Article Text

Abstract
Background—ArylamineN-acetyltransferases in humans (NAT1 and NAT2) catalyse the acetylation of arylamines including food derived heterocyclic arylamine carcinogens. Other substrates include the sulphonamide 5-aminosalicylic acid (5-ASA), which is an NAT1 specific substrate; N-acetylation of 5-ASA is a major route of metabolism. NAT1 and NAT2 are both polymorphic.
Aims—To investigate NAT expression in apparently healthy human intestines in order to understand the possible role of NAT in colorectal cancer and in the therapeutic response to 5-ASA.
Methods—The intestines of four organ donors were divided into eight sections. DNA was prepared for genotypingNAT1 and NAT2 and enzymic activities of NAT1 and NAT2 were determined in cytosols prepared from each section. Tissue was fixed for immunohistochemistry with specific NAT antibodies. Western blotting was carried out on all samples of cytosol and on homogenates of separated muscle and villi after microdissection.
Results—NAT1 activity of all cytosols was greater than NAT2 activity. NAT1 and NAT2 activities correlated with the genotypes of NAT1 and NAT2 and with the levels of NAT1 staining determined by western blotting. The ratio of NAT1:NAT2 activities showed interindividual variations from 2 to 70. NAT1 antigenic activity was greater in villi than in muscle. NAT1 was detected along the length of the villi in the small intestine. In colon samples there was less NAT1 at the base of the crypts with intense staining at the tips.
Conclusions—The interindividual variation in NAT1 and NAT2 in the colon could affect how individuals respond to exposure to specific NAT substrates including carcinogens and 5-ASA.
- arylamine N-acetyltransferase
- 5-aminosalicylate
- colorectal cancer
- drug metabolism
- inflammatory bowel disease
- diet
Statistics from Altmetric.com
- arylamine N-acetyltransferase
- 5-aminosalicylate
- colorectal cancer
- drug metabolism
- inflammatory bowel disease
- diet
Arylamine N-acetyltransferases metabolise a range of hydrazines and arylamines (see Weber and Hein1 for review). The arylamine substrates include environmental carcinogens found in the rubber and dye industry (see Sim et al 2 for review) and cigarette smoke.3 ,4 More importantly for intestinal exposure to carcinogens, heterocyclic amines and their hydroxylamine metabolites, which are substrates for NAT,5 are generated during the cooking of “well done” meat.6-8 NAT also metabolises arylamine drugs including 5-aminosalicylic acid (5-ASA).1 ,9
There are two NAT isozymes in humans, NAT1 and NAT2, both of which are now known to be polymorphic.10 There has been a continued discussion on the role of NAT polymorphism in determining susceptibility to colorectal cancer. Most of the investigations of NAT as a susceptibility factor in colorectal cancer have involved NAT2 polymorphism11 as it has been better characterised than NAT1 polymorphism. The situation with colorectal cancer is not as clear cut as in bladder cancer where slow acetylators at the NAT2locus are undoubtedly at increased risk.12 ,13 In colorectal cancer the role of NAT2 polymorphism has been controversial. NAT2 fast acetylators have been suggested to be at increased risk of colorectal cancer,5 ,14 but others15disagree with these findings. It has been suggested that fastNAT2 alleles are of increased importance as susceptibility factors in association with a particular NAT1 allele—theNAT1*10 allele14—as detected by a mutation in the polyadenylation site but the association of the mutation with colorectal cancer has been disputed.16 Recently it has been shown that alleles genotyped as NAT1*10 by detection of the mutation in the polyadenylation site14 ,17 are likely to include examples of individuals who also have a G to A mutation converting amino acid 187 from an arginine to a glutamine.18 ,19 This mutation affects the activity of NAT1 such that the NAT1*10 variant has a reduced level of activity. The newly identified NAT1*10 variant has been named NAT1*14 18 ,20 and a method for genotyping NAT1 which allows detection ofNAT1*14 has been described.18
Both NAT1 and NAT2 activities have been described in intestine9 ,21 and these enzymes will catalyse theN-acetylation of arylamine drugs, such as 5-ASA; arylhydoxylamines are generated by cytochrome P450 catalysed oxidation of arylamines in cooked meat.7 The role of NAT in carcinogenesis is thought to be important in catalysing the formation of the “ultimate carcinogen”, an N-acetoxyester which reacts with DNA.22 It has been suggested using expressed human NAT1 and NAT2 that the human NAT2 enzyme is more active with heterocyclic amines as substrates than is NAT1.5 It has also been shown, using heterologously expressed human NAT1 and NAT2, that the enzymes can catalyse both activation (O-acetylation) and deactivation (N-acetylation) reactions of heterocyclic amines and amines with a carbon only aromatic ring.23 The reaction rate depends on the isozyme, the reaction, and the substrate. Therefore interindividual variation in intestinal activities of NAT1 and NAT2 is likely to affect the range of carcinogenic compounds generated in situ.
NAT genes are encoded on 8p2224—a region which is frequently deleted in many tumours including colorectal tumours.25
The NAT1 isozyme is likely to be more important in 5-ASA acetylation than NAT2 from comparison of the activities of the expressed proteins. The NAT1 isoenzyme has been shown to be 19 000-fold more active with 5-ASA than the NAT2 isoenzyme.23
In view of the potential role of NAT in intestinal cancer and in metabolism of 5-ASA, which is used for local application in inflammatory bowel disease, it is important to establish the pattern of expression in normal intestine. We have therefore used a combination of enzymic and immunological methods to investigate NAT expression in human intestine obtained from organ donors.
Methods
TISSUE SAMPLES
Human intestines and livers were removed from adult organ donors who were victims of vehicular or cerebrovascular accidents. Briefly, the intestine was rinsed with ice cold 0.9% NaCl to remove particulate debris. The liver was perfused with 0.9% NaCl. Intestinal tissue was sectioned into 30 cm long portions and four equally spaced sections from the duodenum to the ileocaecal junction were numbered as sections 1–4. Equally spaced sections from the caecum to the rectum were numbered 5–8. The portions were placed in numbered polythene bags and the tissue was frozen in liquid nitrogen and then stored at −70°C. Intestinal tissue was defrosted on an ice cold glass plate. The mucosa was scraped from the outer muscle layer using a metal spatula and transferred to four volumes of ice cold homogenisation buffer, 250 mM sucrose, 100 mM phosphate (pH 7.4), 1 mM EDTA, and 1 mM DTT, with the proteinase inhibitor leupeptin (1 μg/ml). Liver was defrosted at room temperature and sliced into small pieces prior to homogenisation as above. Cytosols were prepared by centrifugation. The cytosolic fraction was the supernatant after centrifugation of the homogenate (10 000g for 20 minutes, followed by 100 000 g for 60 minutes at 4°C). Aliquots (1 ml) of cytosols were made and stored in liquid nitrogen apart from transportation on dry ice from Seattle to Oxford. Samples remained frozen and were returned to liquid nirogen immediately on arrival.
ENZYMIC ASSAYS
The N-acetylation of 100 μMp-aminobenzoic acid and 1000 μM sulphamethazine were used as probes for human NAT1 and NAT2 activities respectively. Cytosols (40 μg protein) were incubated for five or 10 minutes at 37°C with either p-aminobenzoic acid or sulphamethazine in the presence of 100 μM acetyl CoA and an acetyl carnitine based recycling system26 in a total volume of 90 μl. Reactions were stopped by the addition of 10 μl of 15% perchloric acid. Acetylated metabolites in the supernatant after centrifugation (20 000g for five minutes at 4°C) were measured by high pressure liquid chromatography with ultraviolet detection (266 nm), as previously described.27
ANTIBODIES
Antibodies recognising NAT1 and NAT2 (called 177), specific for NAT1 (called 184), or specific for NAT2 (called 4768) were prepared as described previously.28 Western blots were carried out as described previously using a dilution of the first antibody of 1/500 to 1/12 000. The second antibodies (goat antirabbit immunoglobulin) were coupled with horseradish peroxidase and were used at a dilution of 1/10 000. Slides bearing 10 μm thick sections of formalin fixed, paraffin wax embedded human small intestine and colon were subjected to immunohistochemical staining using the Vecastain Elite ABC peroxidase (Universal) kit (Vector Labs, Peterborough, UK). Endogenous peroxidase activity was inhibited using 1% (vol/vol) H2O2in 15 mM Tris-HCl, pH 7.5 containing 140 mM NaCl (TBS); sections were incubated with rabbit antiserum or preimmune serum at 4°C overnight and three 10 minute washes were performed between each antibody incubation. Rabbit antisera were diluted in TBS containing 1% normal horse serum. Following immunostaining, sections were counterstained with Gill’s haematoxylin and dehydrated. Coverslips were attached with DPX mountant.
GENOTYPING
DNA was prepared from homogenised pieces of intestine by extraction with phenol and chloroform.13 NAT2genotype was determined by the method of Risch et al 13 and NAT1 genotype was determined by a modification of the method described by Payton and Sim.18Briefly, NAT1 alleles were initially identified by MboII restriction fragment length polymorphism of polymerase chain reaction (PCR) products.14 The presence of a mutation (G in wild type for A in mutant alleles) at nucleotide 560 in the coding region of NAT1 was detected by Bsa0I restriction fragment length polymorphism of the PCR products obtained using primer N539 as the sense primer with primer N714 as the antisense primer (table 1). Allele specific PCR using primer N539 as sense primer with N1110b as antisense primer was used to confirm the mutant allele as NAT1*14. Amplication conditions and restriction enzyme digests were as defined previously.18
Primers used for detecting NAT1*14 allele
Results
GENOTYPING AND INTESTINAL ACTIVITY OFN-ACETYLTRANSFERASES
Table 2 illustrates the genotype of NAT1 and NAT2 in the four intestinal samples. The activities of NAT1 and NAT2 were determined using p-aminobenzoic acid and sulphamethazine, respectively, as substrates. The relation between the phenotype of NAT2 determined from the genotype and the measured activities of samples with sulphamethazine as substrate is as would be expected. Two of the samples were determined as homozygous for slow acetylator alleles and these samples had the lowest activities with sulphamethazine. The sample which was determined as being homozygous fast for NAT2 had the highest NAT2 activity and the individual who was determined as having one fast and one slow NAT2 allele had an intermediate level of activity. The correlation of NAT2 activity in intestinal samples towards sulphamethazine with NAT2 type has been commented on by Ilettet al.21 As in the present study, Ilettet al found the level of NAT1 activity to be greater than the level of NAT2 activity.
Genotypes of NAT in intestinal samples
The NAT1 activities of three of the samples in the present study determined with p-aminobenzoic acid as substrate were similar and these samples were all genotyped as being homozygous for the wild type NAT1*4 allele. One individual had a lower level of NAT1 activity and was genotyped as 1*4, 1*14 (fig 1). It has previously been shown that a mutation at nucleotide 560, as is found in allele NAT1*14, which results in the substitution of an arginine residue in the wild type NAT1 at amino acid residue 187 for a glutamine in the mutant, is associated with a low level of NAT1 activity.18 ,20 The identification of this mutation in an individual with a low level of NAT1 activity in intestine further confirms the relation between this allele and reduced NAT1 activity.

Identification of NAT1 alleles by PCR and restriction enzyme digestion with Bsa0I. Template DNA was amplified with the primer pair N539 (sense) and N714 (antisense) and then digested with Bsa0I. The wild type allele (G at position 560 of coding sequence) NAT1*4 is digested to a 155 kb fragment. The mutant allele (NAT1*14) is not digested by Bsa0I and the band is 175kb. The products were separated on a 4% Metaphor agarose gel. (a) genomic DNA as template. (b) Tracks 1 and 2 show genomic DNA of individual 22/3 as template. Tracks 3 and 4: the template was the product of a “nested PCR” using initially allele specific amplification (N539 and N1110b as primers) with genomic DNA from individual 18/5. The primers are described in table 1.
ACTIVITY ALONG THE LENGTH OF HUMAN INTESTINE
The distribution of NAT1 and NAT2 along the length of the intestine was measured in two ways. It was measured by determining the activity of NAT1 with p-aminobenzoic acid and the activity of NAT2 with sulphamethazine. NAT1 activity was found in all sections of intestine. In three of the individuals the activity was measured maximally at the region between the small and large intestine (fig 2a). However with sample 18/5, showing the lowest level of NAT1 activity, there was an increase in activity towards the distal end of the intestine (fig 2a). The distribution of activity in samples along the length of the intestine was matched by the NAT1 antigenic activity detected by western blotting of the same amounts of protein from each section of intestine (fig 3). The antiserum identifies a band at around 33 000 molecular weight corresponding to the expected molecular weight of NAT1.10

Detection of NAT enzymic activity in different sections along the length of human intestine. (a) Enzymic acivity of NAT1 was determined in cytosols with p-aminobenzoic acid; (b) NAT2 activity was determined with sulphamethazine. (c) Ratios of activities of NAT1 to NAT2. Samples 1–8 correspond to sections along the length of the intestine from the duodenum (1) to the rectum (8).

Expression of NAT1 in human intestine detected by western blotting. (a) Samples of mucosal cytosol along the length of human intestine from individual 22/3 were prepared and analysed by polyacrylamide gel electrophoresis and western blotting (tracks 2–9). The sample in track 2 is from the most proximal section (corresponding to 1 in fig 2) and the sample in track 9 is from the rectal end. Track 1 contained the molecular weight markers which are not detected by the antiserum. (b) Samples (52 mg total protein) from the cytosols with maximum activity from individual 22/3 (tracks 1 and 2) and from individual 18/5 (tracks 3 and 4) were analysed. The first antibody was antiserum 184 used at a dilution of 1 in 1000 and the second antibody was goat antirabbit immunoglobulin at a dilution of 1 in 10 000. The molecular weights were determined from prestained markers run on the same gel.
The distribution of NAT2 along the length of the intestine showed a decrease from the proximal to the more distal sections of around twofold in the two fast acetylator individuals (fig 2b). If the ratios of NAT1 activity to NAT2 activities are compared along the length of the intestine it is clear that there is considerable interindividual variation within this small group of individuals (fig 2c). Individuals 24/3 and 22/3 are both slow acetylators for NAT2 and have two wild type alleles for NAT1. The NAT1 activities of both of these individuals compared with NAT2 activities show much higher relative NAT1 activities in intestine. This is particularly notable for individual 24/3 who has 50 to 70 times more NAT1 activity than NAT2 activity in the lower sections of intestine. The ratio of NAT1 activity to NAT2 activity is more similar in the two individuals who are homozygous (11/5) or heterozygous (18/5) fast acetylators at the NAT2 locus. The intestine of individual 18/5, who carries the mutantNAT1*14 allele, has a maximum of 10 times more NAT1 than NAT2 activity, but again the biggest difference between the two enzymic activities is in the colon rather than the small intestine. The level of expression of NAT activity in the different individuals is reflected in the antigenic activities detected in samples from the same region of the gut containing the same amount of total protein (fig 3). In both individuals a band appears at around 33 000 molecular weight (the expected molecular weight of NAT1), indicating that the lower activity of 18/5 is not due to proteolytic degradation of NAT1 protein. It seems that in the individual with the NAT1*14 allele there is less protein detectable by the NAT1 specific antiserum, 184. It is unlikely that this is due to a difference in the recognition of the mutant NAT1 by this antibody as the antibody recognises the C-terminal peptide of NAT1 which is the same in the isoenzymes encoded by both theNAT1*4 and NAT1*14 alleles. It would seem that the amount of NAT protein in the individual with the NAT1*14allele is less. Site directed mutagenesis studies are required to confirm whether the protein product of the mutant allele with arginine 187 mutated to glutamine is active or not. cellular distribution of nat Antiserum 177 which recognises NAT1 and NAT2 and antiserum 184 raised against a C-terminal peptide of NAT1 have been shown previously to be specific.28 Paraffin wax sections from the small intestine and colon of all four individuals were stained with either 177 (fig 4) or 184 (fig 5). The pattern of staining in all four individuals was very similar to the representative samples shown in fig 4. In the small intestine, staining was observed in the epithelial cells along the whole length of the villus, from the tip to the crypt of Lieberkuhn (fig 4a, fig 5a). The staining was cytoplasmic and was seen in all epithelial cell types. In goblet cells the antibody seems to be excluded from the mucous inclusions which are visible as transparent areas within the stained cytoplasm. In the colon, however, staining was restricted to the epithelial cells at the luminal surface, the crypts being devoid of staining (fig 4b). The most intense staining was found at the luminal tip of the columnar epithelial cells. As in the small intestine, the antibody seemed to be excluded from the mucous inclusions of goblet cells. No staining was detected in either the small intestine or the colon when preimmune rabbit serum was used instead of antiserum 177 or184.

Expression of NAT in human intestine detected by immunohistochemistry with antiserum 177. Sections of duodenum (a,c) and colon (b,d) were stained with antiserum 177 (diluted 1 in 3000) (a,b) or the corresponding dilutions of preimmune serum (c,d). Intestine from individuals 18/5 (a,c) and 24/3 (b,d) are shown. Original magnification: (c) ×100; (a,b,d) ×200.
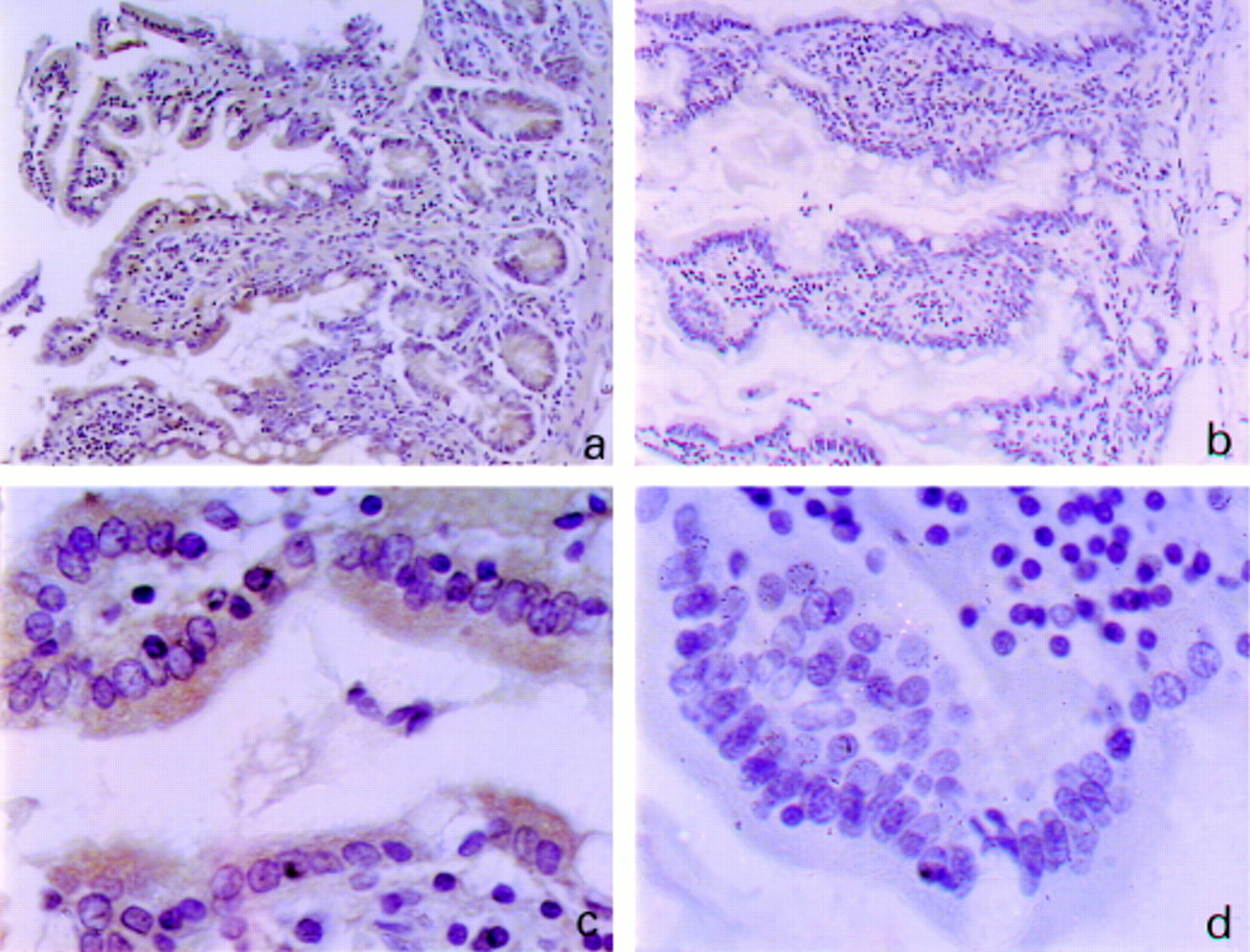
Expression of NAT1 in human intestine detected by immunohistochemistry with antiserum 184. Small intestine from individual 22/3 was stained with antiserum 184 (diluted 1/800) (a,c) or with the corresponding dilution of preimmune serum (b,d). Original magnification: (a,b) ×200; (c,d) ×1000.
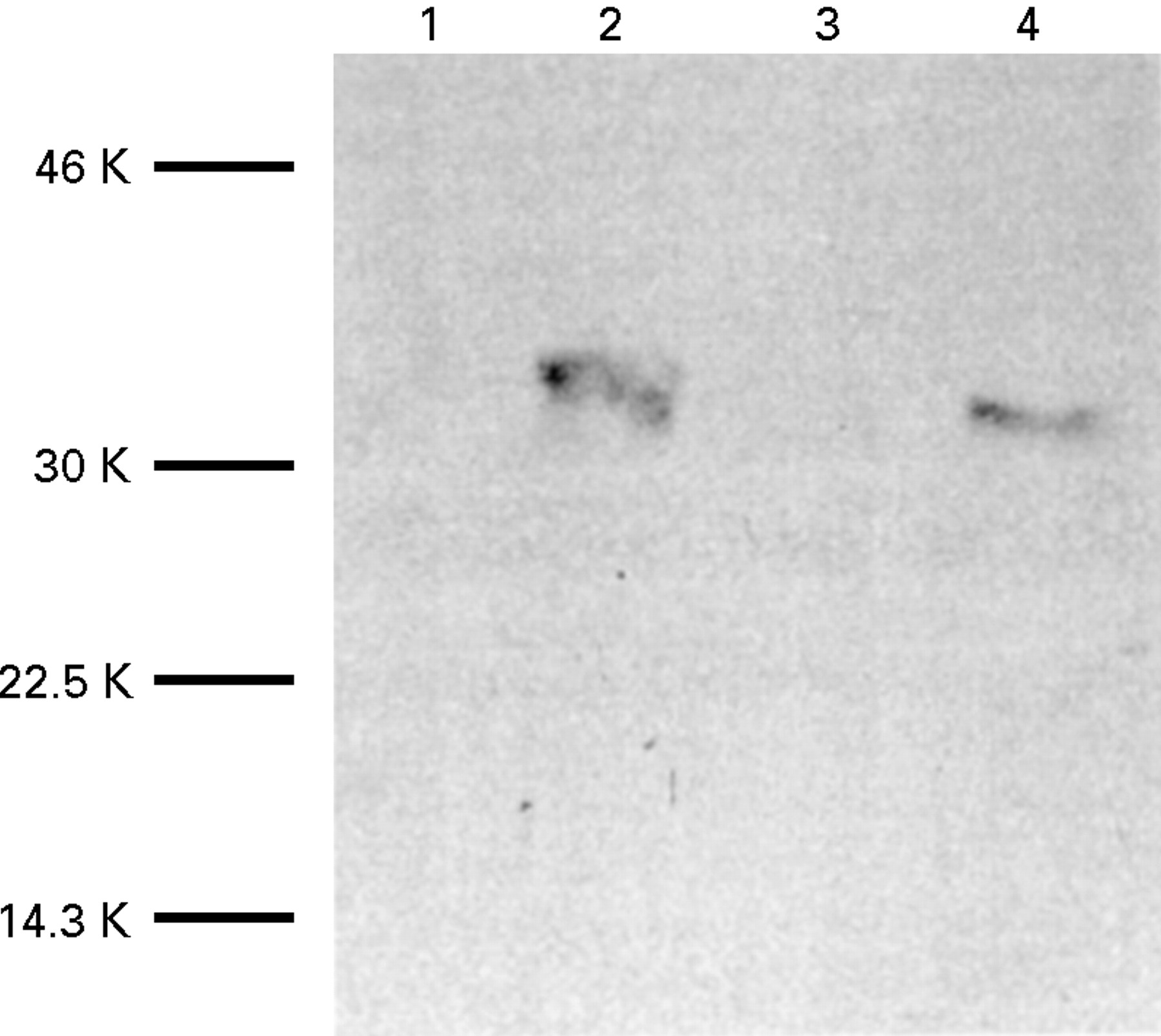
The existence of NAT in villi has also been identified by microdissection of villi from the underlying muscle. A western blot showed no band for the same amount of muscle protein whereas the villi showed a band corresponding to NAT1 (fig 6). These results agree with a previous preliminary report29 which illustrated that the NAT1 activity in cytosol from mucosa of small intestine was six times greater than the activity associated with a muscle cytosol, while no NAT2 activity could be detected in the muscle cytosol.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Determination of NAT expression in villi and muscle by western blotting. Villi were microdissected from muscle samples of small intestinal samples (corresponding to track 4 of fig 3a) and large intestine (corresponding to track 7 of fig 3a). The mucosa or muscle mixtures (40 μg total protein of each) were analysed by gel electrophoresis and western blotting using antibody 184 as above. Tracks 2 and 4 show villi and tracks 1 and 3 show muscle. Tracks 1 and 2 are small intestine and tracks 3 and 4 are large intestine.
Discussion
The detection of NAT activity in intestine was established by the early work of Jenne9 who showed using isoniazid that intestinal mucosa and liver had equivalent specific activities of NAT. Isoniazid is now known to be an NAT2 substrate.30 However, it seems that NAT activity in human intestine is mainly due to NAT1. At the time when Jenne carried out his studies, although he usedp-aminosalicylate, now known to be a NAT1 substrate,23 he did not report on activities of individual intestinal homogenates with both isoniazid andp-aminosalicylate. It is not possible to compare his data with the present comparison of NAT1 and NAT2 activities in human intestine. Jenne suggested that the activities of NAT2 in liver and intestine were very similar. In a preliminary report31 for three individuals where NAT2 in liver and intestine were measured it was found that NAT2 specific activities in liver were either two- to threefold higher than in intestine in fast NAT2 acetylators or of a comparable value in slow NAT2 acetylators. Comparison of activities of NAT1 and NAT2 in intestines of the same individual has shown in all samples that there is a much higher specific activity of NAT1 than NAT2. This is found in both fast and slow acetylators although the difference between the two activities is much more notable in slow acetylators at the NAT2 locus. These results are fully in agreement with the studies of Ilett et al.21 The present results also show that the genotype of NAT1 affects the activities of NAT1 detected in intestine. The detection of NAT1 along the length of intestine suggests that as far as the colon is concerned the activity of NAT1 is likely to be extremely important in generating compounds from aromatic arylamines or hydroxylamines which are likely to have a role in colon carcinogenesis.6-8 The interindividual variation even in the four samples which were available indicates that genotyping for NAT1 in colorectal cancer has a sound biochemical basis. The lack of a clear correlation of NAT2 type with colorectal cancer susceptibility15 may be due to the low level of NAT2 which is present in colon in comparison with NAT1.
Animal studies have also shown that NAT is present in intestine in hamster,32 mouse,32-34 where it has been localised to the villi of the small intestine,33 and rat.32 In these rodents there is evidence for a difference in expression of individual NAT isozymes along the length of the intestine. Therefore the understanding of the expression of human NAT isoenzymes along the length of the intestine together with comparison of the substrate specificities of the NAT isoenzymes of different species will allow the establishment of sensible models for understanding human colon carcinogenesis35 and may serve as an additional means of validating animal models of inflammatory bowel disease.36
The identification of NAT in epithelial cells may be important because susceptibility factors in tumour development include pre-existing adenomas.37 Ulcerative colitis or Crohn’s disease are also associated with epithelial dysplasia and have been considered to be precursor lesions, rather than predisposing factors for colon carcinoma. In the inflammatory bowel disorders, the methods are available to determine whether NAT1 expression is altered. NAT1 polymorphism may also have a role in the therapeutic response or adverse effects to 5-ASA38 and the epithelial localisation of NAT1 which is shown here to be the most active NAT in colon should be borne in mind when designing other drugs active in inflammatory bowel disease.
Acknowledgments
We are grateful to Aileen Watson and Susan Rolls for excellent technical assistance, to Gamini Chandrasena for advice and help in processing human intestine, to Mike Ball for expert assistance with histology and photomicrography, and to Dr Bryan Warren for helpful discussions. The Oxford work was supported by the Association for International Cancer Research and the Wellcome Trust. The work was also funded in part by grants NIHA127664 and NIHDK41978 (JDU).