


Abstract
CYP2C9 is a polymorphic gene for which there are four known allelic variants; CYP2C9*1,CYP2C9*2, CYP2C9*3, andCYP2C9*4. In the present study, DNA from 140 European Americans and 120 African Americans was examined by single-strand conformational polymorphism and restriction fragment length polymorphism analyses, resulting in the identification of a new CYP2C9 variant, CYP2C9*5. This variant is derived from a C1080G transversion in exon 7 of CYP2C9 that leads to an Asp360Glu substitution in the encoded protein. TheCYP2C9*5 variant was found to be expressed only in African Americans, such that approximately 3% of this population carries the CYP2C9*5 allele. The variant was expressed in, and purified from, insect cells infected with a recombinant baculovirus. Comparative kinetic studies using the purified wild-type protein CYP2C9*1; the Ile359Leu variant, CYP2C9*3; and the Asp360Glu variant, CYP2C9*5 were carried out using (S)-warfarin, diclofenac, and lauric acid as substrates. The major effect of the Asp360Glu mutation was to increase the K mvalue relative to that of CYP2C9*1 for all three substrates: 12-fold higher for (S)-warfarin 7-hydroxylation, 5-fold higher for the 4′-hydroxylation of diclofenac, and 3-fold higher for the ω-1 hydroxylation of lauric acid. V max values differed less than K m values between the CYP2C9*1 and CYP2C9*5 proteins. In vitro intrinsic clearances for CYP2C9*5, calculated as the ratio ofV max/K m, ranged from 8 to 18% of CYP2C9*1 values. The corresponding ratio for CYP2C9*3 was 4 to 13%. Accordingly, the in vitro data suggest that carriers of the CYP2C9*5 allele would eliminate CYP2C9 substrates at slower rates relative to persons expressing the wild-type protein.
Genetically determined differences in drug-metabolizing enzyme capacity can be an important factor in determining interindividual variability in drug response (Evans and Relling, 1999). The clinical significance of single-nucleotide polymorphisms (SNPs) in the NAT2,CYP2D6, and CYP2C19 genes that underlie interindividual and ethnic differences in acetylation and cytochrome P450 (P450)-mediated “poor metabolizer” phenotypes, respectively (Wood, 1998), has been well established over the last decade. SNPs in the CYP2C9 gene have also increasingly been recognized to have clinically significant consequences.
CYP2C9, the major CYP2C isoform found in the human liver (Goldstein and de Morais, 1994; Richardson et al., 1997), participates in the oxidation of several endogenous compounds such as progesterone and arachidonic acid (Rifkind et al., 1995; Yamazaki et al., 1998); its most prominent function, however, is the oxidative metabolism of drugs and other xenobiotics. CYP2C9 hydroxylates a wide array of generally acidic substrates in diverse therapeutic categories, including anticonvulsants, diuretics, and nonsteroidal anti-inflammatories (Miners and Birkett, 1998). Of special interest is the CYP2C9-mediated metabolism of drugs with a narrow therapeutic index, such as (S)-warfarin and phenytoin, because a marked impairment in CYP2C9 metabolic activity can lead to severe toxicity (Steward et al., 1997; Ninomiya et al., 2000).
In addition to the wild-type protein, CYP2C9*12 (Arg144, Ile359), SNPs within the coding region of the gene produce three variant allozymes, CYP2C9*2 (Cys144, Ile359), CYP2C9*3 (Arg144, Leu 359), and CYP2C9*4 (Arg144, Thr359), each with reduced intrinsic metabolic activity (Rettie et al., 1994; Haining et al., 1996;Sullivan-Klose et al., 1996; Crespi and Miller, 1997; Ieiri et al., 2000). Frequencies of the CYP2C9*2 and CYP2C9*3alleles in white populations are in the range of 0.06 to 0.13 (Stubbins et al., 1996; Sullivan-Klose et al., 1996; Yasar et al., 1999), but are an order of magnitude lower in African Americans in the only study reported to date (Sullivan-Klose et al., 1996).
The CYP2D6*17 allele is selectively expressed in African and African-American populations (Masimirembwa et al., 1996; Bradford et al., 1998; Leathart et al., 1998), and so it seemed possible that population differences in polymorphisms might also exist for theCYP2C9 gene. The present study identifies a newCYP2C9 allele, CYP2C9*5 (Arg144, Ile359,Glu360), that is expressed in African Americans but not in European Americans. The functional consequences of the Asp360Glu mutation were evaluated by expression of CYP2C9*5 in insect cells and determination of the kinetic parameters,V max and K m, for (S)-warfarin, diclofenac, and lauric acid metabolism. The likely in vivo consequences for expression of this new variant of CYP2C9 are discussed.
Materials and Methods
Chemicals and Reagents.
Diclofenac, lauric acid, 7-ethoxycoumarin, 15-hydroxypentadecanoic acid, penicillin-G, streptomycin sulfate, amphotericin-B, and dNTPs were obtained from Sigma-Aldrich Co. (St. Louis, MO). (S)-Warfarin, hydroxylated warfarin metabolites, and the hydroxylated lauric acid metabolites were synthesized as described previously (Haining et al., 1996; Guan et al., 1998). 3′-Hydroxy, 4′-hydroxy, and 5-hydroxy-diclofenac were provided by Novartis (Basel, Switzerland). Transformer site-directed mutagenesis kits were purchased from CLONTECH (Palo Alto, CA), restriction enzymes from New England BioLabs (Beverly, MA), and AmpliTaq from Applied Biosystems (Foster City, CA). Excell-405 expression culture medium was obtained from JRH Biosciences (Lenexa, KS), and heat-inactivated fetal bovine serum from Invitrogen (Carlsbad, CA). The pFastBac vector and the Bac-to-Bac expression system were also purchased from Invitrogen.
Subjects.
Two hundred and sixty genomic DNA samples (140 from European Americans and 120 from African Americans) were obtained from peripheral leukocytes. All subjects were unrelated adults residing in mid-Tennessee, and race was defined by self-reporting (Senior and Bhopal, 1994). The protocol was approved by the Vanderbilt University Institutional Review Board-Health Services and written informed consent was obtained.
PCR-RFLP Analysis.
Detection of the CYP2C9*2 andCYP2C9*3 alleles were performed using a polymerase chain reaction-based restriction fragment length polymorphism (PCR-RFLP) analysis. Briefly, PCR was carried out in a 50-μl final reaction volume including 2.5 mM MgCl2, 10 mM Tris-HCl, 50 mM KCl, 10 mM dNTPs, 1.5 U of AmpliTaq DNA polymerase and approximately 200 ng of human genomic DNA, and the primer (0.32 μM) pairs 5′-TACAAATACAATGAAAATATCATG and 5′-CTAACAACCAGACTCATAATG-3′ forCYP2C9*2 and 5′-AATAATAATATGCACGAGGTCCAGAGATGC- 3′ and 5′- GATACTATGAATTTGGGACTTC- 3′ for CYP2C9*3. Aliquots of each PCR product (15 μl) were subjected to restriction enzyme analysis with 5 U of AvaII (CYP2C9*2) or NsiI (CYP2C9*3). After overnight incubation at 37°C, digested products were separated by electrophoresis using a 3% agarose gel.
Single-Strand Conformational Polymorphism Analysis.
A nonisotopic single-strand conformational polymorphism (SSCP) analysis (Hongyo et al., 1993) was used for the identification of allelic variants. Briefly, PCR products encompassing the whole of CYP2C9 exon 7 were obtained using primers 5′-TACTTGTGTCTTATCAG-3′ and 5′-CATGGAGTTGCAGTGTAGGAG-3′. Subsequently, a mixture consisting of 10 μl of PCR product (20–200 ng of DNA), 3.6 μl of Ficoll loading buffer (0.25 μl bromphenol blue and 0.25% xylene cyanol [MW, 400,000; 15% (w/v)], and 11.4 μl of 1.5× TBE electrophoresis buffer; addition of distilled water to a final volume of 1000 ml) was prepared to yield a total volume of 25 μl (2.5-fold dilution of the original PCR product). This mixture was heated at 94°C for 4 min to ensure maximal denaturation and, then, immediately plunged into ice before loading the sample onto a precast 20% nondenaturating polyacrylamide TBE minigel (Novex, San Diego, CA). A thermostatically controlled refrigerated circulator and heat-exchange system (Thermoflow; Novex) were used to maintain a constant (± 0.5°C) gel temperature. The best single-strand DNA separation was achieved using a preset temperature of 7.5°C with the gel run at 300 V (30.3 V/cm) for 5 h. Subsequently, the gels were stained with a 0.5 μg/ml solution of ethidium bromide in 1.5× TBE buffer for 20 min. The bands were visualized using a UV viewing box and photographed. Variations in the single-strand mobility patterns were clearly visualized and permitted abnormal bands to be easily cut out from the gel. Reamplification and sequencing was carried out by soaking the band of interest in 50 μl of distilled water overnight. Subsequently, 5 μl of this solution was used as DNA templates for PCR in the same manner outlined previously. The PCR products were purified using an anion exchange column (QIAquick; QIAGEN, Chatsworth, CA), then sequenced (ABI 3700; PerkinElmer Life Sciences, Boston, MA).
Mutagenesis and Vector Construction.
Site-directed mutagenesis of the CYP2C9 gene in pUC19 (Haining et al., 1996) was achieved with the Transformer site-directed mutagenesis kit (CLONTECH, Palo Alto, CA). The sequence of the selection oligonucleotide chosen for mutagenesis was that provided in the Transformer protocol, which changes a unique Aat II restriction endonuclease site into an EcoRV site. The mutagenic oligonucleotide consisted of the sequence 5′-CAGAGATACATTGAGCTTCTCCCCACCAGCCTG-3′. The mutation was confirmed using automated sequencing and the variant subcloned into pFastBac as described previously (Haining et al., 1999).
Baculovirus-Mediated Expression and Purification from Insect Cells.
Growth and passage of Trichoplusia ni cells was carried out at 27°C on 100-mm culture dishes using Excell-405 medium supplemented with 0.5% heat-inactivated fetal bovine serum, 100 μg/ml penicillin-G, 100 μg/ml streptomycin sulfate, and 0.25 μg/ml amphotericin-B. Viral construction for the wild-type and mutant genes was achieved using the Bac-to-Bac system. Expression and purification of the wild-type and mutant proteins were carried out as described previously (Haining et al., 1996), with the exception that a DEAE-Sepharose column chromatography was omitted from the purification steps. Only cultures exhibiting expression > 200 nM were used for enzyme purification. Specific contents of the final enzyme preparations exceeded 12 nmol of holo-P450/mg of protein.
Peptide Mapping by MALDI-TOF Mass Spectrometry.
Protein samples (500 pmol) were added to buffer (50 mM Tris-HCl, pH 7.6, 1 mM CaCl2) to a final concentration of 2.5 μM. Sequencing grade modified trypsin (Promega, Madison, WI) was added [1.5 μg, 5% (w/w)] and the samples incubated at 37°C for 30 min. The samples were then made 0.5% in trifluoroacetic acid and desalted using C18 ZipTips (Millipore, Bedford, MA) according to the manufacturer's instructions. Each sample (0.6 μl) was then spotted on top of 0.6 μl of an α-cyano-4-hydroxycinnamic acid solution on a MALDI target plate and allowed to dry. Mass spectra were obtained on a Bruker Biflex III MALDI-TOF mass spectrometer, operating in positive-ion reflectron mode and calibrated with external standards before each use. Each spectrum was the sum of at least 50 laser shots. Data were processed using the Bruker XMass system (Bruker, Newark, DE). Peptide molecular masses (monoisotopic MH+) were calculated using the program Sherpa (University of Washington, Seattle, WA).
General Enzyme Assay Conditions.
Unless otherwise noted, incubation mixtures contained 100 mM potassium phosphate, pH 7.4, 50 pmol of CYP2C9, 100 pmol P450 reductase, 50 pmol of cytochrome b5, 20 μg of dilaurylphosphatidylcholine, 1 mM NADPH, and the appropriate substrate in a final volume of 1.0 ml. HPLC analysis of (S)-warfarin and diclofenac metabolites was performed using a Hewlett-Packard model 1050 (Hewlett Packard, Palo Alto, CA), and GC/FID analysis of lauric acid metabolites with a Hewlett-Packard model 5890.
Metabolic Assays and Enzyme Kinetics.
Rates of formation of 4′-hydroxydiclofenac by reconstituted enzyme mixtures were determined by HPLC as described previously (Haining et al., 1999), except that enzymatic reactions were initiated with NADPH and allowed to proceed for 15 min. Diclofenac concentrations ranging from 1 to 100 μM were used for determination of kinetic parameters. Rates of formation of 11-hydroxylauric acid were quantified by gas chromatography, also as described previously (Haining et al., 1999), except that 15-hydroxypentadecanoic acid was used as the internal standard. Concentrations of lauric acid for kinetic studies ranged from 10 to 150 μM. (S)-Warfarin 7-hydroxylation was measured by HPLC using the fluorometric method of Lang and Bocker (1995). Briefly, metabolic reactions contained 100 pmol of P450, 200 pmol of reductase, 100 pmol of cytochrome b 5 in a final volume of 0.5 ml. After 30 min, the reaction was quenched with 10 μl of 70% perchloric acid and 10 ng of 7-ethoxycoumarin was added as an internal standard. Denatured protein was precipitated by centrifugation at 7000 rpm for 5 min and 100 μl was injected for HPLC analysis. The concentration range used for kinetic determinations was from 2 to 100 μM. To conserve P450 reductase and cytochrome b5 to ensure that all kinetic measurements were obtained with the same purified batches of these accessory proteins, incubations with each of the three substrates were conducted in duplicate. K m andV max values were estimated using the k.cat program (Biometallics, Inc.). K m andV max parameters for the CYP2C9*1-mediated metabolism of (S)-warfarin, diclofenac, and lauric acid were similar to those reported previously (Haining et al., 1999). Spectral P450 measurements were obtained using the method of Estabrook et al. (1972).
Results
Identification of a Novel CYP2C9 Variant in African Americans.
The occurrence of functionally defective CYP2C9*2 andCYP2C9*3 alleles in African Americans has been reported to be very low with allele frequencies in the range of 0.5 to 1% (Sullivan-Klose et al., 1996). Therefore, we sought to confirm the low frequency of CYP2C9*3 in African Americans and determine whether other polymorphisms of functional significance exist in this population. The CYP2C9*3 variant was readily detected usingNsiI-based RFLP analysis (Fig.1A) and by SSCP analysis (Fig. 1B). Eight percent of whites, but only 2% of African Americans, were carriers of the CYP2C9*3 allele (Table 1). None of the subjects were homozygous for the CYP2C9*3genotype.

A, PCR-based RLFP analysis of CYP2C9wild type (*1/*1; sample 22k) and heterozygous CYP2C9*3 variant (*3/*1; sample 8m) among African-American subjects. B, SSCP analysis band patterns for CYP2C9*1(22k) and *3 (8m)
CYP2C9 variant alleles (A) and genotypes (B) among healthy European Americans (n = 140) and African Americans (n = 120)
SSCP analysis revealed the presence of an additional variant in exon 7 of CYP2C9 associated with a C1080G transversion resulting in an Asp360Glu substitution (CYP2C9*5). Interestingly, this variant was observed only among African Americans (Fig.2). In fact, it was a dominant mutation found in this population because more than 3% of the African-American subjects were found to carry the CYP2C9*5 allele (Table 1). Not surprisingly, considering the relatively low allele frequency, no study subjects were homozygous for this newly identified variant. Direct sequencing of exon 7 of a restricted set of white (n = 50) and African-American (n = 75) DNA samples failed to reveal the presence of the CYP2C9*4(data not shown).

SSCP analysis and DNA sequencing: identification of a new CYP2C9 variant in a female African-American (sample 28d-I) subject versus wild type (22k) and CYP2C9*3(8m)
Heterologous Expression and Characterization of CYP2C9*5.
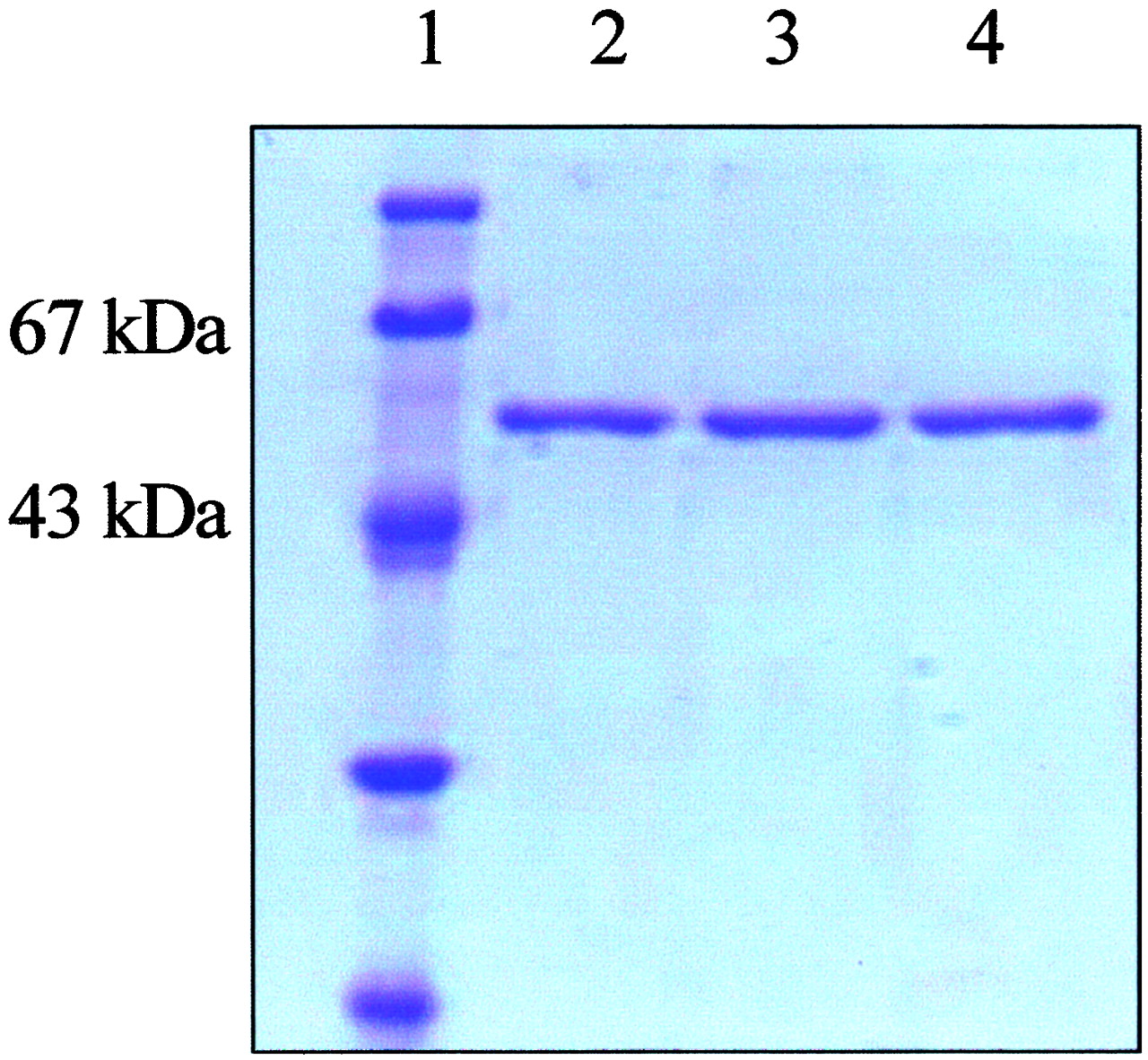
To gain insights into the metabolic capabilities of CYP2C9*5, the variant cDNA was created by site-directed mutagenesis and subcloned into pFastBac for production of a recombinant baculovirus that was used to infect insect cells. Separate cultures were infected with recombinant viruses constructed previously (Haining et al., 1996) coding for the CYP2C9*1 and CYP2C9*3 variants. After 48 to 72 h, concentrations of spectrally determined P450 exceeded 200 nM, at which point cells were harvested for purification by a combination of hydrophobic and hydroxyapatite chromatography. These procedures yielded highly purified preparations of each of the three variants (Fig.3).

SDS-polyacrylamide gel electrophoresis analysis of purified CYP2C9*1, CYP2C9*3, and CYP2C9*5. Proteins were expressed, purified, and quantified as described under Materials and Methods. CYP2C9 proteins were separated on a 9% acrylamide resolving gel and visualized by staining with Coomassie blue. Lane 1, molecular mass markers (Invitrogen). Lanes 2 to 4, 10 pmol of CYP2C9*1, CYP2C9*5, and CYP2C9*3, respectively.
To confirm the identity of purified CYP2C9*5, the enzyme preparation was digested with trypsin and a peptide map generated by MALDI-TOF mass spectrometry. Substitution of Asp at position 360 with Glu should result in an increase in mass of 14 Da for the 18-amino-acid tryptic fragment corresponding to CYP2C9 residues 358 to 375. Wild-type CYP2C9*1 exhibited a prominent isotope peak envelope corresponding to this fragment at m/z 1999, whereas, as expected, the corresponding peak in CYP2C9*5 was shifted tom/z 2013 (Fig. 4).

Confirmation of the Asp360Glu mutation in recombinant CYP2C9*5 by MALDI-TOF mass spectrometry of tryptic digest obtained as described under Materials and Methods. Numbers in parentheses are calculated monoisotopic masses. Top, partial MALDI spectrum depicting isotope clusters for the186RFDYKDQQFLNLMEK200 and358YIDLLPTSLPHAVTCDIK375 tryptic fragments atm/z 1975 andm/z 1999, respectively, from wild-type CYP2C9*1. Bottom, the corresponding portion of the MALDI spectrum for CYP2C9*5, demonstrating both conservation of the 186–200 peptide and an increase of 14 Da (to m/z 2013) for the 358–375 fragment that now contains the Asp360Glu mutation.
Relative Catalytic Efficiencies of CYP2C9*5, CYP2C9*3, and CYP2C9*1.
Upon reconstitution with P450 reductase, cytochromeb 5, and lipid, each of the CYP2C9 variants catalyzed the metabolism of (S)-warfarin, diclofenac, and lauric acid with widely different apparentK m and V maxvalues. Both CYP2C9*3 and CYP2C9*5 exhibited catalytic efficiencies for (S)-warfarin 7-hydroxylation, diclofenac 4′-hydroxylation, and lauric acid ω-1-hydroxylation that were approximately 10 ± 5% of those measured for the wild-type enzyme (Table2). Although in the case of CYP2C9*3 this lower activity was a consequence of increasedK m coupled with significant decreases inV max, for CYP2C9*5, the primary effect was on Km . Indeed, for the substrates (S)-warfarin and diclofenac,V max was largely unaffected by the Asp360Glu mutation, but K m increased 12-fold and 4.6-fold, respectively.
Mean kinetic parameters for the formation of (S)-7-hydroxy warfarin, 4′-hydroxy diclofenac, and 11-hydroxy lauric acid by CYP2C9 variants. Metabolic incubations were carried out in duplicate and kinetic parameters were obtained as detailed under Materials and Methods. Duplicate velocity measurements varied less than 10% from the average when using (S)-warfarin and diclofenac as substrates, and by less than 18% for lauric acid incubations.
Discussion
The current study describes the identification and characterization of a new allelic variant in exon 7 of theCYP2C9 gene that is expressed in African Americans. A C-to-G transversion at nucleotide position 1080 causes a mis-sense mutation at codon 360, which results in substitution of an Asp residue with Glu. This relatively conservative change nonetheless substantially reduces the catalytic efficiency of the translated product, CYP2C9*5.
Codons 359 and 360 seem to define a “hot-spot” for mutation in theCYP2C9 gene (Table 3). In addition to the CYP2C9*3 and CYP2C9*5 alleles, which cause the functionally defective Leu359 and Glu360 mutations, the recently described CYP2C9*4 allele codes for a Thr359 mutation that significantly reduces enzyme activity both in vitro and in vivo (Ieiri et al., 2000; Imai et al., 2000). Interestingly, a nonconservative mutation of Cys for the naturally occurring Tyr at adjacent position 358 had no discernible effect on oxazaphosphorine metabolism (Chang et al., 1997). It is difficult to rationalize these diverse structural effects on CYP2C9 function; however, it is notable that amino acids 359 and 360 lie within one of the substrate recognition sites defined by Gotoh (1992), and that side chain size has been considered to be a major determinant of substrate specificity within such recognition sites (Negishi et al., 1996).
CYP2C9 sequence variation at codons 359 and 360 representing theCYP2C9*3, *4, and *5 alleles.
The current data, based on the frequency of five discreteCYP2C9 alleles in African American and European American populations, indicate that only the CY2C9*2 andCYP2C9*3 variants are of significance in European American subjects. Indeed, this study and others (Stubbins et al., 1996;Sullivan-Klose et al., 1996; Yasar et al., 1999) suggest that, on average, some 30% of whites are carriers of one or more defective alleles for CYP2C9. In contrast, less than 10% of African Americans carry defective alleles and these are distributed in a roughly equal manner between the CYP2C9*2,CYP2C9*3, and CYP2C9*5 variants.
In vitro, the CYP2C9*2 and CYP2C9*3 proteins exhibit reduced intrinsic metabolic capabilities relative to the wild-type enzyme. The magnitude of this effect is greater for CYP2C9*3 (Yamazaki et al., 1998; Rettie et al., 1999). In vivo, the *2/*3 and *3/*3 genotypes are associated with the tolbutamide poor metabolizer phenotype (Sullivan-Klose et al., 1996), and the *3/*3 genotype is clearly linked to warfarin sensitivity (Steward et al., 1997), impaired metabolism of losartan (McCrea et al., 1999), and substantially reduced oral clearances of glipizide and phenytoin (Kidd et al., 1999). Recently, a strong association has been demonstrated between the presence of the CYP2C9*3 allele and a low dose requirement for the warfarin congener, acenocoumarol (Thijssen et al., 2000). In addition, patients with CYP2C9 *1/*2 and*1/*3 genotypes exhibit a graded reduction in their maintenance dose of warfarin compared withCYP2C9*1 homozygotes (Aithal et al., 1999), in line with the relative catalytic efficiencies of CYP2C9*1, CYP2C9*2, and CYP2C9*3 toward (S)-warfarin (Yamazaki et al., 1998; Rettie et al., 1999). Therefore, there seems to be a reasonable correlation between the in vitro activities of these variant forms of CYP2C9 and the in vivo consequences for the expression of the alleles.
Interestingly, the relative reduction in CYP2C9*5-mediated intrinsic clearance differed among the three substrates, from 6-fold for diclofenac 4′-hydroxylation to 31-fold for the 7-hydroxylation ofS-warfarin. This variation has also been shown in vitro for CYP2C9*3. In a recent study involving seven CYP2C9 substrates, the reduction in intrinsic clearance produced by the Ile359Leu substitution ranged from 3-fold for diclofenac 4′-hydroxylation to 27-fold for piroxicam 5′-hydroxylation (Takanashi et al., 2000). Thus, the magnitude of impairment of oxidative metabolism caused by expression of the CYP2C9*3 and CYP2C9*5 variant proteins may be substrate dependent.
In summary, CYP2C9*5, a new variant allele, has been identified among African Americans. Purified, recombinant CYP2C9*5 exhibited reduced catalytic efficiencies toward (S)-warfarin, diclofenac, and lauric acid similar to those measured for CYP2C9*3. If the in vitro-in vivo correlations noted above for CYP2C9*2 and CYP2C9*3 can be extended to CYP2C9*5, carriers of theCYP2C9*5 allele are likely to be at a risk level similar to that of carriers of the CYP2C9*3 allele in terms of reduced clearance of CYP2C9 substrates. However, this hypothesis remains to be tested in vivo and, until confirmed, the predicted clinical consequences of expression of the CYP2C9*5 allele should be viewed with caution.
Acknowledgments
We acknowledge Rommel Tirona, Edna F. Choo, and Atsuko Suzuki for CYP2C9*2 genotyping, and the Molecular Biomarkers Laboratory in the Center for Ecogenetics and Environmental Health at the University of Washington.
Footnotes
- Received February 20, 2001.
- Accepted May 14, 2001.
-
↵1 CYP2C9 allele submitted to and number designated by the Human Cytochrome P450 Allele Nomenclature Committee.
-
This work was supported in part by United States Public Health Service Grants GM31304 and GM32165. The Molecular Biomarkers Laboratory in the Center for Ecogenetics and Environmental Health at the University of Washington is supported by National Institute of Environmental Health Sciences Grant ES07033.
-
Portions of this work have been reported in abstract form: Schwarz UI, Choo EF, Dresser GK, Stein CM, Wood AJJ, Roden DM, Wilkinson GR and Kim RB (2000) Identification of a new CYP2C9 variant in African-Americans. Clin Pharmacol Ther 67:169.
-
↵2 CYP2C9 variant proteins (CYP2C9*1, CYP2C9*2, etc.) are named according to the recommendations made by the Human Cytochrome P450 Allele Nomenclature Committee (http://www.imm.ki.se/CYPalleles).
Abbreviations
- P450
- cytochrome P450
- SNP
- single nucleotide polymorphism
- PCR-RFLP
- polymerase chain reaction-restriction fragment length polymorphism
- SSCP
- single-strand conformational polymorphism
- TBE
- Tris/borate/EDTA
- MALDI-TOF
- matrix-assisted laser desorption ionization time-of-flight
- HPLC
- high-performance liquid chromatography
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}