


Abstract
Mouse CYP2J5 is abundant in kidney and active in the metabolism of arachidonic acid to epoxyeicosatrienoic acids. Western blots of microsomes prepared from mouse kidneys demonstrate that after puberty, CYP2J5 protein is present at higher levels in male mice than in female mice. Northern analysis reveals that CYP2J5 transcripts are more abundant in adult male versus female kidneys, indicating that gender differences in renal CYP2J5 expression are regulated at a pretranslational level. Castration of male mice results in decreased renal CYP2J5 expression, and treatment of castrated male mice or female mice with 5α-dihydrotestosterone increases expression to levels that approximate those in intact male mice. In contrast, treatment of ovariectomized female mice or castrated male mice with 17β-estradiol causes a further reduction in CYP2J5 expression. Growth hormone-deficient (lit/lit) mice respond similarly to castration and 5α-dihydrotestosterone treatment, indicating that the androgen effects are not mediated by alterations in the growth hormone secretory pattern. Mice that lack a functional androgen receptor (Tfm hemizygous) have reduced levels of renal CYP2J5 and do not respond to 5α-dihydrotestosterone treatment. Similarly, wild-type male mice treated with flutamide, an androgen antagonist, exhibit reduced renal CYP2J5 levels. Female estrogen receptor-α knockout (αERKO) mice, which are known to have elevated circulating testosterone levels, have significantly increased renal CYP2J5 expression compared with wild-type female mice, and these differences are abrogated by ovariectomy or treatment with flutamide. Based on these data, we conclude that the renal expression of CYP2J5 is up-regulated by androgen and down-regulated by estrogen.
Cytochromes P450 (P450s) are heme-containing enzymes that have been the focus of study by toxicologists and pharmacologists because of their ability to catalyze the metabolism of a wide range of xenobiotics, including clinically used drugs, environmental pollutants, and chemical carcinogens (Nelson et al., 1996). A subset of these P450 enzymes also metabolize endogenous substrates such as arachidonic acid to compounds that act as signaling molecules that affect fundamental biologic processes (Zeldin, 2001; Roman, 2002). Among the bioactive eicosanoids that can be formed are the cis-epoxyeicosatrienoic acids (EETs), which are converted to their corresponding vic-dihydroxyeicosatrienoic acids by soluble epoxide hydrolase (Zeldin et al., 1993). Biological activities that have been attributed to the EETs and/or the dihydroxyeicosatrienoic acids include effects on peptide hormone secretion, vascular smooth muscle tone, cellular proliferation, inflammation, and hemostasis (Zeldin, 2001; Roman, 2002). EETs have also been shown to contribute to integrated kidney function, either directly by affecting renal tubular transport processes and vascular tone or indirectly by mediating the actions of several renal hormones, including renin, angiotensin II and arginine vasopressin (Zeldin, 2001; Roman, 2002). Importantly, existing data suggests a role for the renal P450 arachidonic acid epoxygenase pathway in the pathogenesis of hypertension in humans and rodents (Zeldin, 2001; Roman, 2002).
Hepatic EET biosynthesis can be altered by factors that affect P450 enzyme expression. For example, treatment of rats with phenobarbital induces liver CYP2B1/2 and increases hepatic arachidonic acid epoxygenase activity (Capdevila et al., 1990). Similarly, treatment of fish or avian species with aromatic hydrocarbons, such as benzo[a]pyrene or 2,3,7,8-tetrachlorodibenzo-p-dioxin, increases EET biosynthesis, probably by inducing liver CYP1A subfamily P450s (Gilday et al., 1998; Schlezinger et al., 1998). In contrast, fasting of rats leads to reduced liver CYP2C11 expression and decreased hepatic arachidonic acid epoxygenase activity (Qu et al., 1998). Much less is known about factors that regulate expression of kidney P450 epoxygenases. Dietary salt has been shown to induce rat CYP2C23, resulting in increased renal EET biosynthesis and enhanced urinary excretion of epoxygenase metabolites (Holla et al., 1999; Oyekan et al., 1999). Conversely, sodium nitroprusside, an NO donor, inhibited renal microsomal conversion of arachidonic acid to EETs (Oyekan et al., 1999).
A large number of studies have documented a role for sex hormones in regulating the expression of liver P450s (Morgan et al., 1985; Waxman et al., 1985; McClellan-Green et al., 1989; Thummel and Schenkman, 1990; Legraverend et al., 1992). Indeed, hepatic metabolism of xenochemicals is known to be sexually dimorphic and metabolism by sex-specific P450s leads to sex-dependent susceptibility to chemical toxicity and/or carcinogenicity (Waxman and Chang, 1995). It is now well established that sexual dimorphism of hepatic P450s is regulated, in part, by growth hormone secretory patterns (McClellan-Green et al., 1989; Legraverend et al., 1992; Mode et al., 1992; Park et al., 1999). Pulsatile growth hormone release stimulates expression of male-specific P450s, whereas a continuous growth hormone secretory pattern suppresses expression of male-specific P450s and stimulates expression of female-specific P450s. A number of mechanisms have been proposed to explain the sexually dimorphic effects of growth hormone on liver P450 gene transcription, including phospholipase A2 activation and subsequent P450-catalyzed formation of an active arachidonic acid metabolite (Tollet et al., 1995), activation of the signal transducer and activation of transcription STAT5b (Udy et al., 1997), and regulation of STAT5b nuclear translocation by the estrogen receptor-α (Sueyoshi et al., 1999).
In contrast, much less is known about the regulation of renal P450s by sex hormones. Sexual dimorphism has been reported in the expression of several different kidney P450s, including members of the CYP2A, CYP2B, CYP2C, CYP2E, CYP3A, CYP4A, and CYP4B subfamilies (Henderson et al., 1990; Henderson and Wolf, 1991; Heng et al., 1997; Holla et al., 2001). These effects seem to be mediated by testosterone, which has the capacity to both induce and repress P450 gene expression (Henderson et al., 1990). As in the liver, sexual dimorphism of kidney P450s is thought to contribute to susceptibility to renal toxic and carcinogenic effects of certain drugs (e.g., acetaminophen) and environmentally relevant chemicals (e.g., nitrosodimethylamine) (Hong et al., 1989; Hoivik et al., 1995).
Previously, we reported the cDNA cloning, heterologous expression, and enzymatic characterization of CYP2J5, a new mouse P450 that is abundant in kidney and active in the metabolism of arachidonic acid to EETs (Ma et al., 1999). In light of evidence that 1) renal EET biosynthesis may be important in the pathogenesis of hypertension in both rats and humans (Zeldin, 2001; Roman, 2002), 2) gender differences in the incidence and severity of hypertension occur in humans (Chen, 1996), and 3) disruption of the Cyp2j5 gene causes spontaneous, sexually dimorphic hypertension in mice (J. Ma, K. Athirakul, T. Coffman, and D. C. Zeldin, unpublished observations), we examined the extent of sexual differentiation of CYP2J5. Herein, we report that renal CYP2J5 expression is higher in male mice than in female mice after puberty. Correspondingly, adult mouse renal arachidonic acid epoxygenase activity is also higher in male mice. Mechanistic studies involving castration/ovariectomy with/without hormone replacement, androgen receptor-deficient (Tfm hemizygous) and estrogen receptor-deficient (αERKO) mice, and growth hormone deficient (lit/lit) mice demonstrate that renal CYP2J5 levels are regulated at the pretranslational level by both androgens and estrogens but that this regulation does not depend on growth hormone.
Materials and Methods
Materials. [α-32P]dATP and [1-14C]arachidonic acid were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). Timed-release (60-day) 5α-dihydrotestosterone (DHT), 17β-estradiol (EST), flutamide, and corresponding placebo pellets were purchased from Innovative Research of America (Sarasota, FL). All other chemicals and reagents were purchased from Sigma-Aldrich Chemical Company (St. Louis, MO) unless otherwise specified.
Experimental Animals. All animal studies were conducted in accordance with principles and procedures outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals and approved by the National Institute of Environmental Health Sciences Committee on Animal Care and Use. Kidney and liver tissues were obtained from male and female C57BL/6J mice (aged from fetal day 18 to 16 weeks postnatal) purchased from Hilltop Laboratories (Scottsdale, PA). Animals were weaned at 3 weeks of age, fed National Institutes of Health 31 rodent chow (Agway, St. Mary, OH) ad libitum thereafter, and sacrificed by lethal CO2 inhalation. For some experiments, mice were castrated or ovariectomized at 8 weeks of age (or subjected to sham castration or ovariectomy) and then treated with subcutaneous 60-day timed-release pellets containing either DHT (15 mg/pellet), EST (0.18 mg/pellet), flutamide (25 mg/pellet), or placebo for 3 to 4 weeks before sacrifice at 11 to 12 weeks of age. For other experiments, mice were castrated or ovariectomized 24 h after birth and treated with either subcutaneous injection of testosterone propionate (125 μg) or vehicle (corn oil) on days 1 and 3, or treated with subcutaneous 60-day timed-release pellets containing DHT (15 mg/pellet), EST (0.18 mg/pellet), or placebo at 8 weeks of age for 3 to 4 weeks before sacrifice at 11 to 12 weeks of age. Mice with targeted disruption of estrogen receptor-α (homozygous αERKO) (Lubahn et al., 1993) and corresponding wild-type control mice were obtained from Taconic (Germantown, NY). Androgen receptor mutant mice (Tfm hemizygous) (Lyon and Glenister, 1974) and corresponding wild-type control mice were purchased from The Jackson Laboratory (Bar Harbor, MA). Growth hormone-releasing hormone receptor homozygous mutant mice (lit/lit) (Jansson et al., 1986) and corresponding heterozygous control mice (lit/+) were also purchased from The Jackson Laboratory.
Protein Immunoblotting. Kidneys and livers were perfused in situ with ice-cold phosphate-buffered saline, dissected free of surrounding tissue, frozen in liquid nitrogen, and stored at -80°C until use. Microsomal fractions were prepared by differential centrifugation at 4°C as described previously (Ma et al., 1999). Recombinant CYP2J2, CYP2J3, CYP2J5, CYP2J6, CYP2J8, and CYP2J9 proteins were prepared in insect cells using the baculovirus system as described previously (Wu et al., 1996, 1997; Ma et al., 1999, 2002; Qu et al., 2001). Polyclonal antibodies against the CYP2J5-specific peptide HLDQNFVNRFMTPVR (anti-CYP2J5pep IgG) were raised in New Zealand white rabbits using methods published previously (Ma et al., 1999). This antibody has been shown to strongly react with recombinant mouse CYP2J5 but does not cross-react with other mouse P450s, including CYP2A4, CYP2A5, CYP2C29, CYP2C37, CYP2C38, CYP2C39, and CYP2C40 (Ma et al., 1999). For immunoblotting, microsomal fractions or recombinant P450s were electrophoresed in 8 to 16% Tris glycine gels (80 × 80 × 1 mm) purchased from Novex (San Diego, CA), and the resolved proteins were transferred electrophoretically onto nitrocellulose membranes. Membranes were immunoblotted using rabbit anti-CYP2J5pep IgG, goat anti-rabbit IgG conjugated to horseradish peroxidase (Bio-Rad, Richmond, CA), and the ECL Western blotting detection system (Amersham Biosciences, Piscataway, NJ) as described previously (Ma et al., 1999). Densitometry was performed on autoradiographs using a ChemiImager 4000 Imaging System (Alpha Innotech Corp., San Leandro, CA). Protein determinations were performed using reagents from Bio-Rad. Preimmune serum, collected from the rabbits before immunization, did not cross-react with CYP2J5 or with microsomal fractions prepared from mouse tissues.
Northern Blot Hybridization. RNA was prepared from frozen mouse tissues using the RNeasy Midi-prep kit from QIAGEN (Valencia, CA). Total RNA (30 μg) was denatured and electrophoresed in 1.2% agarose gels containing 2.2 M formaldehyde. After capillary-pressure transfer to Hybond-N+ membranes (Amersham Biosciences), the blots were hybridized with the 1.9-kilobase CYP2J5 cDNA probe (GenBank accession no. U62294) as described previously (Ma et al., 1999). The cDNA probe was gel-purified using a Qiaex gel extraction kit (QIAGEN), labeled with [α-32P]dATP using the Megaprime DNA labeling system (Amersham Biosciences), and purified using G-50 Sephadex columns (Stratagene, La Jolla, CA). Autoradiographs were scanned and relative CYP2J5 mRNA levels were determined by normalization to the 28S rRNA signal.
Quantitative Real-Time TaqMan RT-PCR Analysis. CYP2J5 mRNA levels were quantified by TaqMan real-time reverse transcription-polymerase chain reaction (RT-PCR). First-strand cDNA was synthesized from 2.5 μg of total RNA using a high-capacity cDNA archive kit (Applied Biosystems, Foster City, CA). PCR was performed using TaqMan universal PCR master mix on the ABI Prism 7700 sequence detection system (Applied Biosystems,). CYP2J5 specific probe and primers (assay ID Mm00487292_ml) were designed and synthesized by Applied Biosystems. Reactions were performed in quadruplicate in a MicroAmp optical 96-well plate in a total volume of 50 μl. Each reaction mixture contained 200 μM concentrations of dATP, dCTP, and dGTP, 400 μM dUTP, 5.5 mM MgCl2, 1.25 units of AmpliTaq Gold DNA polymerase, 300 nM concentrations of each primer, 100 nM probe, and 50 ng of reverse-transcribed RNA. PCR conditions were as follows: 50°C for 2 min; 95°C for 10 min; 95°C for 15 s, 60°C for 1 min (40 cycles). Each assay plate contained unknown samples, no template controls, and serially diluted concentrations of cDNA sample from which a standard curve was generated.
Incubations of Mouse Kidney and Liver Microsomes with Arachidonic Acid. Microsomal fractions were prepared from freshly isolated adult male and female mouse kidneys and livers. Reaction mixtures containing 0.05 M Tris-Cl buffer, pH 7.5, 0.15 M KCl, 0.01 M MgCl2, 8 mM sodium isocitrate, 0.5 IU/ml isocitrate dehydrogenase, 1 mM NADPH, 100 μM [1-14C]arachidonic acid, and 1 to 2 mg/ml microsomal protein were incubated at 37°C with constant mixing. At different time points, aliquots were withdrawn, and the reaction products were extracted into diethyl ether and analyzed by reverse-phase high-performance liquid chromatography as described previously (Wu et al., 1996; Ma et al., 1999; Qu et al., 2001). All products were identified by comparing their high-performance liquid chromatography properties with those of authentic EET and hydroxyeicosatetraenoic acid standards.
Statistical Analysis. All values are expressed as mean ± S.E. Data were analyzed either using the Student's t test (two-tailed distribution, assuming equal variances) or by analysis of variance using SYSTAT software (SYSTAT Inc., Evanston, IL). When F values indicated that a significant difference was present, Fisher's least significant difference post hoc test for multiple comparisons was used. Values were considered significantly different if p was <0.05.
Results
Expression of CYP2J5 Protein in Male and Female Kidneys and Livers. We have shown previously that CYP2J5 is abundant in mouse kidney and present at lower levels in mouse liver (Ma et al., 1999). More recently, we documented the presence of other CYP2J proteins in kidney and/or liver including human CYP2J2, rat CYP2J3, and mouse CYP2J6, CYP2J8, and CYP2J9 (Wu et al., 1996, 1997; Qu et al., 2001; Ma et al., 2002). Immunoblots of lysates prepared from insect cells expressing recombinant CYP2J proteins demonstrated that the anti-CYP2J5pep IgG reacted strongly with recombinant mouse CYP2J5 but did not cross-react with other CYP2J isoforms (Fig. 1). This antibody also did not cross-react with non-CYP2J P450s, including members of the CYP2A and CYP2C subfamilies (Ma et al., 1999). Based on these data, we conclude that anti-CYP2J5pep IgG is immunospecific for CYP2J5.

Immunoreactivity of anti-CYP2J5pep IgG. Microsomal fractions prepared from insect cells expressing recombinant CYP2J2, CYP2J3, CYP2J5, CYP2J6, CYP2J8, and CYP2J9 (1 pmol of P450/lane) were electrophoresed on Tris-glycine gels, and the resolved proteins were transferred to a nitrocellulose membrane. The membrane was then immunoblotted with the anti-CYP2J5pep IgG and goat anti-rabbit IgG conjugated to horseradish peroxidase as described under Materials and Methods. The immunoreactive proteins were visualized using the enhanced chemiluminescence detection system and autoradiography.
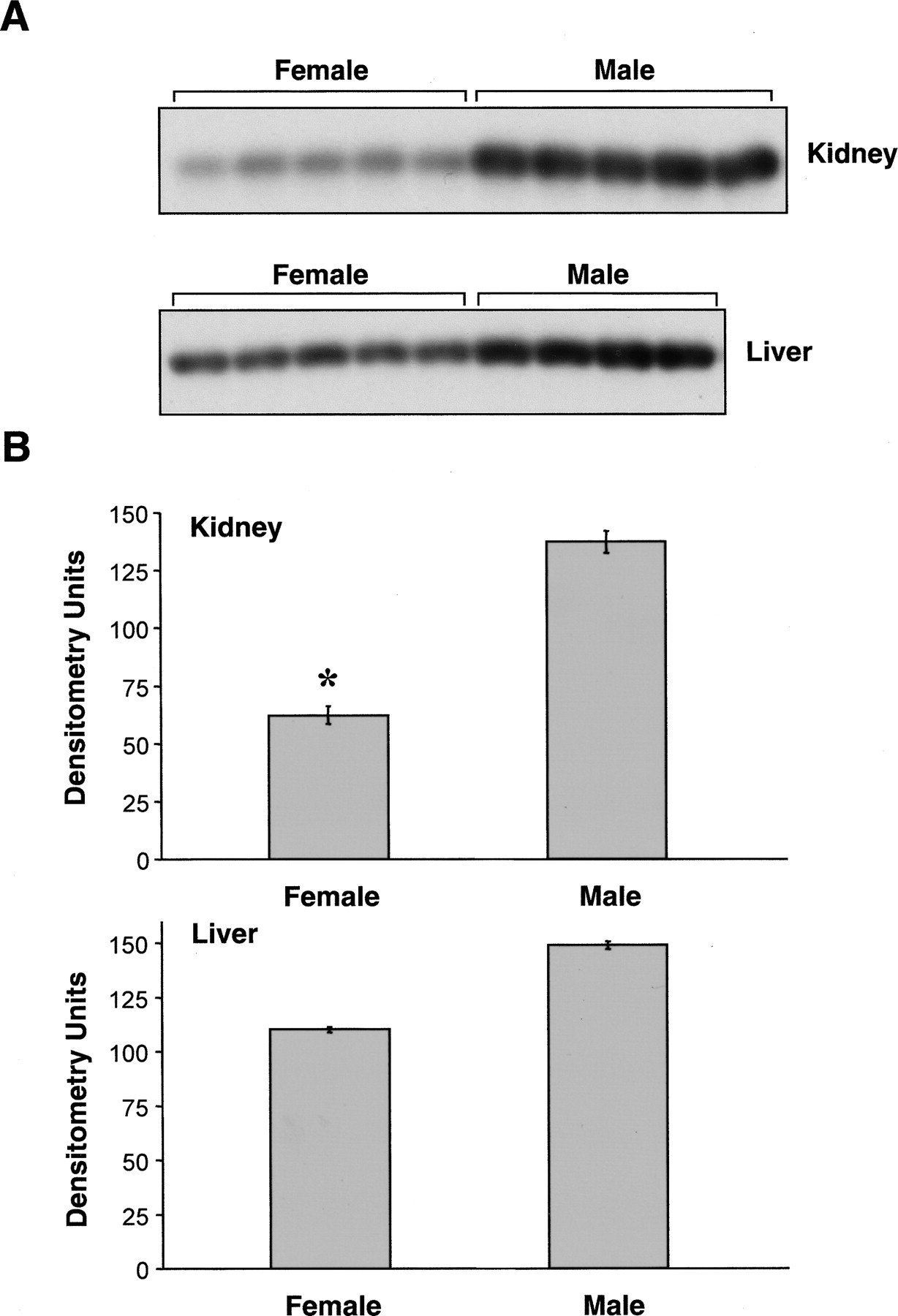
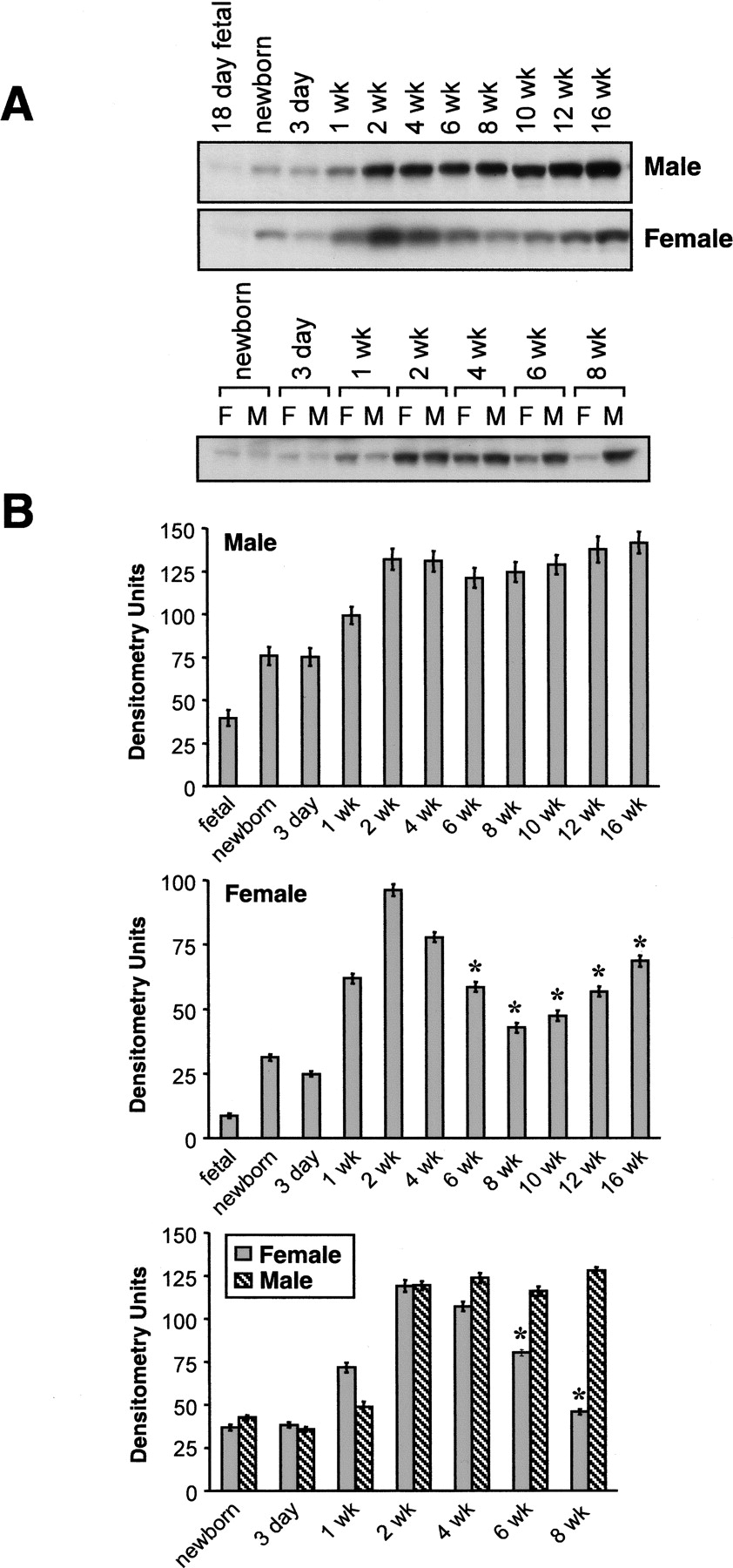
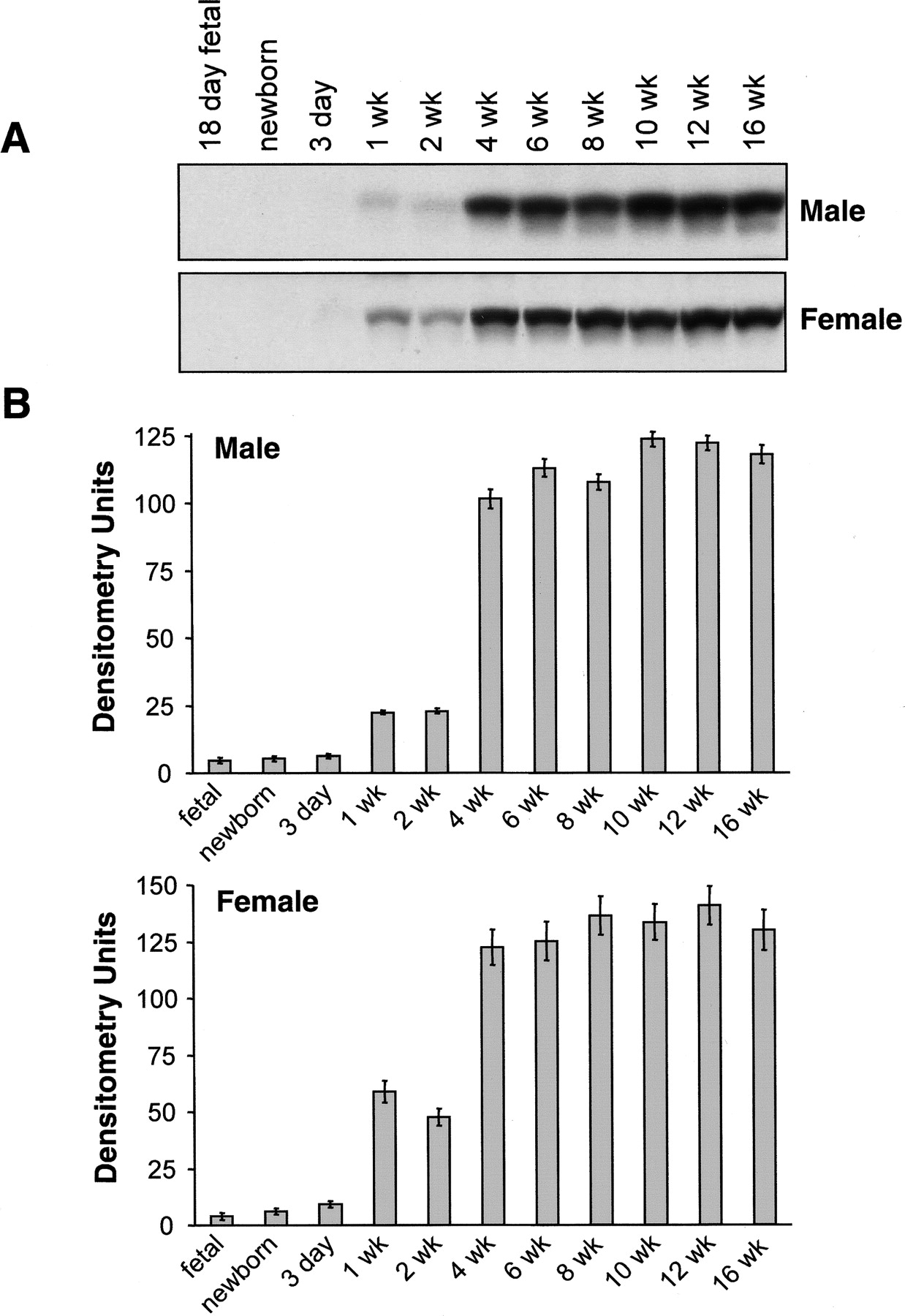
To examine whether there are gender differences in the expression of CYP2J5 protein, we immunoblotted male and female mouse kidney and liver microsomes with the anti-CYP2J5pep IgG. As shown in Fig. 2, A and B, CYP2J5 expression was significantly higher in adult male versus adult female mouse kidney. In contrast, these gender differences in CYP2J5 protein expression were much less pronounced in adult mouse liver. To determine the influence of age on CYP2J5 expression in male and female mice, we immunoblotted tissues collected from mice aged from fetal day 18 to 16 weeks' postnatal. Interestingly, we found that renal CYP2J5 protein levels were comparable in male and female mice during the prepubertal period (Fig. 3, A and B). After 4 weeks of age (postpuberty), male CYP2J5 expression remained relatively constant, whereas female CYP2J5 expression decreased markedly, achieving minimal levels at 8 weeks of age. In contrast, the expression of CYP2J5 was comparable at all ages (both pre- and postpubertal periods) in male and female mouse liver (Fig. 4, A and B). Taken together, these data indicate that kidney CYP2J5 protein is present at higher levels in male mice versus female mice after puberty, whereas liver CYP2J5 expression is comparable in male and female mice at all ages.
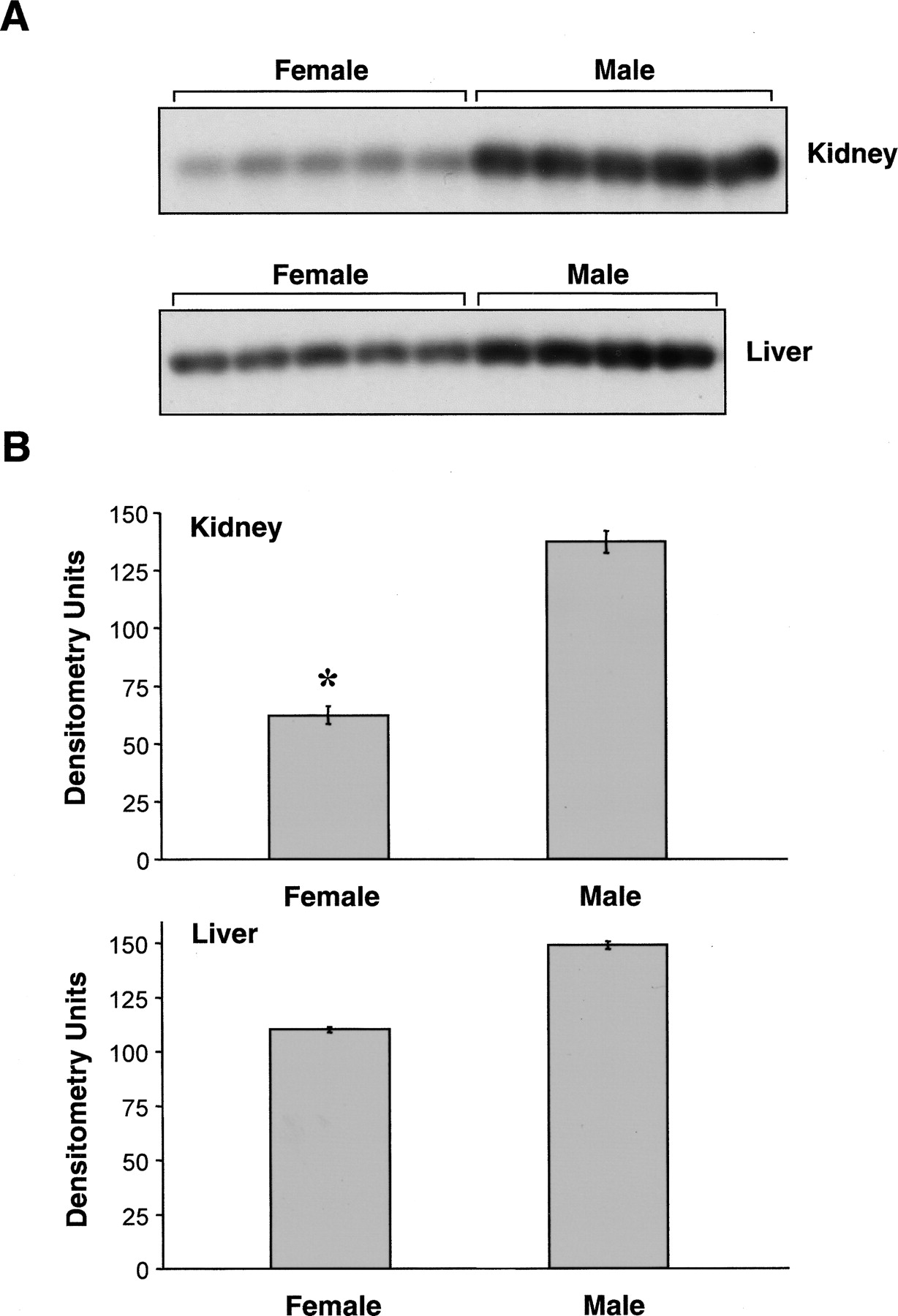
Expression of CYP2J5 protein in male and female mouse kidney and liver. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys and livers of male and female mice (n = 4–5 animals per group) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The kidney immunoblot was exposed for 1 min and the liver immunoblot was exposed for 3 min. The immunoblots shown are each representative of three independent determinations. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 4 to 5 animals per group with the value for each animal taken as the average of the three determinations. *, p < 0.05 versus male kidney.

Influence of age on expression of CYP2J5 protein in male and female kidneys. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys of male and female mice (age from fetal day 18 to 16 weeks postnatal) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblots were exposed for 1 min. Each microsomal sample represents kidney tissues pooled from n = 3 to 10 animals. The immunoblots shown are each representative of three independent determinations with these pooled samples. M, male; F, female. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of three determinations. *, p < 0.05 versus male kidney.

Influence of age on expression of CYP2J5 protein in male and female livers. A, microsomal fractions (30 μg of protein/lane) prepared from livers of male and female mice (age from fetal day 18 to 16 weeks postnatal) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblots were exposed for 3 min. Each microsomal sample represents liver tissues pooled from n = 3 to 10 animals. The immunoblots shown are each representative of three independent determinations with these pooled samples. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of three determinations.
CYP2J5 mRNA Abundance in Male and Female Kidneys and Livers. To examine if the differences in CYP2J5 protein expression between male and female mice were accompanied by corresponding changes in CYP2J5 mRNA levels, we performed Northern analysis. In agreement with the immunoblotting results, we found that CYP2J5 mRNA abundance was significantly higher in adult male versus adult female kidneys (Fig. 5, A and B), indicating that gender differences in renal CYP2J5 expression are probably regulated at a pretranslational level. In contrast, CYP2J5 mRNA was slightly higher in adult female versus adult male livers (Fig. 5, C and D). These changes in mRNA abundance were not a result of differences in the amount of RNA applied to each lane, as assessed by ethidium bromide staining of the gel before transfer or the membrane after transfer.

Abundance of CYP2J5 mRNAs in male and female kidneys and livers. A, total RNA (30 μg/lane) prepared from kidneys of male and female mice (n = 5 animals per group) was denatured, electrophoresed, transferred to Hybond-N+ membranes and blot hybridized with the radiolabeled CYP2J5 cDNA as described under Materials and Methods. Top, autoradiograph of blot after 24-h exposure time. Bottom, ethidium bromide-stained gel before transfer. B, kidney autoradiographs were scanned and relative CYP2J5 mRNA levels were determined. Values shown are the means ± S.E. of n = 5 animals per group. *, p < 0.05 versus male kidney. C, total RNA (30 μg/lane) prepared from livers of male and female mice (n = 5 animals per group) was denatured, electrophoresed, transferred, and blot hybridized as described. Top, autoradiograph of blot after 48-h exposure time. Bottom, ethidium bromide-stained gel before transfer. D, liver autoradiographs were scanned and relative CYP2J5 mRNA levels were determined. Values shown are the means ± S.E. of n = 5 animals per group.
Metabolism of Arachidonic Acid by Male and Female Mouse Kidneys and Livers. We have shown previously that CYP2J5 is an active arachidonic acid epoxygenase and probably contributes to EET biosynthesis in mouse kidney and liver (Ma et al., 1999). To examine whether the gender differences in CYP2J5 protein expression were accompanied by corresponding changes in total epoxygenase activity, we incubated microsomal fractions prepared from adult male and female mouse tissues with radiolabeled arachidonic acid. As shown in Table 1, renal microsomal arachidonic acid epoxygenase activity was ∼2-fold higher in male mice versus female mice. In contrast, there was no significant difference between male and female mice in liver microsomal arachidonic acid epoxygenase activity (Table 1).
Epoxygenase activity in male and female kidney and liver microsomes The total epoxygenase activity of microsomes prepared from adult male and female kidneys and livers was determined as described under Materials and Methods. Values shown are means ± S.E. (n = 3–4 per group) and are expressed in picomoles of EET + dihydroxyeicosatrienoic acid formed per milligram of microsomal protein per minute at 37° C.
Effects of Androgen on Renal CYP2J5 Expression. The increased expression of CYP2J5 in male versus female kidneys after puberty suggests the possibility of an androgen-mediated effect. Indeed, the mouse kidney is known to be an androgen responsive tissue (Henderson et al., 1990; Henderson and Wolf, 1991; Holla et al., 2001). Treatment of adult female mice with pellets containing DHT for 3 to 4 weeks significantly increased renal CYP2J5 protein expression to levels that approached those present in intact adult male mice (Fig. 6, A and B). Likewise, castration of male mice at 8 weeks of age resulted in a reduction in renal CYP2J5 protein expression, an effect that was reversed by treatment with DHT for 3 to 4 weeks (Fig. 6, A and B). To examine whether the effects of androgen on CYP2J5 expression were mediated at the pretranslational level, we determined CYP2J5 mRNA levels by quantitative real-time TaqMan RT-PCR. In agreement with the immunoblotting results, we found that renal CYP2J5 mRNA levels were significantly higher in male mice versus female mice, significantly lower in castrated male mice versus intact male mice, and significantly higher after treatment of female mice or castrated male mice with DHT (Fig. 6C). Together, these data suggest that renal CYP2J5 expression is regulated, at least in part, by androgen.

Effect of androgen on renal CYP2J5 protein expression in female and castrated male mice. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys of intact female mice treated with DHT pellets for 3 to 4 weeks (a), intact female mice treated with blank pellets (b), intact male mice treated with blank pellets (c), castrated male mice treated with DHT pellets for 3 to 4 weeks (d), and castrated male mice treated with blank pellets (e) (n = 3–5 animals per group) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of three independent determinations. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3–5 animals per group with the value for each animal taken as the average of two or three determinations. *, p < 0.05 versus intact male kidney; #, p < 0.05 versus DHT-treated intact female or DHT-treated castrated male kidney. C, total RNA prepared from kidneys of intact female mice treated with DHT pellets for 3 to 4 weeks (a), intact female mice treated with blank pellets (b), intact male mice treated with blank pellets (c), castrated male mice treated with DHT pellets for 3 to 4 weeks (d), and castrated male mice treated with blank pellets (e) was subjected to quantitative real-time TaqMan RT-PCR analysis as described under Materials and Methods. Values shown are the means ± S.E. of n = 3 animals per group. *, p < 0.05 versus intact male kidney; #, p < 0.05 versus DHT-treated intact female or DHT-treated castrated male kidney.
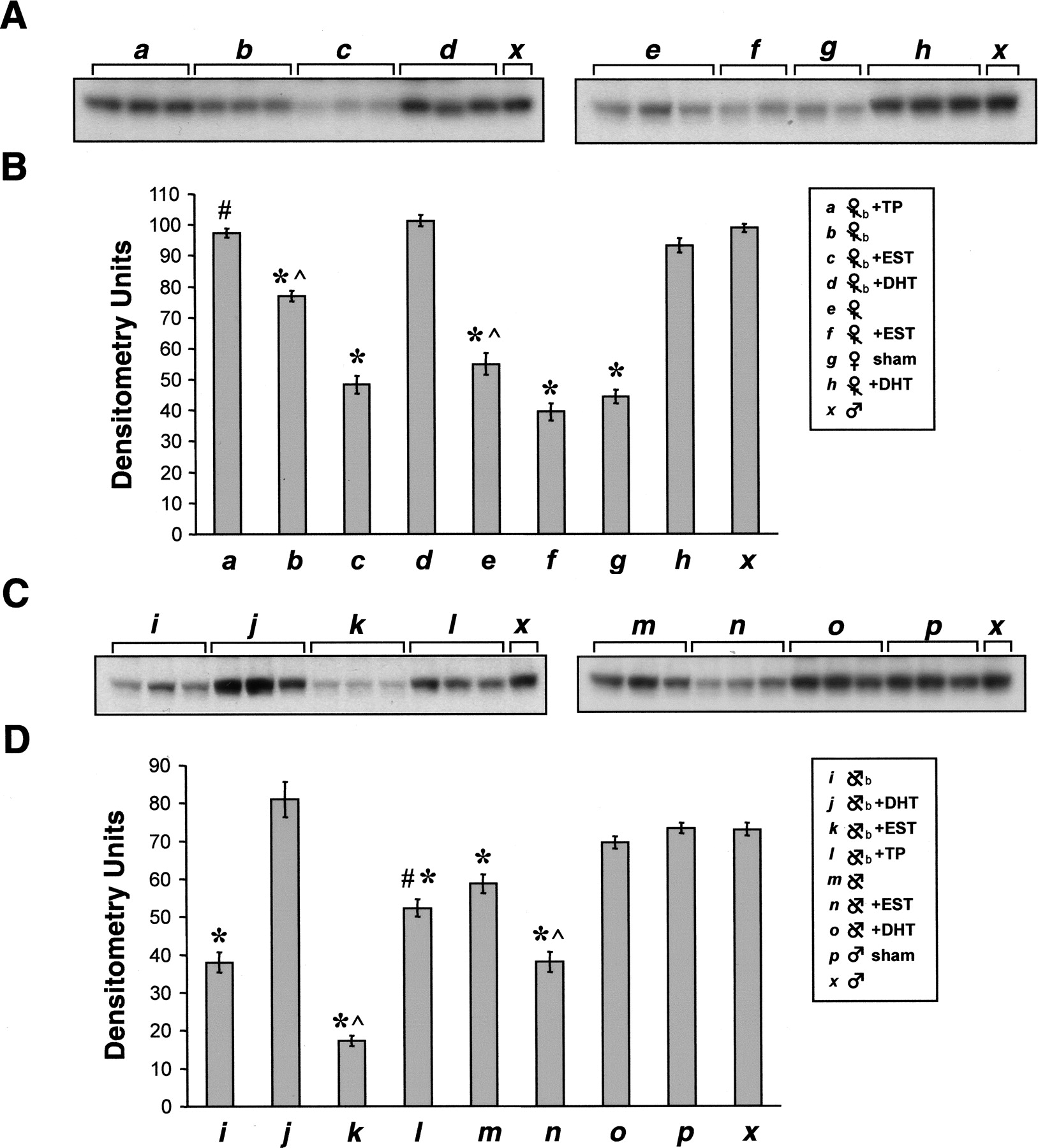
Neonatal actions of androgen have been shown to regulate sex-specific expression of P450 genes in adults (Wong et al., 1987; Waxman and Chang, 1995). Neonatally ovariectomized female mice treated with two subcutaneous injections of testosterone propionate on postnatal days 1 and 3 had significantly higher renal CYP2J5 expression as adults than neonatally ovariectomized female mice treated with vehicle on postnatal days 1 and 3 (Fig. 7, A and B). In fact, the level of renal CYP2J5 expression in neonatally ovariectomized female mice injected with testosterone propionate was comparable with that of intact male mice and ovariectomized female mice treated with DHT pellets for 3 to 4 weeks as adults (Fig. 7, A and B). Similarly, neonatally castrated male mice treated with two subcutaneous injections of testosterone propionate on postnatal days 1 and 3 had significantly higher renal CYP2J5 expression as adults than neonatally castrated male mice treated with vehicle on postnatal days 1 and 3 (Fig. 7, C and D). The effect of neonatal testosterone propionate injection on renal CYP2J5 expression in neonatally castrated male mice was less pronounced than the effect of 3 to 4 weeks of DHT treatment during adulthood (Fig. 7, C and D).

Effect of androgen and estrogen on renal CYP2J5 protein expression in female and male mice. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys of female mice treated in various ways (n = 3 animals per group) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. a, mice ovariectomized at birth and injected with testosterone propionate on days 1 and 3; b, mice ovariectomized at birth and injected with vehicle on days 1 and 3; c, mice ovariectomized at birth and treated with EST pellets for 3 to 4 weeks beginning at week 8; d, mice ovariectomized at birth and treated with DHT pellets for 3 to 4 weeks beginning at week 8; e, mice ovariectomized at week 8 and treated with blank pellets for 3 to 4 weeks; f, mice ovariectomized at week 8 and treated with EST pellets for 3 to 4 weeks; g, mice sham ovariectomized at week 8 and treated with blank pellets for 3 to 4 weeks; h, mice ovariectomized at week 8 and treated with DHT pellets for 3 to 4 weeks; x, intact untreated male mouse. The immunoblots shown are each representative of three independent determinations. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group with the value for each animal taken as the average of two or three determinations. *, p < 0.05 versus intact male; #, p < 0.05 versus vehicle injected ovariectomized female; ^, p < 0.05 versus intact female. C, microsomal fractions (30 μg of protein/lane) prepared from kidneys of male mice treated in various ways (n = 3 animals per group) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. i, mice castrated at birth and injected with vehicle on days 1 and 3; j, mice castrated at birth and treated with DHT pellet for 3 to 4 weeks beginning at week 8; k, mice castrated at birth and treated with EST pellets for 3 to 4 weeks beginning at week 8; l, mice castrated at birth and treated with testosterone propionate on days 1 and 3; m, mice castrated at week 8 and treated with blank pellets for 3 to 4 weeks; n, mice castrated at week 8 and treated with EST pellets for 3 to 4 weeks; o, mice castrated at week 8 and treated with DHT pellets for 3 to 4 weeks; p, mice sham castrated at week 8 and treated with blank pellets for 3 to 4 weeks; x, intact untreated male mouse. The immunoblots shown are each representative of three independent determinations. D, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group with the value for each animal taken as the average of two or three determinations. *, p < 0.05 versus intact male; #, p < 0.05 versus vehicle injected castrated male; ^, p < 0.01 versus vehicle-injected or blank pellet-treated castrated male.
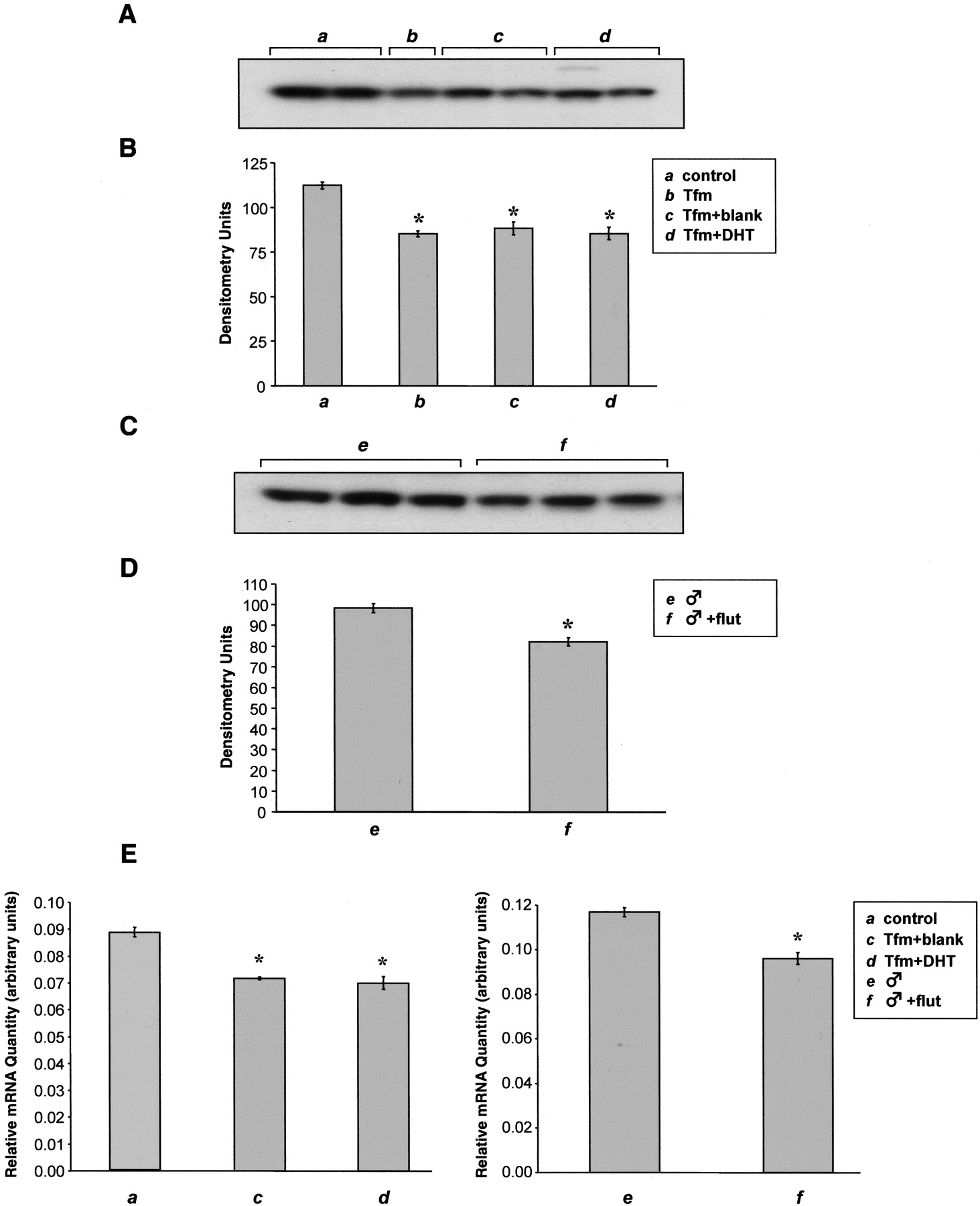
To evaluate the role of the androgen receptor in regulating renal CYP2J5 levels, we examined expression in testicular feminized (Tfm hemizygous) male mice, which carry a defect in the androgen receptor (Lyon and Glenister, 1974). Compared with control male mice, Tfm hemizygous mice had significantly reduced renal CYP2J5 protein expression (Fig. 8, A and B). Moreover, treatment of Tfm hemizygous mice with DHT for 3 to 4 weeks had no significant effect on renal CYP2J5 levels. Likewise, flutamide, an androgen receptor antagonist, caused a small but significant reduction in intact wild-type male renal CYP2J5 protein expression (Fig. 8, C and D). To examine whether these effects on CYP2J5 expression were mediated at the pretranslational level, we quantified CYP2J5 mRNA levels by real-time TaqMan RT-PCR. In agreement with the immunoblotting results, we found that renal CYP2J5 mRNA levels were significantly reduced in Tfm hemizygous mice (with or without DHT treatment) and significantly reduced after treatment with flutamide (Fig. 8E). Taken together, these data support a role for the androgen receptor in regulating renal CYP2J5 expression.

Effect of androgen receptor deficiency and flutamide on renal CYP2J5 protein expression. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys (n = 3 animals per group) of control male mice (a), Tfm hemizygous mice without treatment (b), Tfm hemizygous mice treated with blank pellet for 3 to 4 weeks (c), and Tfm hemizygous mice treated with DHT pellet for 3 to 4 weeks (d) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of three independent determinations. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group with the value for each animal taken as the average of two or three determinations. *, p < 0.05 versus control male. C, microsomal fractions (30 μg of protein/lane) prepared from kidneys (n = 3 animals per group) of intact male mice treated with blank pellets (e) and intact male mice treated with flutamide pellets for 3 to 4 weeks (f) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of three independent determinations. D, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group with the value for each animal taken as the average of the three determinations. *, p < 0.05 versus intact male treated with blank pellet. E, total RNA prepared from kidneys of control male mice (a), Tfm hemizygous mice treated with blank pellet for 3 to 4 weeks (c), Tfm hemizygous mice treated with DHT pellet for 3 to 4 weeks (d), intact male mice treated with blank pellets (e), and intact male mice treated with flutamide pellets for 3 to 4 weeks (f) was subjected to quantative real-time TaqMan RT-PCR analysis as described under Materials and Methods. Values shown are the means ± S.E. of n = 3 animals per group. *, p < 0.05 versus control male or intact male treated with blank pellet.
Effect of Growth Hormone on Renal CYP2J5 Expression. It is well established that sexual dimorphism of hepatic P450s is regulated by growth hormone secretory patterns (McClellan-Green et al., 1989; Legraverend et al., 1992; Mode et al., 1992; Park et al., 1999). Thus, pulsatile release of growth hormone stimulates expression of male-specific P450s, whereas continuous growth hormone release suppresses expression of male-specific P450s and stimulates expression of female-specific P450s. To investigate the potential role of growth hormone on renal CYP2J5 expression, we examined the effects of castration and DHT treatment in homozygous lit/lit mice, which are deficient in the pituitary receptor for growth hormone releasing hormone and have circulating growth hormone levels only 5 to 10% of normal (Jansson et al., 1986). Adult female lit/lit mice had significantly lower renal CYP2J5 expression than adult male lit/lit mice (Fig. 9, A and B). These differences are comparable with those between adult male and female lit/+ mice, which have normal circulating growth hormone levels. Treatment of adult female lit/lit mice with DHT pellets for 3 to 4 weeks significantly increased renal CYP2J5 expression (Fig. 9, A and B). The magnitude of these changes was comparable with those present in adult female lit/+ mice treated with DHT. Likewise, castration of adult male lit/lit mice resulted in a significant reduction in renal CYP2J5 expression, an effect that was reversed by treatment with DHT (Fig. 9, A and B). Together, these data suggest that the renal CYP2J5 expression is not significantly influenced by growth hormone secretory patterns.

Effect of androgen on renal CYP2J5 protein expression in homozygous growth hormone releasing hormone receptor-deficient (lit/lit) mice. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys (n = 3 animals per group) of intact lit/lit female mice treated with blank pellets (a), intact lit/lit female mice treated with DHT pellets for 3 to 4 weeks (b), castrated lit/lit male mice treated with blank pellets (c), castrated lit/lit male mice treated with DHT pellets for 3 to 4 weeks (d), intact lit/lit male mice treated with blank pellet (e), intact lit/+ female mouse treated with vehicle (f), intact lit/+ female mouse treated with DHT pellets for 3 to 4 weeks (g), and intact lit/+ male mouse treated with blank pellets (h) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of three independent determinations. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group, with the value for each animal taken as the average of two or three determinations. *, p < 0.05 versus intact lit/lit or lit/+ male; #, p < 0.05 versus nonDHT treated lit/lit or lit/+ female.
Effect of Estrogen on Renal CYP2J5 Expression. The reduced expression of CYP2J5 in female versus male kidneys after puberty could also be a result of the influence of estrogens. Indeed, Morgan et al. (1985) have shown that treatment of male rats with estradiol feminizes hepatic P45016α levels, and Sueyoshi et al. (1999) have recently shown that estrogen receptor-α feminizes hepatic CYP2A4 and CYP2D9 expression in mice. Ovariectomy of female mice, either during the neonatal period or at 8 weeks of age, increases renal CYP2J5 expression compared with intact female mice (Fig. 7, A and B). Likewise, treatment of ovariectomized mice with EST at 8 weeks of age for 3 to 4 weeks decreases renal CYP2J5 expression compared with ovariectomized mice treated with vehicle (Fig. 7, A and B). Similarly, treatment of castrated male mice with EST at 8 weeks of age for 3 to 4 weeks leads to a reduction in renal CYP2J5 expression compared with castrated mice treated with vehicle (Fig. 7, C and D). Together, these data suggest that renal CYP2J5 expression is also regulated by estrogen.
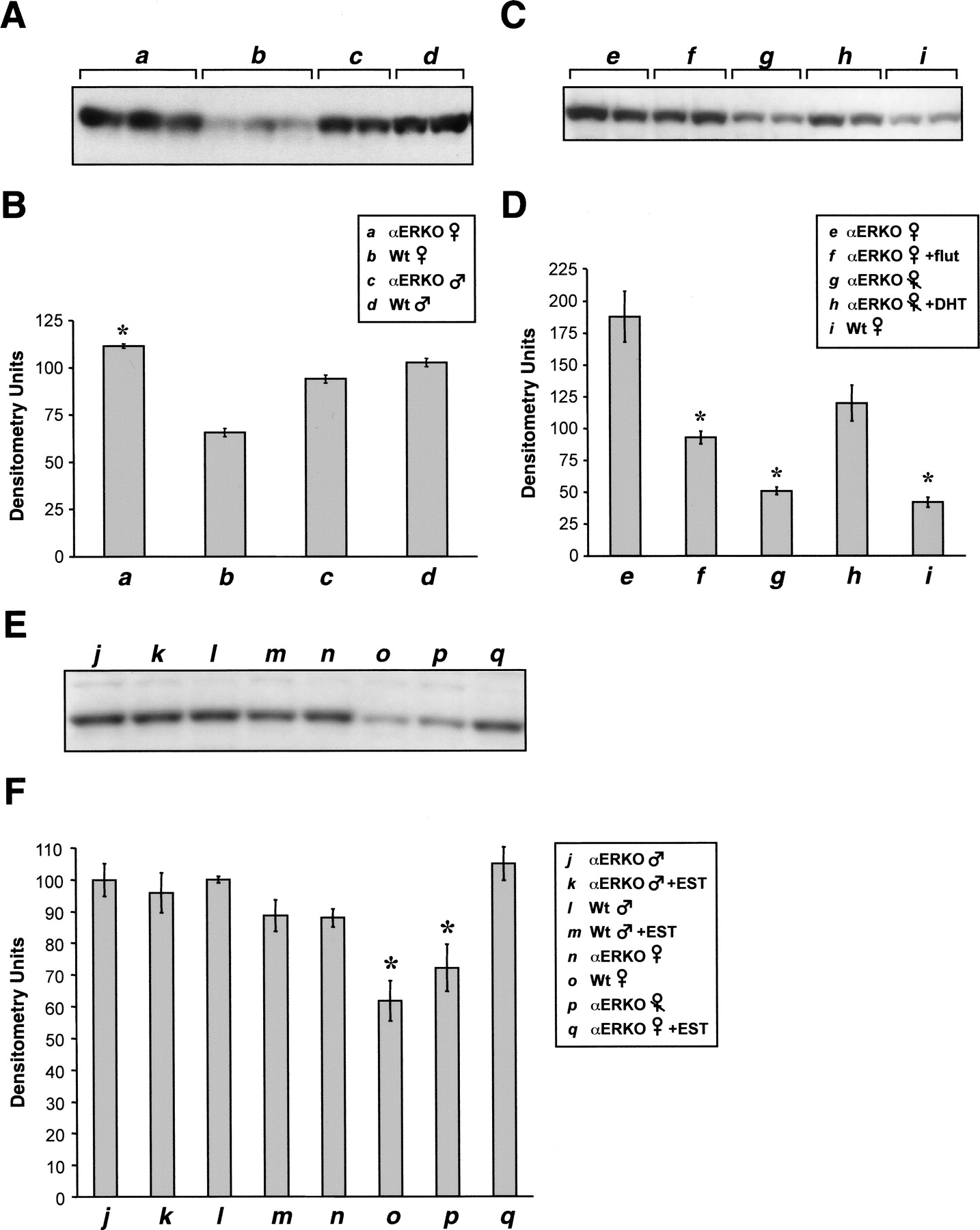
To examine the role of the estrogen receptor-α in this process, we examined CYP2J5 expression in male and female αERKO mice. There was no significant difference in renal CYP2J5 expression between αERKO and wild-type male mice (Fig. 10, A and B). In contrast, αERKO female mice have higher renal CYP2J5 expression than wild-type female mice. Indeed, CYP2J5 expression in αERKO female mice is comparable with that in wild-type male mice. These data suggest a role for the estrogen receptor-α in regulating renal CYP2J5 expression. However, the αERKO female mice are known to have elevated circulating testosterone levels (Eddy et al., 1996). Either ovariectomy of αERKO female mice or treatment of intact αERKO female mice with the androgen antagonist flutamide resulted in a significant reduction in renal CYP2J5 expression compared with intact, untreated αERKO female mice (Fig. 10, C and D). The effect of ovariectomy on renal CYP2J5 expression in αERKO female mice was reversed by treatment with DHT for 3 to 4 weeks. Moreover, renal CYP2J5 levels in ovariectomized or flutamide-treated αERKO female mice were similar to those in wild-type female mice, suggesting that the observed CYP2J5 expression phenotype of the female αERKO mice was caused largely by excess circulating androgens. We also examined the effect of estrogen on CYP2J5 expression in intact wild-type and αERKO male and αERKO female mice. Interestingly, treatment with EST for 3 to 4 weeks had little effect on renal CYP2J5 levels in these intact animals (Fig. 10, E and F). This suggests that elevated circulating androgen levels can override the down-regulatory effects of estrogen in intact mice.

Expression of renal CYP2J5 in homozygous estrogen receptor-α knockout mice and wild type controls. A, microsomal fractions (30 μg of protein/lane) prepared from kidneys (n = 3 animals per group) of intact αERKO female mice (a), intact wild-type female mice (b), intact αERKO male mice (c), and intact wild-type male mice (d) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of three independent determinations. B, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals with the value for each animal taken as the average of two or three determinations. *, p < 0.05 versus wild type of same sex. C, microsomal fractions (30 μg of protein/lane) prepared from kidneys (n = 3 animals per group) of intact αERKO female mice (e), intact αERKO female mice treated with flutamide pellets for 3 to 4 weeks (f), αERKO female mice ovariectomized at week 8 and treated with placebo pellets for 3 to 4 weeks (g), αERKO female mice ovariectomized at week 8 and treated with DHT pellets for 3 to 4 weeks (h), and intact wild-type female mice (i) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of five independent experiments. D, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group with the value for each animal taken as the average of three to five determinations. *, p < 0.05 versus intact αERKO female. E, microsomal fractions (30 μg of protein/lane) prepared from kidneys (n = 3 animals per group) of intact αERKO male mice treated with placebo pellets for 3 to 4 weeks (j), intact αERKO male mice treated with EST pellets for 3 to 4 weeks (k), intact wild-type male mice treated with placebo pellets for 3 to 4 weeks (l), intact wild-type male mice treated with EST pellets for 3 to 4 weeks (m), intact αERKO female mice treated with placebo pellets for 3 to 4 weeks (n), intact wild-type female mice treated with placebo pellets for 3 to 4 weeks (o), αERKO female mice ovariectomized at week 8 and treated with placebo pellets for 3 to 4 weeks (p), and intact αERKO female mice treated with EST pellets for 3 to 4 weeks (q) were electrophoresed, transferred to nitrocellulose, and immunoblotted with the anti-CYP2J5pep IgG as described. The immunoblot shown is representative of six independent experiments. F, autoradiographs were scanned and relative CYP2J5 protein levels were determined. Values shown are the means ± S.E. of n = 3 animals per group with the value for each animal taken as the average of six determinations. *, p < 0.05 versus intact αERKO female.
Discussion
Cytochromes P450 catalyze the oxidative metabolism of a wide variety of xenobiotic substrates, including pharmaceuticals, industrial chemicals, environmental pollutants, and chemical carcinogens (Nelson et al., 1996). In the kidney, sexual dimorphism in the expression of several of these P450s has been proposed to contribute to gender-related differences in susceptibility to toxic and/or carcinogenic effects of these xenochemicals (Hong et al., 1989; Henderson et al., 1990; Henderson and Wolf, 1991; Hoivik et al., 1995; Heng et al., 1997). Kidney P450s are also involved in the metabolism of endogenous substrates such as arachidonic acid to bioactive eicosanoids that affect renal fluid-electrolyte transport and vascular tone and therefore contribute significantly to renal function and the regulation of blood pressure (Zeldin, 2001; Roman, 2002). Indeed, existing data suggest a role for the renal P450 arachidonic acid metabolic cascade in the pathogenesis of systemic hypertension (Holla et al., 2001; Zeldin, 2001; Roman, 2002), a disease in which gender differences in incidence and severity are known to occur in rodent models and in humans (Chen, 1996; Holla et al., 2001). Herein, we report that CYP2J5, a recently described mouse P450 that is particularly abundant in the kidney and active in the metabolism of arachidonic acid to EETs (Ma et al., 1999), is present at higher levels in male than in female kidney after puberty. Correspondingly, adult mouse renal arachidonic acid epoxygenase activity is also higher in male mice versus female mice. We also demonstrate that renal CYP2J5 levels are regulated at the pretranslational level by both androgens and estrogens, that this regulation does not depend on growth hormone, and that the effects of androgen are mediated by the androgen receptor.
A great deal is known regarding regulation of hepatic P450s. Xenochemicals such as β-naphthoflavone, phenobarbital, and clofibrate are known to induce liver CYP1A, CYP2B, CYP4A subfamily P450s, respectively (Capdevila et al., 1990; Wu et al., 1997; Gilday et al., 1998; Schlezinger et al., 1998). Other factors, including nutritional status, developmental factors, inflammatory stimuli, nitric oxide, hormonal status, and certain disease states, such as diabetes, are also known to influence hepatic P450 expression and monooxygenase activities (Thummel and Schenkman, 1990; Nelson et al., 1996; Qu et al., 1998; Ma et al., 1999). In particular, a number of studies have documented a role for sex hormones in regulating the expression of liver P450s (Morgan et al., 1985; Waxman et al., 1985; McClellan-Green et al., 1989; Thummel and Schenkman, 1990; Legraverend et al., 1992). These effects seem to be regulated at the mRNA level, at least in part by growth hormone, and probably involve activation of STAT5b (McClellan-Green et al., 1989; Legraverend et al., 1992; Mode et al., 1992; Udy et al., 1997; Park et al., 1999). In the present work, we show that liver CYP2J5 expression is regulated during development but that expression levels are comparable in male and female mice at all ages. Thus, in both sexes, hepatic CYP2J5 levels are low or absent during the fetal, newborn, and early postnatal periods, gradually increase at 1 to 2 weeks of age, and achieve maximal levels by 4 weeks of age. Similarly, adult liver CYP2J5 mRNA content is comparable in male and female mice. In agreement with this data, we found that hepatic microsomal arachidonic acid epoxygenase activity was also similar in adult male and female mice, although multiple P450s probably contribute to this activity (Capdevila et al., 1990).
In contrast to the liver, little is known about regulation of kidney P450s. Capdevila and coworkers have reported that dietary salt induces rat CYP2C23, leading to increased renal EET biosynthesis (Holla et al., 1999). Oyekan et al. (1999) showed that inhibition of nitric oxide synthase with Nω-nitro-l-arginine-methyl ester significantly increased rat renal CYP4A expression and conversion of arachidonic acid to 20-hydroxyeicosatetraenoic acid. Sexual dimorphism has been reported in the expression of a number of kidney P450s, including CYP2A, CYP2B, CYP2C, CYP2E, CYP3A, CYP4A, and CYP4B subfamily members (Henderson et al., 1990; Henderson and Wolf, 1991; Heng et al., 1997; Holla et al., 2001). Although the molecular mechanisms responsible for these effects remain unknown, they seem to be mediated, at least in part, by testosterone, which has the capacity to either induce or repress P450 gene expression in an isoform-specific fashion (Henderson et al., 1990; Henderson and Wolf, 1991). We found that renal CYP2J5 protein expression increased gradually during postnatal development in both male and female mice up to approximately 4 weeks of age, after which female kidney CYP2J5 levels decreased whereas male kidney CYP2J5 levels remained elevated. Consequently, CYP2J5 is expressed at higher levels in adult male versus adult female kidney. The gender differences in expression of renal CYP2J5 are mirrored by changes in the corresponding mRNAs (which are also higher in adult male mice than in female mice), suggesting that the mechanism responsible for the sexually dimorphic pattern of CYP2J5 expression involves regulation at the pretranslational level. Henderson and coworkers observed that, except for the mouse CYP4As, gender differences in renal P450 protein levels were accompanied by corresponding changes in renal P450 mRNA content and suggested a mechanism that involved transcriptional activation of these genes (Henderson et al., 1990; Henderson and Wolf, 1991). Sundseth and Waxman (1992) showed that male-specific rat renal CYP4As exhibit a correspondence between mRNA and protein. In contrast, Pan et al. (1992) showed that sex differences in renal CYP2E1 expression were regulated at the post-transcriptional level.
To further evaluate the mechanisms involved in regulating renal CYP2J5 levels, we examined the effects of androgens and estrogens. Castration of male mice results in a significant decrease in renal CYP2J5 protein expression, and treatment of castrated male or female mice with DHT increases expression to levels that approximate those in intact male mice. Moreover, mice that lack a functional androgen receptor (Tfm hemizygous) have reduced levels of renal CYP2J5 protein and do not respond to DHT treatment. Similarly, wild-type male mice treated with flutamide, an androgen receptor antagonist, exhibit reduced renal CYP2J5 protein levels. Together, these data support a role for androgens in up-regulating renal expression of CYP2J5 and implicate the androgen receptor in mediating these effects. Interestingly, neonatal administration of testosterone propionate to castrated male mice or ovariectomized female mice results in increased renal CYP2J5 expression during adulthood, suggesting that neonatal androgens can program or “imprint” a masculinizing effect on renal CYP2J5 expression, as has been shown to occur with liver P450s involved in steroid metabolism (Wong et al., 1987; Waxman and Chang, 1995). We also examined the effects of estrogens on renal CYP2J5 expression. We found that treatment of ovariectomized female mice or castrated male mice with 17β-estradiol causes a reduction in CYP2J5 protein expression. Moreover, female estrogen receptor-α knockout (αERKO) mice have significantly increased renal CYP2J5 expression compared with wild-type female mice. However, renal CYP2J5 levels in ovariectomized or flutamide-treated αERKO female mice were similar to those found in wild-type female mice, suggesting that excess circulating androgens were primarily responsible for the observed CYP2J5 expression phenotype of the female αERKO mice. In addition, estrogen had no effect on CYP2J5 expression in intact male mice or αERKO female mice. Together, these data suggest a role for estrogens in down-regulating renal CYP2J5 expression only in the absence of elevated circulating androgen levels.
Sexual dimorphism of hepatic P450s has been shown to be regulated by growth hormone (McClellan-Green et al., 1989; Legraverend et al., 1992; Mode et al., 1992; Park et al., 1999). Like the liver, the kidney is considered a growth hormone-responsive tissue in that it expresses a growth hormone receptor gene (Mathews et al., 1989) and responds to growth hormone by synthesis of growth factors, such as insulin-like growth factor I (Mathews et al., 1986). To investigate the role of growth hormone in regulating renal CYP2J5 expression, we examined the expression of CYP2J5 in homozygous lit/lit mice which have markedly reduced circulating growth hormone levels. Interestingly, we found that lit/lit mice exhibit sexual dimorphism in renal CYP2J5 expression (i.e., have higher levels in male mice than in female mice). Moreover, castrated lit/lit mice have reduced renal CYP2J5 expression and treatment with DHT restores CYP2J5 levels. Taken together, these data indicate that the androgen effects on CYP2J5 expression are not mediated by alterations in the growth hormone secretory pattern. Henderson et al. (1990) have observed that expression patterns of other renal P450s in lit/lit mice are comparable with those in normal strains.
The following data suggest that the renal EET biosynthetic pathway may be important in the pathogenesis of hypertension: 1) the rat renal epoxygenases are under regulatory control by dietary salt and their inhibition leads to the development of salt-dependent hypertension (Holla et al., 1999; Oyekan et al., 1999); 2) spontaneously hypertensive rats have altered renal epoxygenase and epoxide hydrolase activities, and treatment of these animals with agents that either deplete renal P450 or inhibit epoxide hydrolase normalizes blood pressure (Sacerdoti et al., 1988; Yu et al., 2000a,b); 3) the salt-sensitive phenotype in the Dahl rat model of genetic hypertension is associated with an inability to increase renal epoxygenase activity in response to dietary salt intake (Makita et al., 1994); 4) targeted disruption of the soluble epoxide hydrolase gene lowers blood pressure (Sinal et al., 2000); and 5) the urinary excretion of epoxygenase metabolites is increased in women with pregnancy-induced hypertension (Catella et al., 1990). Moreover, recent data implicate the CYP2J subfamily, in particular, in the development of hypertension as follows: 1) renal CYP2J expression is localized to sites of the nephron in which EETs have been shown to mediate the actions of hormones that affect blood pressure (Ma et al., 1999); 2) spontaneously hypertensive rats exhibit altered renal CYP2J expression (Yu et al., 2000a); 3) the CYP2J cluster has been shown to map to a chromosomal region that cosegregates with the hypertensive phenotype in Dahl salt-sensitive rats (Stec et al., 1996; Ma et al., 1998); and 4) disruption of the Cyp2j5 gene causes spontaneous hypertension in mice (J. Ma, K. Athirakul, T. Coffman, and D. C. Zeldin, unpublished observations). Importantly, as is the case in human essential hypertension and spontaneous hypertension in rodents (Chen, 1996), the hypertension observed in Cyp2j5-null mice occurs in a sexually dimorphic fashion. CYP2J2, the human ortholog of mouse CYP2J5, has a broad tissue distribution and is known to be abundant in human kidney (Wu et al., 1996). Whether CYP2J2 expression in human kidney occurs in a sexually dimorphic fashion remains unknown.
In summary, we show that renal CYP2J5 expression is higher in male mice than in female mice after puberty. Correspondingly, adult mouse renal arachidonic acid epoxygenase activity is also higher in male mice. Mechanistic studies involving castration/ovariectomy with/without hormone replacement, androgen receptor-deficient (Tfm hemizygous) and estrogen receptor-deficient (αERKO) mice, and growth hormone-deficient (lit/lit) mice demonstrate that renal CYP2J5 levels are regulated at the pretranslational level by both androgens and estrogens but that this regulation does not depend on growth hormone. Given the data supporting a role for P450 epoxygenases in general and CYP2J5 in particular in the development of hypertension, we speculate that studies on the regulation of CYP2J5 by sex hormones may provide insight into the basic mechanisms involved in the sexual dimorphism of this disease.
Acknowledgments
We thank Drs. Joyce Goldstein and Masahiko Negishi for providing helpful comments during preparation of this manuscript. We also acknowledge the technical assistance of Tracy Demianeko.
Footnotes
-
This work was supported by the National Institute of Environmental Health Sciences/National Institutes of Health Division of Intramural Research.
-
ABBREVIATIONS: P450, cytochrome P450; EET, cis-epoxyeicosatrienoic acid; STAT, signal transducer and activator of transcription; αERKO, estrogen receptor-deficient mice; DHT, 5α-dihydrotestosterone; EST,17β-estradiol; RT-PCR, reverse transcription-polymerase chain reaction.
- Received April 24, 2003.
- Accepted December 12, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}