold acetonitrile. The mixtures were processed and analyzed for 1′-OH MDZ by liquid chromatography/mass spectrometry as previously described (Paine et al., 2004). For all microsomal preparations, the amount of 1′-OH MDZ formed was linear with respect to incubation time and amount of microsomal protein.
Data Analysis
Apparent Permeability Coefficient of DHB and BG. The Papp of each furanocoumarin was calculated according to the traditional equation: Papp = (dQ/dt) × (1/AC0), where dQ/dt denotes the flux (centimeter per second) of the furanocoumarin from the dosing to receiving compartment under sink conditions (i.e., before >10% of the initial dose translocated to the receiving compartment), A denotes the surface area of the culture insert (4.2 cm2), and C0 denotes the concentration of furanocoumarin in the dosing compartment at time 0.
IC50 Determination for DHB and BG toward CYP3A4 Activity. The formation rate of 1′-OH MDZ or 6β-OH TST, as percent of control (vehicle), in the presence of increasing concentrations of DHB or BG was plotted against the natural logarithm of the inhibitor concentration. The concentration of inhibitor corresponding to 50% of the control activity (IC50) was calculated by linear regression.
Statistical Analysis. All statistical tests were performed using Statview (v5.01; SAS Institute Inc., Cary, NC). Data are presented, where appropriate, as means ± S.D. of at least three culture inserts. Comparisons between Papp values were made using the Student's unpaired t test (α = 0.05). Comparisons among the extents of CYP3A4 inhibition by DHB or BG after different exposure times were made using 1-way analysis of variance, followed by the Student-Newman-Keuls (for equal sample sizes) or Scheffe's (for unequal sample sizes) post hoc test when an overall significant difference (p value <0.05) ensued.
Results
Disposition of DHB and BG
In the absence of HSA in the basolateral compartment, ≥90% of the DHB dose was recovered throughout the time course, whether the dose was applied to the apical or baso-lateral compartment. Recovery of the BG dose following apical dosing was similar at early time points (0-2 h) but was decreased at the later time points (to ∼70% by 4 h). Recovery of the BG dose following basolateral dosing continuously decreased over time (to ∼40% by 4 h). These incomplete recoveries were attributed to nonspecific binding by BG to the inserts and/or wells rather than the laminin coating the inserts, as studies repeated with Caco-2 cells cultured on uncoated (i.e., laminin-free) inserts produced identical results (not shown). Therefore, to compare the disposition profiles of DHB and BG in the Caco-2 cell system, the amount of each furanocoumarin measured in the apical, basolateral, or cellular compartment was normalized to the total amount recovered.
Whether applied to the apical (Fig. 1A) or basolateral (not shown) compartment, DHB rapidly diffused across the cell monolayer such that, by 4 h, ∼50% of the total amount recovered resided in the receiver compartment. Relatively constant amounts of DHB were measured in the cell scrapings throughout the time course (4-5 and 2-3% of the total amount recovered following apical and basolateral dosing, respectively). The Papp of DHB in the A→ B direction (3.0 ± 0.3 × 10-5 cm/s) was similar to that in the B→ A direction (3.2 ± 0.2 × 10-5 cm/s) (p = 0.17). The presence of HSA in the basolateral compartment accelerated the A→ B translocation of DHB such that, by 1 h, ∼50% of the total amount recovered resided in the basolateral compartment (Fig. 1B). By 4 h, >90% of the dose was recovered from the basolateral compartment. The amount measured in cell scrapings was relatively constant (∼3%) up to 30 min, then continually declined for the remainder of the time course (to <0.5% by 4 h). The A→ B Papp approximately doubled (to 6.4 ± 0.6 × 10-5 cm/s) (p < 0.001).

Time course of disposition of DHB and BG in modified Caco-2 cells following application (30 μM) to the apical compartment. Incubation medium in the basolateral compartment was devoid of (A and C) or supplemented with (B and D) HSA (4%). For BG, in the absence of basolateral HSA (C), the amount detected in the baso-lateral compartment was below the limit of quantification (0.25 nmol).
In contrast to DHB, whether dosed apically (Fig. 1C) or basolaterally (not shown), BG was below the limit of quantification (0.25 nmol) in the receiver compartment throughout the time course. Consequently, neither the A→ B nor B→ A Papp for BG could be determined. The amount of BG measured in cell scrapings increased with time following either route of administration. By 4 h, approximately 60 and 40% of the total amount recovered was measured in the cell scrapings following apical (Fig. 1C) and basolateral dosing, respectively. The presence of HSA in the basolateral compartment improved the total recovery of apically applied BG at the later time points to nearly 100%. Basolateral HSA facilitated the A→ B translocation of BG such that BG was easily detected in the basolateral compartment at all time points (Fig. 1D). By 4 h, 55% of the dose had translocated to the basolateral compartment. The amount of BG measured in cell scrapings increased with time to 2 h (from 2 to ∼15%), at which an apparent steady state was achieved (Fig. 1D). Basolateral HSA enabled the calculation of the A→ BPapp for BG (0.74 ± 0.03 × 10-5 cm/s).
Comparison of DHB and BG as Inhibitors of CYP3A4
For all ensuing experiments, to mimic the inhibitory effect of grapefruit juice toward intestinal CYP3A4 catalytic activity and protein content, DHB and BG were applied only to the apical compartment of culture inserts. In addition, the incubation periods with each furanocoumarin were at most 3 h. The effects of nonspecific binding by BG to the culture inserts and/or wells (in the absence of basolateral HSA) were therefore assumed to be minimal.
IC50 Determination. With DHB, using midazolam as the substrate, in the absence of basolateral HSA, 1′-OH MDZ formation decreased with increasing concentrations of DHB (Fig. 2A). The presence of basolateral HSA attenuated the inhibition of 1′-OH MDZ formation at all concentrations of DHB (Fig. 2B). The r2 values for the linear regression of the natural logarithm of the average percent control activity versus [DHB] were >0.99, yielding IC50 values of 2.4 and 5.4 μM, respectively, in the absence and presence of basolateral HSA. In contrast to DHB, 1′-OH MDZ formation increased with increasing concentrations of BG up to 10 μM in both the absence (Fig. 2C) and presence (Fig. 2D) of basolateral HSA and remained relatively constant above 10 μM. As such, an IC50 could not be determined. This apparent activation of CYP3A4 activity was also observed with three of the four human jejunal microsomal preparations at BG concentrations of 1 to 2.5 μM (Fig. 3). Using testosterone as the substrate, in the absence of basolateral HSA, 6β-OH TST formation decreased with increasing concentrations of DHB (Fig. 2E). The presence of basolateral HSA had a minimal effect on the inhibition of 6β-OH TST formation (Fig. 2F). The r2 values for the linear regression of the natural logarithm of the average percent control activity versus [DHB] were ≥0.96, yielding IC50 values of 1.0 and 0.7 μM, respectively, in the absence and presence of basolateral HSA. Although BG inhibited testosterone 6β-hydroxylation in the absence of basolateral HSA, the average percent inhibition never exceeded 30% (Fig. 2G), even at concentrations up to 100 μM (not shown). Similarly, in the presence of basolateral HSA, the percent inhibition never exceeded 25% (Fig. 2H). IC50 values therefore could not be determined. Activation of testosterone 6β-hydroxylation by BG was not evident.

Concentration-dependent inhibition of midazolam 1′-hydroxylation and testosterone 6β-hydroxylation by DHB and BG in modified Caco-2 cells. Substrate (8 μM midazolam, 250 μM testosterone) and furanocoumarin were coapplied to the apical compartment and incubated for 30 min. Incubation medium in the basolateral compartment was devoid of (A, C, E, and G) or supplemented with (B, D, F, and H) HSA (4%). Symbols and error bars denote the means and S.D.s, respectively, of three to five culture inserts.

Apparent activation of midazolam 1′-hydroxylation by BG in (three of four) human intestinal microsomal preparations, none of which contained readily detectable immunoreactive CYP3A5 protein. Each symbol denotes the mean of duplicate incubations. HI, human intestine.
Onset of Inhibition of CYP3A4 Catalytic Activity. Using midazolam as substrate, in the absence of basolateral HSA, DHB had both a greater and more rapid inhibitory effect on 1′-OH MDZ formation compared with BG (Fig. 4A). With a preincubation time of <1 min (time 0), relative to vehicle, DHB inhibited activity (by 60%), whereas BG had no effect. With a 0.5-h preincubation, DHB inhibited activity by 85% and BG by only 15%. With a 1-h preincubation, DHB inhibited activity by 95% and BG by 60%. With a 3-h preincubation, DHB showed no further inhibition whereas BG inhibited by 90%. The presence of basolateral HSA had no effect on inhibition by DHB with the shorter preincubation times (0 and 0.5 h) and slightly attenuated the inhibitory effect with the longer times (Fig. 4B). In contrast, basolateral HSA prevented inhibition by BG following a 0.5- and 1-h preincubation and attenuated the inhibitory effect following a 3-h preincubation. Using testosterone as substrate, with all preincubation times, in the absence of basolateral HSA, both furanocoumarins inhibited CYP3A4 catalytic activity to a greater extent than with midazolam (Fig. 4, C versus A). However, DHB again had both a greater and more rapid inhibitory effect on CYP3A4 activity compared with BG (Fig. 4C). With a 0-h preincubation, DHB inhibited 6β-OH TST formation by 80%, whereas BG had no effect. Whereas the maximum inhibitory effect of DHB (95%) appeared to occur within 0.5 h, the effect of BG progressively increased the longer the preincubation time (by 60, 80, and >95% with 0.5, 1, and 3 h, respectively). The presence of basolateral HSA attenuated the inhibitory effect of DHB with all preincubations times, but again the maximum effect (>85%) occurred within 0.5 h (Fig. 4D). The inhibitory effect of BG was also attenuated but progressively increased the longer the preincubation time (by 60, 65, and 75% with 0.5, 1, and 3 h, respectively).

Time-dependent inhibition of midazolam 1′-hydroxylation and testosterone 6β-hydroxylation in modified Caco-2 cells following continual exposure to DHB or BG (30 μM). DHB or BG was applied to the apical compartment and incubated for the indicated times. After removing the furanocoumarin, substrate (8 μM midazolam, 250 μM testosterone) was added alone to the apical compartment and incubated for 1 h. Incubation medium in the basolateral compartment was devoid of (A and C) or supplemented with (B and D) HSA (4%) throughout the experiment. Bars and error bars denote the means and S.D.s, respectively, of three to four culture inserts. *, p < 0.05 (versus time 0); **, p < 0.01 (versus time 0); ***, p < 0.001 (versus time 0).
Onset of CYP3A4 Protein Loss. Cell lysates from the preincubation studies were analyzed for villin and CYP3A4 immunoreactive protein by Western blot. In the absence of basolateral HSA, with a 0.5-h preincubation, compared with vehicle, DHB decreased average CYP3A4 protein content (by 30%), whereas BG had no effect (Fig. 5, A and C). With a 1-h preincubation, DHB further decreased average CYP3A4 content (by 40%), whereas BG had only a modest effect (15%). However, with a 3-h preincubation, both furanocoumarins caused a similar loss (∼50%) in average CYP3A4 content. The presence of basolateral HSA attenuated CYP3A4 protein loss (by 10-20%) by both furanocoumarins with all preincubation times (Fig. 5, B and D).

Time-dependent loss of CYP3A4 immunoreactive protein in modified Caco-2 cells following continual exposure to DHB or BG (30 μM). Cell monolayers were harvested for Western blot analysis after incubation with substrate (see Fig. 4). A representative blot probed for villin (control protein) and CYP3A4 is shown (A and B). For each of three to four blots, CYP3A4 protein content (CYP3A4/villin IOD ratio) in lysates prepared from furanocoumarin-treated cells was normalized to that in vehicle-treated cells; the values obtained from all blots were averaged for each exposure time (0, 0.5, 1, and 3 h) (C and D). Incubation medium in the basolateral compartment was devoid of (A and C) or supplemented with (B and D) HSA (4%) throughout the experiment. Bars and error bars denote means and S.D.s, respectively, of three to four culture inserts. V, vehicle; D, DHB; B, BG. *, p < 0.05 (versus time 0); **, p < 0.01 (versus time 0); ***, p < 0.001 (versus time 0).
Time-Dependent Effects of Short-Term Exposure to DHB and BG
CYP3A4 Catalytic Activity. Following consumption of a single glass of grapefruit juice, it seemed reasonable to assume that the exposure of enterocytes to BG or DHB occurs during the finite period of juice absorption. As such, the effects of a short-term (30-min) exposure of Caco-2 cells to each furanocoumarin were examined. Immediately following exposure to DHB, in the absence of basolateral HSA, midazolam 1′-hydroxylation was inhibited by 90% (Fig. 6A). This pronounced inhibitory effect was sustained for as long as 4 h following removal of DHB, during which CYP3A4 activity remained inhibited by ∼85%. In contrast, immediately following exposure to BG, midazolam 1′-hydroxylase activity was unaffected. However, 1 h following the 30-min exposure to BG, activity was inhibited by 60% and was inhibited to an even greater extent (∼80%) with a longer time period (≥2.5 h) following exposure to BG. In the presence of basolateral HSA, midazolam 1′-hydroxylation was again inhibited to a large extent (∼80%) immediately following exposure of the cells to DHB (Fig. 6B), and the inhibitory effect was attenuated with longer time periods following removal of DHB (by 10-20% with a 1- to 4-h time period, respectively). The inhibitory effect of BG was attenuated in the presence of baso-lateral HSA with all time periods ≥1 h following BG removal (by ∼50%). Using testosterone as the substrate, in the absence of HSA, exposure of the cells to both furanocoumarins led to a greater inhibitory effect on CYP3A4 activity compared with the inhibitory effect using midazolam after all time periods following removal of the furanocoumarin (Fig. 6, C versus A). As with midazolam 1′-hydroxylation, however, testosterone 6β-hydroxylation was markedly inhibited immediately following exposure of the cells to DHB (by 95%), and this extent of inhibition (≥90%) remained for up to 4 h following removal of the furanocoumarin (Fig. 6C). Unlike with midazolam 1′-hydroxylation, testosterone 6β-hydroxylation was effectively inhibited (65%) immediately following the 30-min exposure to BG. This extent of inhibition was greater 1 h after removal of BG (80%). As observed with midazolam 1′-hydroxylation, the extent of inhibition of testosterone 6β-hydroxylation was even greater 2.5 h after removal of BG and appeared sustained up to 4 h. In the presence of basolateral HSA, immediately following exposure to DHB, the extent of inhibition of testosterone 6β-hydroxylation (Fig. 6D) was similar to that in the absence of HSA (∼95%). However, ≥1 h after DHB removal, basolateral HSA slightly attenuated the inhibitory effect (by ∼10%). Immediately following exposure to BG, as in the absence of basolateral HSA, testosterone 6β-hydroxylation was effectively inhibited (by 75%) (Fig. 6D). With time periods ≥1 h after BG removal, basolateral HSA attenuated the inhibitory effect by 20-30%.
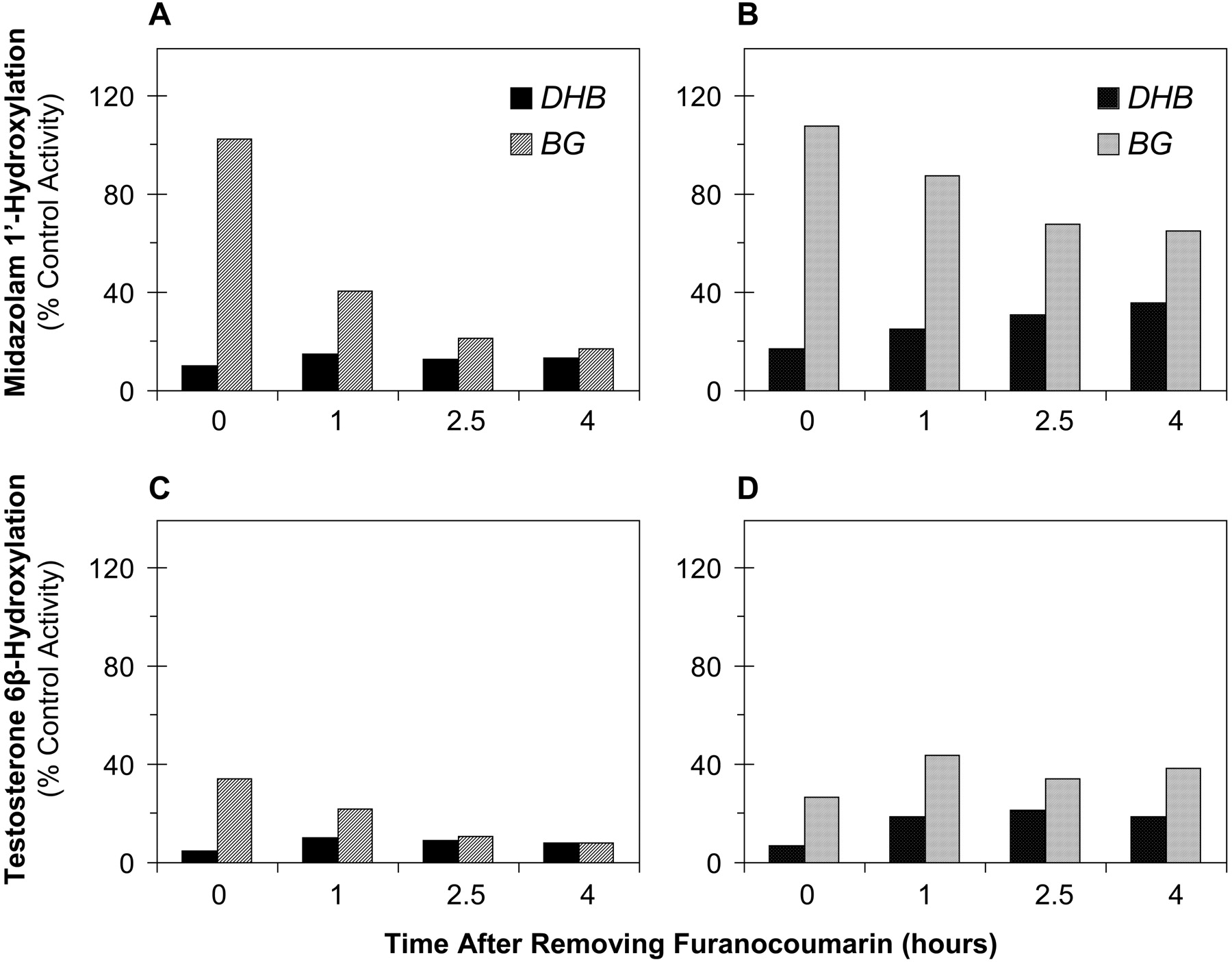
Time-dependent inhibition of midazolam 1′-hydroxylation and testosterone 6β-hydroxylation in modified Caco-2 cells following short-term exposure to DHB or BG (30 μM). DHB or BG was applied to the apical compartment and incubated for 30 min. After removing the furanocoumarin, plain incubation medium was added and incubated for the indicated times. Substrate (8 μM midazolam, 250 μM testosterone) was then added alone to the apical compartment and incubated for 1 h. Incubation medium in the basolateral compartment was devoid of (A and C) or supplemented with (B and D) HSA (4%) throughout the experiment. Bars denote the means of duplicate culture inserts.
CYP3A4 Immunoreactive Protein. The time course of CYP3A4 protein loss following the 30-min exposure to either furanocoumarin (Fig. 7) was similar to that following continuous exposure (Fig. 5).

Time-dependent loss of CYP3A4 immunoreactive protein in modified Caco-2 cells following short-term exposure to DHB or BG (30 μM). Cell monolayers were harvested for Western blot analysis after incubation with substrate (see Fig. 6). A representative blot probed for villin (control protein) and CYP3A4 is shown (A and B). For each of two blots, CYP3A4 protein content (CYP3A4/villin IOD ratio) in lysates prepared from furanocoumarin-treated cells was normalized to that in vehicle-treated cells; the values obtained from the two blots were averaged for each exposure time (0, 1, 2.5, and 4 h) (C and D). Incubation medium in the basolateral compartment was devoid of (A) or supplemented with (B) HSA (4%) throughout the experiment. Bars denote the means of duplicate culture inserts. V, vehicle; D, DHB; B, BG.
Discussion
Human studies have demonstrated that two prominent furanocoumarins contained in grapefruit juice, DHB and BG, are important contributors to grapefruit juice-drug interactions (Bailey et al., 1998b, 2000, 2003; Kakar et al., 2004). Using human intestinal microsomes, we recently reported that BG, but not DHB, acts as a substrate-dependent reversible inhibitor of CYP3A4 (Paine et al., 2004). The in vivo relevance of the substrate dependence of BG was unclear, however, as both furanocoumarins rapidly inactivated the enzyme in a substrate-independent fashion. In addition, the 1000-fold difference in lipid solubility (BG > DHB) might be associated with a time-dependent difference in the concentration of each inhibitor near the enzyme within enterocytes. We therefore compared the time-dependent inhibitory effects of each furanocoumarin using CYP3A4-expressing Caco-2 cells as a model of the human small intestinal epithelial layer.
As predicted, DHB and BG differed markedly in their disposition properties in Caco-2 cells. In the absence of baso-lateral HSA, the superimposable time course profiles and similar Papp for DHB with apical and basolateral dosing indicated that DHB was absorbed across the cell monolayer by passive diffusion. In contrast, BG was never recovered in the receiver compartment (Fig. 1, C versus A), thus its means of absorption could not be ascertained. Whereas steady state cellular levels of DHB were rapidly achieved (within <1 min), cellular levels of BG continually increased over time. The presence of a physiologically relevant concentration of HSA in the basolateral compartment facilitated the A→ B translocation of DHB and BG. This, coupled with lowered cellular levels, suggests that both furanocoumarins bind to HSA. These findings support the contention that, if DHB and BG are absorbed into the vasculature, binding to plasma proteins could explain in part why grapefruit juice does not inhibit hepatic CYP3A4 in vivo (when consumed in usual volumes).
DHB and BG also differed in the concentration-dependent ability to inhibit CYP3A4 catalytic activity. The IC50 values determined for DHB (∼1-5 μM), using testosterone and midazolam as substrate, are at or near the low end of the range of concentrations reported for various brands of grapefruit juice (∼1-60 μM) (Schmiedlin-Ren et al., 1997a; Fukuda et al., 2000; Guo et al., 2000), supporting a role for reversible CYP3A4 inhibition by DHB in grapefruit-drug interactions. With midazolam as substrate, the ∼2-fold higher IC50 for DHB in the presence of HSA likely reflected the 2-fold higher Papp and hence, lower cellular content of DHB (Fig. 1, B versus A). [Cellular midazolam content was minimally reduced by basolateral HSA over a 30-min time interval (Fisher et al., 1999a).] With testosterone as substrate, the lack of an effect of HSA on the IC50 for DHB could have reflected the 2- to 3-fold reduction in cellular testosterone content in the presence of HSA (data not shown), thereby offsetting the reduction in cellular DHB content.
In contrast to DHB, an IC50 for the more lipophilic BG could not be determined; however, consistent with our recent findings with human intestinal microsomes (Paine et al., 2004), substrate-dependent inhibition by BG was evident at concentrations of 10 and 30 μM. Whereas testosterone hydroxylation was inhibited, midazolam hydroxylation was increased (especially in the presence of HSA) (Fig. 2, G and H versus C and D). The apparent activation of midazolam hydroxylation was not an artifact of the cell system, as BG-mediated activation also occurred in incubations with three of four human intestinal microsomal preparations. The one preparation (HI-21) that did not exhibit activation is unexplained, but it was the same preparation used in our recent work (Paine et al., 2004), which accounts for why this was not observed previously. In an earlier study using fresh human hepatocytes and testosterone as substrate, an IC50 of ∼1 μM was reported (Wen et al., 2002). The discrepancy between this and the current results could have been due to the different solvents used to dissolve the BG powder (dimethyl sulfoxide and methanol, respectively) and/or to differences between cells of hepatic and intestinal origin.
As anticipated, based on lipid solubility, DHB and BG differed in the time to onset of CYP3A4 inhibition. For DHB, the rapid onset of substrate-independent inhibition following either continual (Fig. 4) or a short-term (Fig. 6) exposure, and the mild attenuation of inhibition in the presence of HSA, suggested rapid mechanism-based inhibition as being the primary mode of inhibition. Inhibition by BG, however, was delayed in onset (relative to DHB) and showed substrate dependence. With short exposure times, especially in the presence of HSA, the mild or lack of inhibition of midazolam 1′-hydroxylation by BG could have reflected enzyme activation in the face of inhibition (Figs. 4B and 6B). If the presence of basolateral HSA is assumed to represent the more physiologically relevant situation and that 30 min mimics a time the enterocytes are exposed to the furanocoumarins after consumption of a glass of grapefruit juice, these findings suggest that BG would have minimal interaction potential with a rapidly absorbed substrate belonging to the midazolam subgroup (midazolam is generally absorbed within 1 h) (Dundee et al., 1984). Moreover, if activation of CYP3A4 by BG (Fig. 2D) occurs in vivo, the current findings also suggest that coingestion of the substrate with BG might result in an interaction opposite to that expected from grapefruit juice (i.e., a decrease in substrate area under the curve). However, if the substrate belongs to the testosterone subgroup, ingestion with BG might result in a grapefruit juice-type interaction, even if the substrate were rapidly absorbed. The delayed inhibitory effect by BG may also explain why a single glass of lime juice [recently reported to contain a high concentration of BG (100 μM) but undetectable DHB (Bailey et al., 2003)] had minimal effect on the area under the curve of felodipine (Bailey et al., 2003), which belongs to neither the midazolam nor testosterone subgroup (Galetin et al., 2003).
With continual exposure of Caco-2 cells to BG (Fig. 4), the extent of inhibition of CYP3A4 activity appeared to “catch up” with DHB by 3 h, with a loss in the substrate-dependent nature of inhibition. This was consistent with the time-dependent rise in the cellular content of BG (Fig. 1, C and D); however, a time-dependent loss of substrate-dependent inhibition also occurred after a short-term exposure (Fig. 6), when cellular BG levels should have been decreasing. This, along with the BG-mediated decline in CYP3A4 protein, suggested gradual mechanism-based inhibition of the enzyme by BG. Hence, despite that BG is slow to act, a short exposure appears sufficient to initiate the events necessary for substantial inhibition. It is also reasonable to speculate that the concentration of BG at the enzyme gradually rises despite a decrease in cellular content, due to slow diffusion through the cell. Regardless of the mechanism, the current results would predict a greater magnitude of interaction with a substrate that is ingested several hours after consumption of food containing BG but not DHB (e.g., lime juice). Although lime juice is not routinely ingested, per se, it is used to prepare a number of popular beverages (e.g., margaritas), as well as marinades and salad dressings. Whether these lime juice-derived foods contain sufficient BG to cause an interaction remains to be determined.
As with CYP3A4 activity, the time-dependent decrease in CYP3A4 protein content by BG appeared delayed, but had caught up with DHB over time (Figs. 5 and 7). This was surprising since the onset of mechanism-based inhibition (of activity) was considerably delayed with BG relative to DHB. Because protein loss is believed to reflect intracellular degradation after mechanism-based inactivation (Lown et al., 1997), these findings suggest that the half-life of BG-inactivated CYP3A4 may be shorter than that of DHB-inactivated CYP3A4.
In summary, a systematic comparison of BG and DHB using a physiologically relevant intestinal cell system demonstrated a marked difference in both the rate of cell entry and time to onset of CYP3A4 inhibition, consistent with the 1000-fold difference in lipid solubility. DHB rapidly diffused across the apical membrane within minutes and effectively inhibited enzyme activity in a substrate-independent manner, which likely involved rapid mechanism-based inactivation. In contrast, the more lipophilic BG had a much slower rate of entry and a delayed onset of inhibition. With grapefruit juice, the current work suggests that CYP3A4 is maximally inhibited by DHB before BG has the opportunity to cause an effect. With foods containing BG only, minimal interaction should occur with rapidly absorbed drugs belonging to the midazolam subgroup. However, an interaction is predicted when the drug, whether it belongs to the midazolam or testosterone subgroup, is ingested several hours after consumption of the BG-containing food.
Footnotes
-
This work was supported by the National Institutes of Health Grants GM38149 and RR00046.
-
doi:10.1124/jpet.104.076836.
-
ABBREVIATIONS: BG, bergamottin; DHB, 6′,7′-dihydroxybergamottin; 1α,25-(OH)2-D3, 1α,25-dihydroxyvitamin D3; 6β-OH TST, 6β-hydroxytestosterone; 1′-OH MDZ, 1′-hydroxymidazolam; TEMED, N,N,N',N'-tetramethylethylenediamine; HPLC, high-performance liquid chromatography; Papp, apparent permeability coefficient(s); A→ B, apical-to-basolateral; B→ A, basolateral-to-apical; HSA, human serum albumin; DMEM, Dulbecco's modified Eagle's medium; IOD, integrated optical density.
- Received August 28, 2004.
- Accepted October 13, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}