


Abstract
The ATP-binding cassette (ABC) transporter ABCG2 has been implicated to play a significant role in the response of patients to medication and/or the risk of diseases. To clarify the possible physiological or pathological relevance of ABCG2 polymorphisms, we have functionally validated single nucleotide polymorphisms (SNP) of ABCG2. In the present study, based on the currently available data on SNPs and acquired mutations, we have created a total of 18 variant forms of ABCG2 (V12M, G51C, Q126stop, Q141K, T153M, Q166E, I206L, F208S, S248P, E334stop, F431L, S441N, R482G, R482T, F489L, F571I, N590Y, and D620N) by site-directed mutagenesis and expressed them in insect cells. Because porphyrins are considered to be endogenous substrates for ABCG2, we have investigated the porphyrin transport activity of those variant forms in vitro. We herein provide evidence that the variants Q126stop, F208S, S248P, E334stop, and S441N are defective in porphyrin transport, whereas F489L exhibited impaired transport, approximately 10% of the activity observed for the wild type. Furthermore, Flp-In-293 cells expressing those variants were photosensitive. Thus, among those genetic polymorphisms of ABCG2, at least the hitherto validated alleles of Q126stop, S441N, and F489L are suggested to be of clinical importance related to the potential risk of porphyria.
ATP-binding cassette (ABC) transporters form one of the largest protein families encoded in the human genome, and more than 48 human ABC protein genes have been identified (Klein et al., 1999). It has been reported that mutations of certain ABC protein genes are causative for several genetic disorders in humans (Dean et al., 2001; Borst and Elferink, 2002; Gottesman et al., 2002; Ishikawa, 2003; Varadi et al., 2003).
Human ABCG2 is a member of the ABC transporter family (Allikmets et al., 1998; Doyle et al., 1998; Miyake et al., 1999). The ABCG2 gene is located on chromosome 4q22, spans more than 66 kilobases, and consists of 16 exons ranging from 60 to 532 base pairs (Bailey-Dell et al., 2001). ABCG2 extrudes xenobiotics and certain drugs from cells, thereby mediating drug resistance and affecting the pharmacological behavior of many compounds.
Furthermore, ABCG2 is expressed in a variety of stem cells, and its potential role in the regulation of hematopoietic development is suggested (Zhou et al., 2001). Expression of Abcg2, a murine ortholog of human ABCG2, is a conserved feature of murine stem cells from different sources, including the bone marrow, skeletal muscle, and embryonic stem cells (Zhou et al., 2001). Abcg2 mRNA is expressed at high levels in primitive murine hematopoietic stem cells and is sharply down-regulated with differentiation. Enforced expression of the human ABCG2 cDNA directly conferred the side population phenotype to bone marrow cells. Loss of the Abcg2 gene led to a reduction in the number of side population cells in the bone marrow and skeletal muscle (Zhou et al., 2002). Abcg2 null hematopoietic cells were significantly more sensitive than normal hematopoietic cells to mitoxantrone in drug-treated transplanted mice. Mice lacking Abcg2 displayed a previously unknown type of protoporphyria, where erythrocyte levels of protoporphyrin IX were increased 10-fold (Jonker et al., 2002). It has recently been demonstrated that under hypoxic conditions heme molecules and/or porphyrins are detrimental to Abcg2 (–/–) progenitor cells (Krishnamurthy et al., 2004). Abcg2 specifically binds heme, and cells lacking Abcg2 accumulate porphyrins. In addition, Abcg2 expression is up-regulated by hypoxia, where the hypoxia-inducible transcription factor complex HIF-1 is involved. These findings suggest that Abcg2 confers a strong survival advantage for stem cells under hypoxic conditions. Furthermore, erythroid cells engineered to express Abcg2 had significantly lower intracellular levels of protoporphyrin IX, suggesting that the protoporphyrin IX level is modulated by human ABCG2 (Zhou et al., 2005; Latunde-Dada et al., 2006).
Single nucleotide polymorphisms (SNPs) of ABC transporters have been reported to play a significant role in the patients' responses to medication and/or the risk of diseases (Kalow et al., 2001). Sequencing of the ABCG2 gene from human samples has revealed more than 80 different, naturally occurring sequence variations (Honjo et al., 2002; Iida et al., 2002; Imai et al., 2002; Backstrom et al., 2003; Itoda et al., 2003; Zamber et al., 2003; de Jong et al., 2004; Mizuarai et al., 2004; Bosch et al., 2005; Kobayashi et al., 2005). The analysis and validation of those SNPs occurring in the ABCG2 gene are of clinical importance.
To clarify the possible physiological or pathological relevance of the ABCG2 polymorphism, we have attempted to functionally characterize the variants. By using plasma membrane vesicles, we have recently developed a high-speed screening system to precisely evaluate functional changes associated with such genetic polymorphisms in vitro (Ishikawa et al., 2005a). Because porphyrins are considered to be endogenous substrates for ABCG2, we have investigated the transport of porphyrins with a total of 18 variant forms of human ABCG2 in the plasma membrane vesicle system. We herein provide evidence that the variants Q126stop, F208S, S248P, E334stop, S441N, and F489L are defective or impaired in the transport of porphyrins, suggesting that those genetic polymorphisms in the ABCG2 gene may be related to the risk of certain diseases resulting from disruption of porphyrin homeostasis.
Materials and Methods
Chemicals and Biological Reagents. The following compounds and therapeutic drugs were purchased from the commercial sources indicated in parentheses: ATP, hematoporphyrin, and protoporphyrin IX (Sigma-Aldrich, St. Louis, MO); pheophorbide a (Frontier Scientific, Inc., Logan, UT); methotrexate and l-glutamine (Wako Pure Chemicals, Osaka, Japan); creatine kinase, creatine phosphate, EGTA, EDTA, Tris, HEPES, and high-glucose Dulbecco's modified Eagle's medium (Nacalai Tesque, Kyoto, Japan); fetal calf serum (FCS) (Dainippon Pharmaceuticals, Osaka, Japan); and antibiotic-antimycotic cocktail solution and hygromycin B (Invitrogen, Carlsbad, CA). All other chemicals used were of analytical grade.
SNP Data on Nonsynonymous Polymorphisms of HumanABCG2Gene. SNP data on the polymorphisms of ABCG2 were obtained from the National Center for Biotechnology Information dbSNP database and recent publications (Honjo et al., 2002; Iida et al., 2002; Imai et al., 2002; Backstrom et al., 2003; Itoda et al., 2003; Zamber et al., 2003; de Jong et al., 2004; Mizuarai et al., 2004; Bosch et al., 2005; Kobayashi et al., 2005). Figure 1 illustrates the positions of the nonsynonymous polymorphisms of ABCG2 that were functionally analyzed in the present study.
Preparation of Plasmids Carrying ABCG2 Variant cDNA. Wild-type (WT) ABCG2 cDNA was inserted into the pFastBac1 plasmid as described previously (Ishikawa et al., 2003). Nonsynonymous SNP variants were generated by using the QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA). Polymerase chain reaction (PCR) was carried out in an iCycler (Bio-Rad, Hercules, CA) by using PfuTurbo DNA polymerase, the ABCG2-pFastBac1 plasmid, and specific primers. Table 1 summarizes primer sequences, %GC, and melting temperature (Tm). The PCR was initiated by incubation at 95°C for 30 s and then followed by 12 cycles of reactions at 95°C for 30 s, at Tm for 1 min, and at 68°C for 14 min. After the PCR, the reaction mixture was incubated with DpnI endonuclease at 37°C for 1 h to digest the original template plasmid. Each variant cDNA generated in the pFastBac1 plasmid was subjected to nucleotide sequence analysis (Hitachi, Tokyo, Japan).
PCR primers and conditions for site-directed mutagenesis to create variants of ABCG2
Sites of mutagenesis are indicated by underlining. GC indicates the percentage of guanine and cytosine contents in the PCR primer set. Tm shows the melting temperature (Tm) for each PCR primer set.
Expression of ABCG2 Variants in Sf9 Cells. Competent DH10Bac Escherichia coli cells were transformed by the variant ABCG2 plasmids. Then, the variant ABCG2 cDNA was transposed into a bacmid, which is a baculovirus shuttle vector carrying the baculovirus genome, in DH10Bac cells with the help of a helper plasmid. The recombinant bacmid was isolated and purified.
Insect Spodoptera frugiperda Sf9 cells were grown in EX-CELL 420 Insect serum-free medium (JRH Biosciences, Lenexa, KS) supplemented with 1% (v/v) heat-inactivated FCS, 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen) with gentle shaking at 27°C. Sf9 cells were then transfected with the ABCG2-recombinant bacmid in the presence of Cellfectin reagent (Invitrogen) according to the manufacturer's protocol. Ninety-six hours after the transfection, the culture medium containing the recombinant baculovirus was harvested by centrifugation. To amplify recombinant baculovirus, Sf9 cells were further infected with the harvested virus and maintained at 27°C for 72 h. After the incubation, the culture medium was harvested by centrifugation. This process was repeated two times.
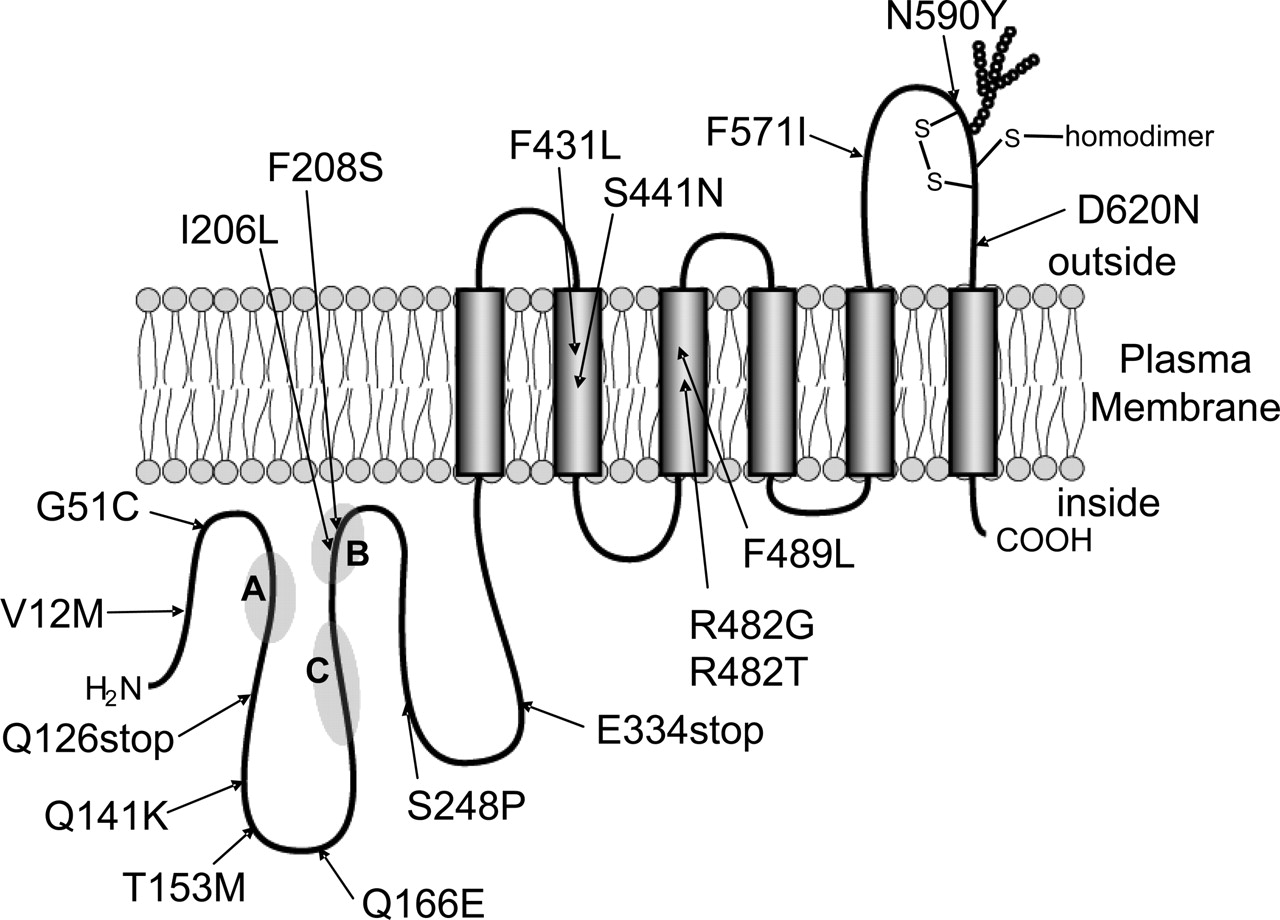
Schematic illustration of human ABCG2 and its nonsynonymous polymorphisms. SNP data on the polymorphisms of ABCG2 were obtained from the National Center for Biotechnology Information dbSNP database and recent publications (Honjo et al., 2002; Iida et al., 2002; Imai et al., 2002; Backstrom et al., 2003; Itoda et al., 2003; Zamber et al., 2003; de Jong et al., 2004; Mizuarai et al., 2004; Bosch et al., 2005; Kobayashi et al., 2005). The variants R482G and R482T are acquired mutations. A, B, and C indicate the motifs of Walker A (amino acids 80–86), Walker B (amino acids 205–210), and signature C (amino acids 186–200).
Sf9 cells (1 × 106 cells/ml) were infected with the amplified recombinant baculoviruses and cultured in EX-CELL 420 Insect serum-free medium at 27°C with gentle shaking. Three days after the infection, cells were harvested by centrifugation. Cells were subsequently washed with phosphate-buffered saline at 4°C, collected by centrifugation, and stored at –80°C until used.
Preparation of the Plasma Membrane Vesicles from Sf9 Cells. Plasma membrane vesicles were prepared from ABCG2-expressing Sf9 cells as described previously (Ishikawa et al., 2005a). The frozen cell pellet was thawed quickly, diluted 40-fold with a hypotonic buffer (0.5 mM Tris-HEPES, pH 7.4, and 0.1 mM EGTA), and then homogenized with a Potter-Elvehjem homogenizer. After centrifugation at 2000g, the supernatant was further centrifuged at 100,000g for 30 min. The resulting pellet was suspended in 0.25 M sucrose containing 10 mM Tris-HEPES, pH 7.4. The crude membrane fraction was layered over 40% (w/v) sucrose solution and centrifuged at 100,000g for 30 min. The turbid layer at the interface was collected, suspended in 0.25 M sucrose containing 10 mM Tris-HEPES, pH 7.4, and centrifuged at 100,000g for 30 min. The membrane fraction was collected and resuspended in a small volume (150–250 μl) of 0.25 M sucrose containing 10 mM Tris-HEPES, pH 7.4. After the protein concentration was measured by the BCA Protein Assay kit (Pierce Chemical, Rockford, IL), the membrane solution was stored at –80°C until used.
Immunological Detection of ABCG2 Expression in Plasma Membrane. Expression of ABCG2 in cell membranes was determined by immunoblotting with BXP-21 (Signet Laboratories, Dedham, MA), a specific antibody to human ABCG2. In brief, proteins of the isolated plasma membrane were separated by electrophoresis on 7.5% SDS polyacrylamide slab gels, and the proteins were electroblotted onto Hy-bond ECL nitrocellulose membranes (GE Healthcare, Little Chalfont, Buckinghamshire, UK). Immunoblotting was performed by using BXP-21 (1:500 dilution) as the first antibody and an anti-mouse IgG-horseradish peroxidase-conjugate (Cell Signaling Technology Inc., Beverly, MA) (1:3000 dilution) as the secondary antibody. Horseradish peroxidase-dependent luminescence was developed by using Western Lighting Chemiluminescent Reagent Plus (PerkinElmer Life and Analytical Sciences, Boston, MA) and detected by Lumino Imaging Analyzer FAS-1000 (Toyobo Engineering, Osaka, Japan).
Detection of ATP-Dependent Transport of Hematoporphyrin. The frozen stocked membrane was thawed quickly at 4°C. Plasma membrane vesicles (50 μg of protein) were incubated with 20 or 100 μM hematoporphyrin in the presence or absence of 1 mM ATP in 100 μl of the standard incubation medium (0.25 M sucrose and 10 mM Tris-HEPES, pH 7.4, 10 mM creatine phosphate, 100 μg/ml creatine kinase, and 10 mM MgCl2) at 37°C. After a specified incubation period, the reaction mixture was mixed with 220 μl of ice-cold stop solution (10 mM EDTA, 0.25 M sucrose, and 10 mM Tris-HEPES, pH 7.4); and then 100 μl of the resulting solution was loaded onto a Sephadex G-25 spin-column (volume of 1 ml) equilibrated with 0.25 M sucrose and 10 mM Tris-HEPES, pH 7.4. The column was immediately centrifuged in a swing-type rotor at 1600g for 5 min, whereby the eluate was collected in a microcentrifuge tube. A portion (50 μl) of the eluate was mixed with 950 μl of 10 mM NaOH solution to dissolve plasma membrane vesicles. Hematoporphyrin in the resulting solution was quantitatively analyzed by measuring its fluorescence in a Hitachi fluorescence spectrophotometer type F2000 (excitation at 400 nm; emission at 613 nm). The fluorescence spectrum of hematoporphyrin was recorded with a Horiba SPX Fluoro-Max-2 fluorescence spectrophotometer.
Detection of ATP-Dependent Transport of Methotrexate. The incubation medium contained plasma membrane vesicles (50 μg of protein), 200 μM[3′,5′,7′-3H]methotrexate (MTX; GE Healthcare), 0.25 M sucrose, 10 mM Tris-HEPES, pH 7.4, 10 mM MgCl2, 1 mM ATP, 10 mM creatine phosphate, and 100 μg/ml creatine kinase in a final volume of 100 μl. The reaction was started by adding [3H]methotrexate to the incubation medium. The reaction was carried out at 37°C, and the amount of [3H]methotrexate incorporated into the vesicles was measured by a rapid filtration technique as described previously (Ishikawa et al., 2005a).
Expression of ABCG2 and Its Variants in Flp-In-293 Cells. Flp-In-293 cells (Invitrogen) were maintained in high-glucose Dulbecco's modified Eagle's medium supplemented with 10% (v/v) heat-inactivated FCS, 2 mM l-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin in a humidified atmosphere of 5% CO2 in air. The number of viable cells was determined in a hemocytometer by trypan blue dye exclusion.
Flp-In-293 cells were transfected with the ABCG2-pcDNA5/FRT vector, the Flp recombinase expression plasmid pOG44, and LipofectAMINE 2000 (Invitrogen) as described previously (Wakabayashi et al., 2006). Single colonies resistant to hygromycin B (Invitrogen) were picked and subcultured. Selection of positive colonies was performed by immunoblotting, as described below. The resulting cells are described hereinafter as Flp-In-293/ABCG2 cells. Mock cells (Flp-In-293/Mock) were prepared by transfecting Flp-In-293 cells with pcDNA5/FRT and pOG44 vectors in the same manner as described above.
Photosensitivity Assay. The effect of pheophorbide a on the photosensitivity of mock or ABCG2-expressing Flp-In-293 cells was measured as described by Robey et al. (2005). In brief, cells were seeded in 96-well plates (15,000 cell/well) and cultured at 37°C for 24 h. Pheophorbide a was added to the culture medium at increasing concentrations and then incubation was continued in the dark for 4 h. Thereafter, the culture medium was replaced with fresh medium. Cells were then exposed to light for 90 min in the cell culture chamber, where 96-well plates were placed on a light viewer (model 5700; Hakuba Photo Industry, Co., Ltd., Aichi, Japan). Thereafter, cells were incubated in the dark for 24 h, and cell viability was measured by the MTT assay as described previously (Mitomo et al., 2003).
Profiling of Drug Resistance of ABCG2-Expressing Flp-In-293 Cells. A growth inhibition (IC50) assay was performed by seeding Flp-In-293 cells at a density of 2000 cells per well in 96-well plates containing the culture medium. After 24 h, a test drug (SN-38, mitoxantrone, doxorubicin, or daunorubicin) was added to the culture medium at different concentrations, and cells were further incubated with the drug in a humidified tissue culture chamber (37°C; 5% CO2) for 72 h. Surviving cells were detected by the MTT assay (Mitomo et al., 2003) with modifications. In brief, 10 μl of MTT solution (5 mg/ml) was added to the culture medium, and cells were incubated for 4 h at 37°C. Thereafter, 100 μl of 10% (w/v) SDS in phosphate-buffered saline was added to the culture medium, and the mixture was incubated at 37°C overnight. The absorbance of formazan, a metabolite of MTT, in the resulting solution was photometrically measured at a test wavelength of 570 nm and at a reference wavelength of 630 nm in a Multiskan JX system (Dainippon Pharmaceuticals Co., Osaka, Japan). IC50 values were calculated from dose-response curves (i.e., cell survival versus drug concentration) obtained in duplicated experiments.
Results
Detection of ATP-Dependent Porphyrin Transport in the Membrane Vesicle System. In the present study, we first established an assay method to accurately measure ABCG2-mediated transport of hematoporphyrin in vitro. For this purpose, the wild type of human ABCG2 was expressed in Sf9 insect cells, and plasma membrane vesicles were prepared as described previously. Hematoporphyrin was chosen as a model compound for porphyrins, because it is hydrophilic and its solubility in aqueous solutions is higher than those of protoporphyrin IX and pheophorbide a.
Membrane vesicles prepared from Sf9 cells were incubated with 100 μM hematoporphyrin in the presence or absence of ATP to measure ATP-dependent transport of hematoporphyrin. However, the rapid filtration method widely used for the transport assay was not applicable to this porphyrin transport assay. Because hematoporphyrin was bound to the filter membrane surface, it caused high background levels in the transport measurements. Therefore, we undertook an alternative method. That is to say, after the incubation, we separated the membrane vesicles from the incubation mixture by means of a spin column (1 ml of bed volume) packed with Sephadex G-25 (Fig. 2A). Eluate fractions from the column were treated with 10 mM NaOH to dissolve the plasma membrane, and then the fluorescence of hematoporphyrin was measured (Fig. 2A). As demonstrated in Fig. 2A, hematoporphyrin was transported into membrane vesicles in the presence of ATP. This hematoporphyrin transported into membrane vesicles seemed to be recovered in the fraction 1. In contrast, excess hematoporphyrin remaining in the incubation mixture was trapped in the Sephadex G-25 column, and it was not eluted out under the present experimental conditions. ABCG2 expressed in the plasma membrane was also detected in the fraction 1 by immunoblotting, whereas no trace amount of ABCG2 was detected in the following fractions (Fig. 2A).
Figure 2 shows the fluorescence spectra of hematoporphyrin at different concentrations. There was a linear correlation between the fluorescence intensity at 613 nm (excitation at 400 nm) and its concentration (data not shown). The fluorescence spectrum of the eluate (Fig. 2B, bottom) was the same as that of the hematoporphyrin standard. Thus, based on the fluorescence intensity at 613 nm in fraction 1, we could determine the amount of hematoporphyrin incorporated into plasma membrane vesicles. When the medium osmolarity was increased by enhancing sucrose concentrations up to 1 M, the amount of hematoporphyrin taken up by plasma membrane vesicles markedly decreased (data not shown). Furthermore, 1 mM ortho-vanadate inhibited the uptake almost completely. Thus, these observations reflect transmembrane movement of hematoporphyrin rather than binding at the membrane surface.
Kinetic Properties in ABCG2-Mediated Hematoporphyrin Transport.Figure 3, A and B, shows the time courses for hematoporphyrin transported into membrane vesicles in the presence or absence of ATP. Compared with the mock-infected cells (Fig. 3A), ATP-dependent hematoporphyrin was pronounced in plasma membrane vesicles prepared from ABCG2-expressing Sf9 cells (Fig. 3B). The results provide evidence that ABCG2 indeed transports hematoporphyrin in an ATP-dependent manner. Figure 3C depicts the effect of hematoporphyrin concentration on the rate of hematoporphyrin transport observed in the presence or absence of ATP. The ATP-dependent transport exhibited saturation kinetics with respect to hematoporphyrin concentration. Based on the Lineweaver-Burk plot (Fig. 3B, inset), the apparent Km value was estimated to be 17.8 μM for hematoporphyrin.

Detection of ATP-dependent hematoporphyrin transport into plasma membrane vesicles by using a spin-column. Detection of hematoporphyrin and ABCG2 in fractions eluted from the spin-column (A) and fluorescence spectra of hematoporphyrin (B). Control and ABCG2-expressing plasma membrane vesicles (50 μg of protein) were incubated with 100 μM hematoporphyrin in the presence or absence of 1 mM ATP in the standard incubation medium (0.25 M sucrose and 10 mM Tris-HEPES, pH 7.4, 10 mM creatine phosphate, 100 μg/ml creatine kinase, and 10 mM MgCl2) at 37°C for 20 min. The reaction mixture was mixed with the stop solution and then loaded onto the top of a Sephadex G-25 spin-column equilibrated with 0.25 M sucrose and 10 mM Tris-HEPES, pH 7.4. The column was immediately centrifuged in a swing-type rotor at 1600g for 5 min, whereby the eluate was collected in a microcentrifuge tube. A portion (50 μl) was taken from each eluate fraction and mixed with 950 μl of 10 mM NaOH solution. The fluorescence spectrum of eluate fraction 1 was observed with a Horiba SPX FluoroMax-2 fluorescence spectrophotometer. For immunoblotting, 10 μl of the eluate was subjected to SDS-polyacrylamide gel electrophoresis, and ABCG2 was detected as described under Materials and Methods.
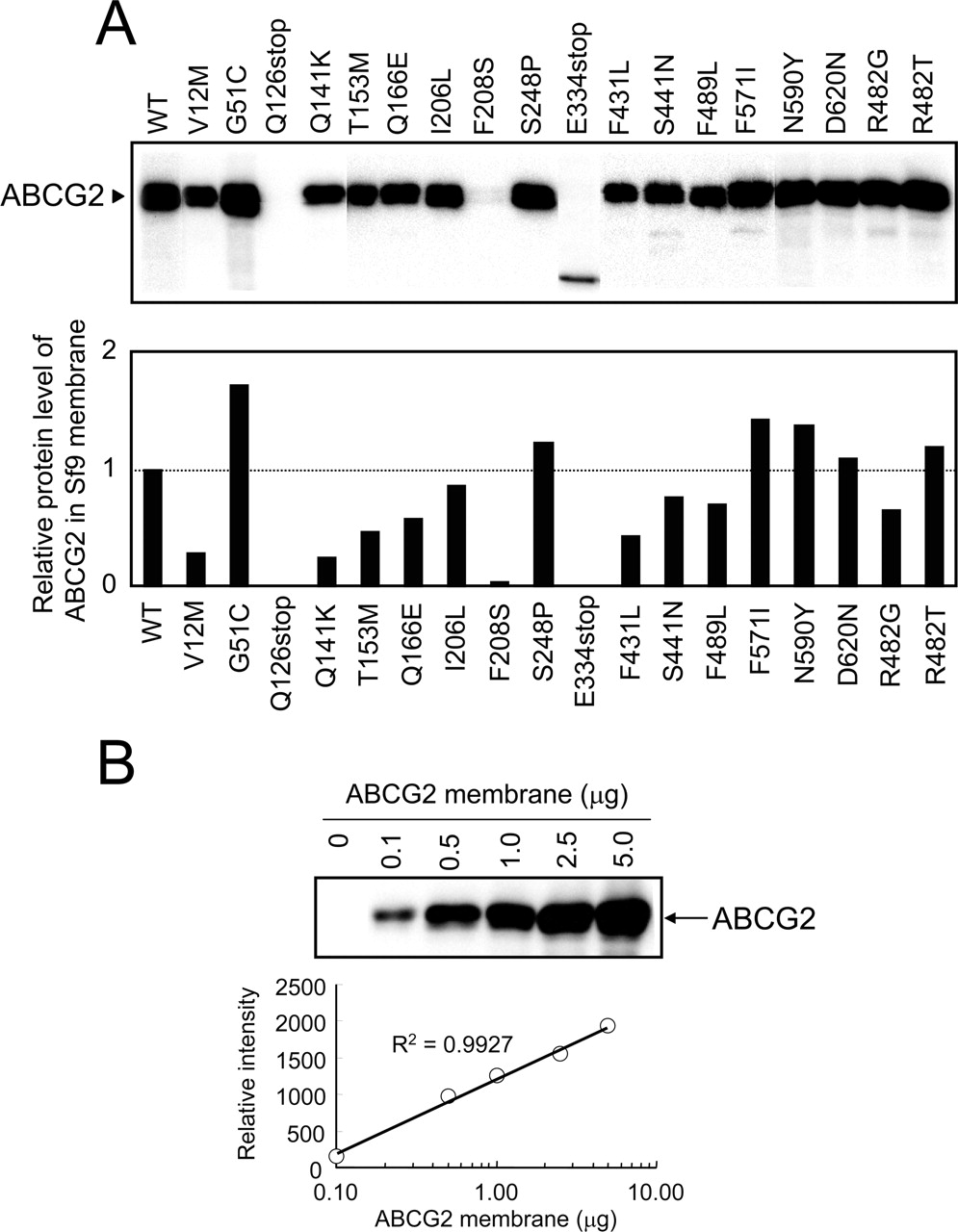
Human ABCG2 and Its Variants Expressed in Plasma Membrane of Sf9 Cells. As one of the specific aims in this study, we intended to examine the porphyrin transport activity of variant forms of ABCG2. For this purpose, based on the currently available data on SNPs and acquired mutations, we generated variant forms (i.e., V12M, G51C, Q126stop, Q141K, T153M, Q166E, I206L, F208S, S248P, E334stop, F431L, S441N, R482G, R482T, F489L, F571I, N590Y, and D620N) by site-directed mutagenesis. The variant forms as well as the WT of ABCG2 were then expressed in Sf9 cells by using the pFastBac1 vector and recombinant baculoviruses. Figure 4A shows the protein levels of those ABCG2 variants and the WT as detected by immunoblotting. Because those samples were treated with mercaptoethanol, ABCG2 was detected as a monomer with a molecular weight of 65,000. For Q126stop and E334stop, however, that treatment created truncated proteins (Fig. 4A). In addition, the expression level of F208S was found to be extremely low.
To quantitatively analyze the expression levels of ABCG2 variants, we analyzed the immunoblotting intensities versus protein levels. Figure 4B clearly demonstrates a linear relationship between the signal intensity of immunoblotting and the logarithmic value of the amount of protein applied to the electrophoresis. Based on the linear relationship, the expression levels of ABCG2 and its variants in different plasma membrane preparations were quantitatively estimated and normalized (Fig. 4A, bottom).
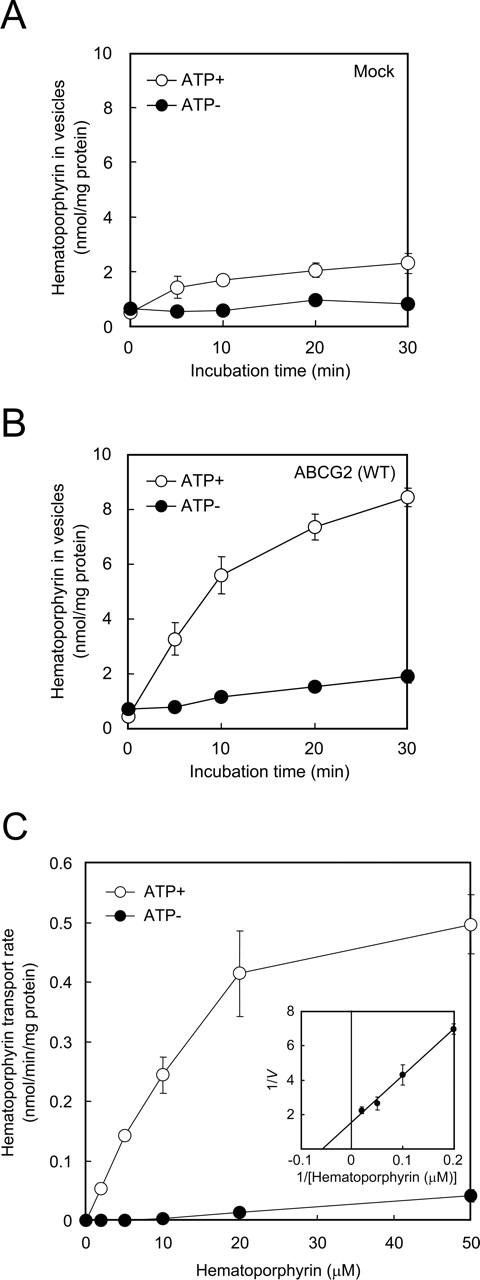
Time courses of ATP-dependent transport of hematoporphyrin into plasma membrane vesicles prepared from mock (A) and ABCG2-expressing Sf9 cells (B). Plasma membrane vesicles (50 μg of protein) were incubated with 100 μM hematoporphyrin in the presence or absence of 1 mM ATP in the standard incubation medium at 37°C for different periods as indicated. The amount of hematoporphyrin transported into membrane vesicles was detected as described under Materials and Methods. Effect of hematoporphyrin concentration on the rate of ATP-dependent transport of hematoporphyrin into plasma membrane vesicles (C) and Lineweaver-Burk plot (inset). ABCG2-expressing plasma membrane vesicles (50 μg of protein) were incubated with hematoporphyrin at different concentrations (0, 10, 20, 50, 100, and 200 μM) in the presence or absence of 1 mM ATP in the standard incubation medium at 37°C for 10 min. The amount of hematoporphyrin transported into membrane vesicles was spectrophotometrically detected.
Functional Validation of ABCG2 Variants: Transport of Hematoporphyrin and Methotrexate.Figure 5 demonstrates ATP-dependent transport of hematoporphyrin (A) and methotrexate (B) mediated by ABCG2 and its variants. Plasma membrane vesicles (50 μg of protein) expressing ABCG2 and its variants were incubated with 20 μM hematoporphyrin or 200 μM [3H]methotrexate in the presence of 1 mM ATP. Each transport activity was calculated by considering the normalized expression levels as described above (Fig. 4A, bottom). It is important to note that the variants Q126stop, F208S, S248P, E334stop, and S441N substantially lack transport activity for both hematoporphyrin and methotrexate. In contrast, the F489L variant, that did not transport methotrexate, exhibited impaired hematoporphyrin transport (Vmax = 0.058 nmol/min/mg of protein and Km = 8.6 μM for F489L versus Vmax = 0.654 nmol/min/mg of protein and Km = 17.8 μM for WT). The F431L variant as well as the acquired mutants R482G and R482T transported hematoporphyrin (Fig. 5, top), although they did not transport methotrexate (Fig. 5, bottom).
Involvement of ABCG2 Variants in Photosensitivity. To examine the potential link of nonsynonymous poymorphisms of ABCG2 with photosensitivity, ABCG2 WT as well as the variants F208S, S248P, S441N, F431L, and F489L were expressed in Flp-In 293 cells. When mock and ABCG2 WT-expressing cells were treated with pheophorbide a, the mock cells were more sensitive to light at low concentrations of pheophorbide a than were the ABCG2 WT-expressing cells (Fig. 6). Flp-In-293 cells expressing S441N were photosensitive (Fig. 6), suggesting that the S441N variant could not extrude pheophorbide a from cells, which lead them to become photosensitive. Similar results were observed with the variants of F208S and S248P (data not shown). Flp-In-293 cells expressing F489L were moderately sensitive to light (Fig. 6), which is consistent with the impaired activity of hematoporphyrin transport by this variant (Fig. 5). The F431L variant that was active in porphyrin transport conferred Flp-In-293 cells resistance to light as did the ABCG2 WT (Fig. 6).

Immunological detection of ABCG2 and its variant forms expressed in plasma membrane vesicles prepared from Sf9 cells (A) and relationship between the relative intensity of ABCG2 immunoreactivity and the amount of membrane proteins (B).
Drug Resistance Profiles of Flp-In-293 Cells Expressing ABCG2 WT and Variants. To gain more insight into the association of ABCG2 variants with cellular resistance to anticancer drugs, we incubated Flp-In-293 cells expressing ABCG2 WT, F431L, S441N, or F489L in the presence of SN-38, mitoxantrone, doxorubicin, or daunorubicin at different concentrations as described under Materials and Methods. Table 3 summarizes the drug resistance profile of those variants-expressing cells. Both Flp-In-293/ABCG2 (WT) and Flp-In-293/ABCG2 (F431L) cells were resistant toward SN-38 and mitoxantrone, whereas the resistance ratio of Flp-In-293/ABCG2 (S441N) and Flp-In-293/ABCG2 (F489L) cells were much lower, being close to that of Flp-In-293/Mock cells. None of the SNP variants of F431L, S441N, and F489L conferred Flp-In-293 cells resistance to doxorubicin or daunorubicin (Table 3), being different from the acquired mutants of R482G and R482T (Yoshikawa et al., 2004).
Drug resistance profiles of ABCG2 WT and variants
The drug resistance profiles of ABCG2 WT and variants were obtained by incubating Flp-In-293/ABCG2 WT, F431L, S441N, or F489L cells in the presence of SN-38, mitoxantrone, doxorubicin, or daunorubicin at different concentrations (0–100 μM) as described under Materials and Methods. IC50 values were calculated from dose-response curves (i.e., cell survival versus drug concentration). Drug resistance ratio = (IC50 value for cells expressing ABCG2 WT or variant)/(IC50 value for mock cells). Data are expressed as mean values ± S.D. (n = 3). Statistical analyses were performed by using Microsoft Excel 2003 software (Microsoft, Redmond, WA). The statistical significance was determined according to the two-sided Student's t test.
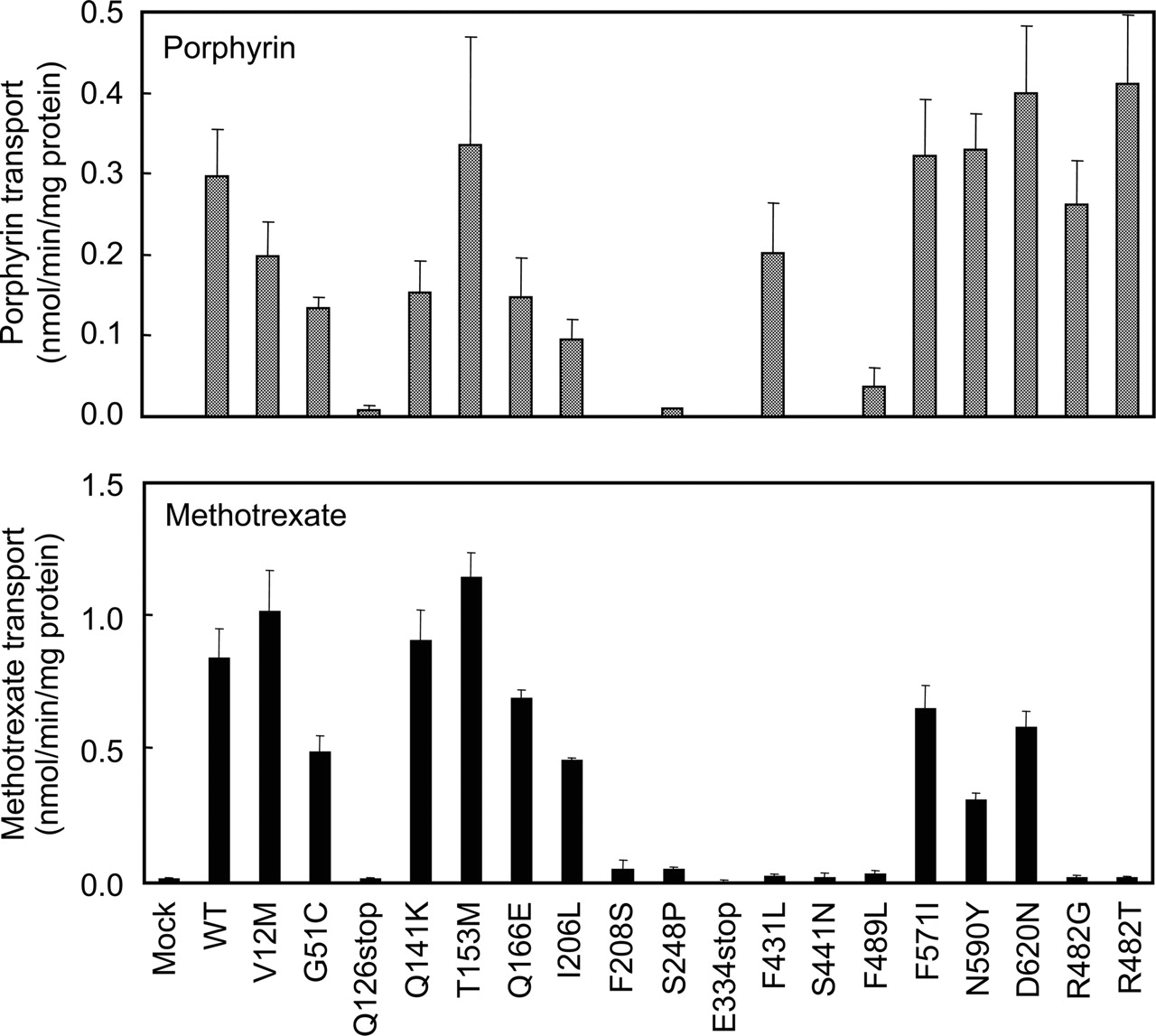
ATP-dependent transport of hematoporphyrin (A) and methotrexate (B) by ABCG2 and its variants. Plasma membrane vesicles (50 μg of protein) were incubated with 20 μM hematoporphyrin or 200 μM [3H]MTX in the presence or absence of 1 mM ATP in the standard incubation medium at 37°C for 10 min (hematoporphyrin) or 20 min (MTX). The ATP-dependent transport of hematoporphyrin or MTX is normalized for the amount of ABCG2 protein detected in Fig. 4.
Discussion
Transport of Porphyrins by ABCG2. In the present study, we have developed a simple method to quantitatively measure the ABCG2-mediated transport of porphyrins (Fig. 2). Our results show that the apparent Km value of ABCG2 WT toward hematoporphyrin was estimated to be 17.8 μM (Fig. 3C). We have recently performed quantitative structure-activity relationship analysis to study the substrate specificity of ABCG2 (Saito et al., 2006) and showed that human ABCG2 has high affinities toward protoporphyrin IX, hematoporphyrin, and pheophorbide a. This suggests that the tetrapyrrole structure is an important molecular feature in the interaction with human ABCG2.
The importance of human ABCG2 in the transport of protoporphyrin IX and pheophorbide a was implicated by Jonker et al. (2002) in experiments with Abcg2-knockout mice. These animals developed a skin phototoxicity that was associated with an accidental increase in dietary chlorophyll that produced an accumulation of pheophorbide a. Furthermore, erythrocyte levels of protoporphyrin IX were increased 10-fold in the Abcg2-knockout mice. Because protoporphyrin IX structurally resembles pheophorbide a, it is suggested that Abcg2 is involved in the excretion of these porphyrin compounds. Robey et al. (2005) have recently demonstrated that photosensitizers, such as pheophorbide a, chlorin e6, protoporphyrin IX, and hematoporphyrin, are substrates for ABCG2. Acquired mutations (R482G and R482T) at amino acid 482 did not affect the transport of those photosensitizers, being consistent with our findings (Fig. 5). Kato et al. (2003) have reported that some patients displayed extended photosensitivity after intravenous administration of the chlorin e6 derivative mono-l-aspartyl chlorin e6. Extended photosensitivity may be associated with naturally occurring polymorphisms of ABCG2.
In humans, pheophorbide a-induced phototoxicity has been reported after the ingestion of chlorella tablets (Jitsukawa et al., 1984), a dried algae preparation taken as a natural health supplement. Several pickled vegetables also contain substantial amounts of pheophorbide a and have been shown to cause phototoxicity in rats (Jitsukawa et al., 1984). In the human population, there are many incompletely understood incidences of idiosyncratic food and drug hypersensitivities, including phototoxicity (Bowers, 1999; Millard and Hawk, 2002; Millard et al., 2004). In this context, it would be of great interest to investigate whether part of these hypersensitivities is related to deficiencies in the function of human ABCG2.

Photosensitivity of Flp-In-293 cells expressing ABCG2 WT, F431L, S441N, or F489L. The effect of pheophorbide a on the photosensitivity of those cells was measured as described under Materials and Methods. Cells were incubated with pheophorbide a at different concentrations (0, 0.02, 0.04, 0.08, 0.16, 0.32, 0.64, 1.25, 2.5, 5, and 10 μM) for 4 h at 37°C. Thereafter, the culture medium was replaced with fresh medium. Cells were then exposed to light for 90 min in the cell culture chamber, where 96-well plates were placed on a light viewer (model 5700; Hakuba).
Genetic Polymorphisms of ABCG2 and Photosensitivity. Genetic polymorphic mutations of ABC transporters have been proposed to play a significant role in the patients' responses to medication and/or the risk of diseases (Kalow et al., 2001). To clarify the possible physiological or pathological relevance of ABCG2 polymorphisms, we have been functionally validating polymorphisms of ABCG2 (Ishikawa et al., 2005b). In the present study, based on the currently available data on SNPs and acquired mutations, we have created a total of 18 variant forms of ABCG2 (V12M, G51C, Q126stop, Q141K, T153M, Q166E, I206L, F208S, S248P, E334stop, F431L, S441N, R482G, R482T, F489L, F571I, N590Y, and D620N) by site-directed mutagenesis and expressed them in insect cells. We provide evidence that the variants Q126stop, F208S, S248P, E334stop, and S441N are defective in the transport of hematoporphyrin (Fig. 5). The F489L variant showed impaired transport activity (Fig. 5). Flp-In-293 cells expressing the F208S, S248P, S441N, and F489L variants were sensitive to light when cells were treated with pheophorbide a. Thus, it is likely that humans with these alleles may be more susceptible to porphyrin-induced phototoxicity.
Table 2 summarizes data on the present study and nonsynonymous polymorphisms hitherto reported. The frequencies of the Q126stop, S441N, and F489L alleles are relatively low (less than 2%) compared with those of the V12M and Q141K alleles. The variant Q126stop was consistently observed in certain Japanese cohorts; however, it was absent in two different Caucasian and African-American groups (Imai et al., 2002; Itoda et al., 2003; Mizuarai et al., 2004; Kobayashi et al., 2005; for a recent review, see Ishikawa et al., 2005) for recent review. Likewise, to date, the F489L allele was found only in the Japanese population (Itoda et al., 2003). In contrast, F208S, S248P and E334stop alleles are registered in the National Center for Biotechnology Information dbSNP database, but their allele frequencies are not available. The most recent version of National Center for Biotechnology Information dbSNP does not seem to have validation for Q166E, F208S, S248P, and E334stop (Table 2) as bona fide SNPs. Thus, the clinical significance of F208S, S248P and E334stop alleles in porphyrin-induced phototoxicity remains to be elucidated.
Porphyrin transport and non-synonymous polymorphisms of ABCG2
Transport of hematoporphyrin and MTX is indicated by either + (positive) or – (negative). Allele frequencies are summarized from previously reported SNP data on human ABCG2. There are differences in the allele frequency of the polymorphisms among different ethnic groups.
ABCG2 plays a physiologically important role in the handling of exogenous and endogenous porphyrins, and its role is even more emphasized in phototoxicity (Jonker et al., 2002). In addition, ABCG2 reportedly contributes to porphyrin homeostasis and regulates survival of stem cells under low oxygen conditions (Krishnamurthy et al., 2004). Therefore, at least, the validated alleles such as Q126stop, S441N, and F489L with a loss of porphyrin transport activity are at potential risk of diseases. It is assumed that their allele frequencies have been maintained at minimal levels throughout the history of Homo sapiens.
Concluding Remarks: Pharmacological Relevance. Because a large number of SNP data are presently available for a variety of drug transporters, it will become increasingly important to accurately determine which polymorphisms influence the substrate specificity of drug transporters. As exemplified by the S441N variant of ABCG2, amino acid substitution at the 441 site caused dramatic changes in the substrate specificity of ABCG2 (Fig. 5). To be precise, the S441N variant completely lost transport activity for both hematoporphyrin and methotrexate (Fig. 5). The F431L variant lacks the activity of methotrexate transport; however, it seems to be normal in terms of hematoporphyrin transport (Fig. 5). Although the F489L variant lacks the activity of methotrexate transport (Fig. 5), it maintains the activity of hematoporphyrin transport at the level of 10% compared with that of WT. As demonstrated in Table 3, the drug resistance profile of ABCG2 was greatly affected by these nonsynonymous polymorphisms. The amino acids at positions of 431, 441, and 489 located in transmembrane domains (Fig. 1) seem to be critically involved in substrate recognition and/or transport of drugs.
One should keep it in mind that the effect of SNPs on transport activity depends on the drugs tested; therefore, the functional analysis of SNPs using a variety of drugs is of great importance. In this context, we have recently developed a new method of QSAR analysis (Saito et al., 2006) to facilitate our understanding of the impact of SNPs on the pharmacological function of ABCG2. Relevant studies are ongoing in our laboratory (A. Tamura, K. Wakabayashi, Y. Onishi, M. Takeda, Y. Ikegami, S. Sawada, M. Tsuji, Y. Matsuda, and T. Ishikawa, manuscript in preparation).
Footnotes
-
This study was supported, in part, by the New Energy and Industrial Technology Development Organization International Joint Research grant program “International standardization of functional analysis technology for genetic polymorphisms of drug transporters” and grant 14370754 from the Japanese Society for the Promotion of Science.
-
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
-
doi:10.1124/mol.106.023556.
-
ABBREVIATIONS: ABC, ATP-binding cassette; SNP, single nucleotide polymorphism; FCS, fetal calf serum; WT, wild type; PCR, polymerase chain reaction; Tm, melting temperature; MTX, methotrexate; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium; SN-38, 7-ethyl-10-hydroxycamptothecin.
- Received February 15, 2006.
- Accepted April 11, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}