


Abstract
Tissue-specific expression of transporters is tightly linked with their physiological functions through the regulation of the membrane transport of their substrates. We hypothesized that epigenetic regulation underlies the tissue-specific expression of mouse liver-specific transporters (Oatp1b2/Slco1b2, Ntcp/Slc10a1, Bsep/Abcb11, and Abcg5/g8). We examined their DNA methylation and histone modification profiles near the transcriptional start site (TSS) in the liver, kidney, and cerebrum. Genome-wide DNA methylation profiling with tissue-dependent differentially methylated region profiling with restriction tag-mediated amplification and subsequent bisulfite genomic sequencing demonstrated that the CpG dinucleotides around the TSS of Oatp1b2 (from -515 to +149 CpGs), Ntcp (from -481 to +495 CpGs), Bsep (from -339 to +282 CpGs), and Abcg5/g8 (from -161 to +5 CpGs for Abcg5, i.e., from -213 to -48 CpGs for Abcg8) were hypomethylated in the liver and hypermethylated in the kidney and cerebrum. The opposite pattern was observed for Pept2/Slc15a2 (from -638 to +4 CpGs), which was expressed in the kidney and cerebrum but not in the liver. These DNA methylation profiles are consistent with the tissue distribution of these transporters. A chromatin immunoprecipitation assay demonstrated that the histone H3 associated with Oatp1b2, Ntcp, Bsep, and Abcg5/g8 promoters was hyperacetylated in the liver but was acetylated very little in the kidney and cerebrum, whereas the upstream region of Pept2 was hyperacetylated only in the kidney and cerebrum. These results suggest the involvement of epigenetic systems in the tissue-specific expression of mouse transporters Oatp1b2, Ntcp, Bsep, Abcg5/g8, and Pept2.
During the past two decades, extensive research has sought to uncover the molecular characteristics of transporters to explain their physiological and pathological functions. These transporters comprise more than 300 genes that are now classified into the solute carrier (SLC) and ATP-binding cassette (ABC) transporter superfamilies. Transporters show tissue-specific expression patterns that have critical implications for the physiological function of certain tissues (Borst and Elferink, 2002; Kusuhara and Sugiyama, 2002; Hediger et al., 2004; Stefkova et al., 2004; Shitara et al., 2006).
Liver tissue is one of the most important tissues in terms of the physiological turnover of endogenous compounds, such as bile acids and sterols, and the clearance of exogenous substances, including clinically used drugs. Several transporters are closely related to the hepatobiliary transport of these compounds. SLC-type transporters mediate the hepatic uptake of bile acids and xenobiotic organic anions, and ABC transporters then contribute to the biliary efflux of bile acids, sterols, and xenobiotic compounds and their metabolites (Faber et al., 2003; Chandra and Brouwer, 2004). Numerous reports suggest the involvement of liver-enriched transcription factors, such as hepatocyte nuclear factors (HNFs) 1, 3, and 4, and proteins of the CCAAT/enhancer-binding protein family, in the transcriptional regulation of liver-specific genes (Cereghini, 1996). However, these transcription factors are not expressed exclusively in the liver but are also observed in other tissues, implying that other machineries cooperate with the transcription factor network to establish and maintain the liver-specific gene expression pattern.
The role of epigenetic systems comprising DNA methylation and histone modification has attracted much interest as a possible mechanism underlying the tissue-specific gene expression (Bird, 2002; Shiota, 2004; Ohgane et al., 2008). Methylation of CpG dinucleotides, the unique target of DNA methylation in vertebrates, can evoke the condensed structure of chromatin by recruiting chromatin-remodeling factors, such as methyl CpG-binding proteins and histone deacetylases, and can prevent the transactivation of neighboring genes by most of the transcription factors. Therefore, gene expression is inversely correlated with DNA methylation in the 5′-flanking region. Our research group and other groups have demonstrated the involvement of DNA methylation-mediated gene silencing in the kidney-specific expression of SLC transporters, including the human organic anion transporter 3/SLC22A8, mouse urate transporter 1/Slc22a12, and human organic cation transporter 2/SLC22A2 (Kikuchi et al., 2006, 2007; Aoki et al., 2008). The DNA methylation status affects the accessibility of HNF1α/β to the recognition sequence within the promoter and confers the tissue-specificity of organic anion transporter 3 and urate transporter 1, whereas organic cation transporter 2 requires upstream stimulating factor 1 as a downstream transcription factor. The role of epigenetic regulation in the expression of liver-specific transporters has not been investigated.
We recently established a novel method, tissue-dependent differentially methylated region (T-DMR) profiling with restriction tag-mediated amplification (D-REAM) (Yagi et al., 2008), for analyzing the DNA methylation profiles of gene promoters that include HpyCH4IV restriction sites on a genome-wide scale. Plurality of the T-DMRs identified in DREAM is associated with the tissue-specific expression of the neighboring genes in the liver. The aim of the present study was to explore systematically the contribution of epigenetic regulation to the liver-specific expression of transporters. We used D-REAM to investigate the DNA methylation status in the 5′-flanking region of almost all mouse SLC and ABC transporter genes in the liver, kidney, and cerebrum and found that the liver-specific transporters, organic anion transporting polypeptide 1b2 (Oatp1b2)/Slco1b2, Na+-taurocholate-cotransporting polypeptide (Ntcp)/Slc10a1, bile salt export pump (Bsep)/Abcb11, Abcg5, and Abcg8, possess T-DMRs hypomethylated in the liver. Oatp1b2 and Ntcp mediate the sodium-independent and -dependent uptake of amphipathic organic anions and bile acids, respectively (Wolkoff and Cohen, 2003; Evers and Chu, 2008). Bsep is an ABC transporter that plays a pivotal role in the canalicular efflux of bile acids (Trauner and Boyer, 2003). Abcg5 and Abcg8 form a heterodimer to function as an efflux transporter for sterols into the bile (Hazard and Patel, 2007; Kidambi and Patel, 2008). The genes for BSEP and ABCG5/G8 are those responsible for the hereditary disorders progressive familial intrahepatic cholestasis type 2 (Strautnieks et al., 1998) and sitosterolemia (Berge et al., 2000; Lee et al., 2001; Lu et al., 2001), respectively. Bisulfite genomic sequencing and a chromatin immunoprecipitation (ChIP) assay were used to confirm and examine more extensively the DNA methylation and histone H3 acetylation profiles, respectively. Here we provide clear evidence that the liver-specific expression of mouse Oatp1b2, Ntcp, Bsep, Abcg5, and Abcg8 is associated with DNA methylation and histone acetylation in their promoters.
Materials and Methods
Materials. All reagents were purchased from Wako Pure Chemicals (Osaka, Japan) unless stated otherwise.
D-REAM. D-REAM and subsequent bioinformatics analysis were carried out as described previously for genomic DNA from the liver, kidney, and cerebrum of C57BL/6NCrj male mice at 12 or 13 weeks of age (Yagi et al., 2008). In brief, the genomic DNA was digested with a methylation-sensitive restriction enzyme HpyCH4IV that recognizes ACGT residues followed by subsequent amplification by modified ligation-mediated polymerase chain reaction (PCR). Because only the unmethylated DNA is digested by HpyCH4IV, the modified ligation-mediated PCR facilitates the selective amplification of the unmethylated region in the genomic DNA. The amplified fragments were then hybridized with mouse promoter tiling array, and the array signal intensities were analyzed to identify regions corresponding to fragments in unmethylated HpyCH4IV sites. Comparison of signals from different samples enabled the identification of differentially methylated regions. Genomic annotations, including Ensembl gene assignments, were performed on the Galaxy website (available at http://main.g2.bx.psu.edu/). The analyzed data were then visualized with the Integrated Genome Browser (available at http://www.affymetrix.com/partners_programs/programs/developer/tools/download_igb.affx). Tags of T-DMR (T-DMRtags) in the liver, kidney, and cerebrum were defined as the HpyCH4IV sites that are hypomethylated in the liver, kidney, and cerebrum compared with other tissues, respectively.
Genomic DNA Extraction and Bisulfite Reaction. Genomic DNA was extracted from the liver, kidney, and cerebrum of 11-week-old male C57BL/6NCrj mice using a Get Pure DNA Kit (Dojindo Molecular Technologies, Gaithersburg, MD) according to the manufacturer's instructions. Five to 10 μg of genomic DNA was digested with EcoRI and subjected to the bisulfite reaction as described previously (Kikuchi et al., 2007). The DNA was then precipitated with ethanol, dried, and resuspended in buffer containing 10 mM Tris-HCl and 1 mM EDTA, pH 8.0.
Combined Bisulfite Restriction Analysis. The DNA fragments encompassing the T-DMRtags of Oatp1b2/Slco1b2, Ntcp/Slc10a1, Bsep/Abcb11, Abcg5, and Pept2/Slc15a2 were amplified by PCR using the following primers: Oatp1b2 -594 (forward) to -347 (reverse); Ntcp +179 (forward) to + 671 (reverse); Bsep -110 (forward) to +416 (reverse); Abcg5 +1035 (forward) to +1212 (reverse); and Pept2 -171 (forward) to +155 (reverse) (Table 1). PCR was performed using Immolace DNA Polymerase (Bioline, London, UK) under the following conditions: 94°C for 10 min; 43 cycles of 94°C for 30 s, 53°C (Oatp1b2 and Pept2) or 55°C (Ntcp, Bsep, and Abcg5) for 30 s, and 72°C for 1 min; and final extension of 72°C for 10 min. The PCR fragments were digested with HpyCH4IV (New England Biolabs, Ipswich, MA) to confirm the methylation status of the T-DMRtags. Because only unmethylated cytosine residues are changed to thymines by the bisulfite reaction, PCR fragments from unmethylated genomic DNA are resistant to HpyCH4IV digestion, whereas those from methylated DNA are digested. The resultant products were electrophoresed on a 2.0% agarose gel and stained with ethidium bromide.
Oligonucleotides used for the combined bisulfite restriction analysis, bisulfite PCR, RT-PCR, and real-time PCR in the ChIP assay
Positions of primers for the RT-PCR are based on the published cDNA sequences, whereas those for the other experiments are relative to the TSS in the genome.
Bisulfite Genomic Sequencing. The DNA fragments covering the genomic region around the TSS of SLC or ABC transporters were amplified by PCR using the following primers: Oatp1b2, -594 (forward) to -347 (reverse) or -40 (forward) to +208 (reverse); Ntcp, -542 (forward) to -1 (reverse) or -46 (forward) to +522 (reverse); Bsep, -404 (forward) to -90 (reverse) or -110 (forward) to +340 (reverse); Abcg5, -315 (forward) to +125 (reverse); and Pept2, -669 (forward) to -144 (reverse) or -171 (forward) to +155 (reverse) (Table 1). The PCR was performed under the following conditions: 94°C for 10 min; 43 cycles of 94°C for 30 s, 53°C [Oatp1b2, -594 (forward)/-347 (reverse) and Pept2, -171 (forward)/+155 (reverse)], 55°C [Oatp1b2, -40 (forward)/+208 (reverse); Ntcp, -46 (forward)/+522 (reverse); Bsep, -404 (forward)/-90 (reverse); Bsep, -110 (forward)/+340 (reverse); and Abcg5, -315 (forward)/+125 (reverse)], or 57°C [Ntcp, -542 (forward)/-1 (reverse) and Pept2, -669 (forward)/-144 (reverse)] for 30 s, and 72°C for 1 min; and final extension of 72°C for 10 min. The PCR products were cloned into the pGEM-T Easy vector (Promega, Madison, WI), and 10 clones chosen randomly from each sample were sequenced to determine the presence of methylated cytosines.
RNA Isolation and RT-PCR. Total RNA was isolated from the liver, kidney, and cerebrum of a 14-week-old male C57BL/6NCrj mouse using ISOGEN (Nippon Gene, Toyama, Japan) according to the manufacturer's instructions. The RNA was then treated with DNase I to remove the contaminated genomic DNA, followed by reverse transcription using a random nonamer primer (Takara, Shiga, Japan). PCR was performed with the forward and reverse primers listed in Table 1 to detect the partial fragments of Oatp1b2, Ntcp, Bsep, Abcg5, Abcg8, Pept2, and β-actin cDNA. PCR was performed under the following conditions: 94°C for 2 min; 30 cycles (Oatp1b2, Bsep, and Abcg8), 32 cycles (Ntcp, Abcg5, and Pept2), or 35 cycles (β-actin) of 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min; and final extension of 72°C for 5 min. The resultant products were electrophoresed on a 2.0% agarose gel and stained with ethidium bromide.
ChIP Assay. The ChIP assay was performed using antiacetylated histone H3 antibody (Millipore, Billerica, MA) according to a procedure described previously (Fujii et al., 2006) and the Farnham Laboratory's protocol (available at http://www.genomecenter.ucdavis.edu/farnham/protocols/tissues.html) with minor modifications. Normal rabbit IgG was used as a negative control to verify the immunoprecipitation specificity. In brief, approximately 250 mg of tissue was chopped into small pieces and treated with 1% formaldehyde to form DNA-protein cross-links. The samples were homogenized, treated with cell lysis buffer, and centrifuged to precipitate the nuclei. The nuclei were then lysed with nuclei lysis buffer and sonicated on ice. The samples were preincubated with salmon sperm DNA/protein A-agarose suspension to reduce the nonspecific background, and the samples were incubated with an antibody against acetylated histone H3, normal rabbit IgG, or an equivalent amount of phosphate-buffered saline as the “no antibody” sample at 4°C overnight. The samples were then mixed with salmon sperm DNA/protein A-agarose suspension, rotated for 60 min at 4°C, and centrifuged at 20,000g for 1 min at 4°C. DNA was purified from the precipitated immune complex after extensive washing and reversing of the DNA-protein cross-links. The supernatant from the “no antibody” sample was collected as the “total input”. The quantitative real-time PCR was performed using a LightCycler and the appropriate software (version 3; Roche Diagnostics, Indianapolis, IN) according to the manufacturer's instructions with the primers shown in Table 1. The protocol for PCR was as follows: 95°C for 30 s, and then 40 cycles of 95°C for 5 s, 58°C for 10 s, and 72°C for 15 s. The enrichment of DNA in the immunoprecipitates was calculated relative to the 1:100 diluted “total input” samples.
Results
Screening of T-DMRtags by D-REAM. Screening with D-REAM revealed that 53 of 297 SLC transporter genes and 11 of 49 ABC transporter genes were associated with T-DMRtags that were hypomethylated in the liver and hypermethylated in the kidney or cerebrum in the genomic region from -6.0 to +2.5 kilobase pairs around the TSS (Supplemental Table 1). We focused on Oatp1b2/Slco1b2, Ntcp/Slc10a1, Bsep/Abcb11, Abcg5, and Abcg8, which are expressed predominantly in the liver, where they play indispensable roles in the hepatobiliary transport of endogenous and exogenous substances. D-REAM also located the T-DMRtag hypomethylated in the kidney and cerebrum whereas it was hypermethylated in the liver in the 5′-flanking region of Pept2, which is expressed predominantly in the kidney and cerebrum.
Combined Bisulfite Restriction Analysis of T-DMR Tags in SLC and ABC Transporters. Integrated Genome Browser plots of comparative D-REAM signals are shown around the TSS of Oatp1b2, Ntcp, Bsep, Abcg5, Abcg8, and Pept2 (Fig. 1A). These data suggest that the CpG dinucleotides at -515, +495, +224, +1141, and +1405 or +1431 base pairs relative to the TSS of Oatp1b2, Ntcp, Bsep, Abcg5, and Abcg8, respectively, are hypomethylated in the liver, whereas these sites are hypermethylated in the kidney and cerebrum. The CpG dinucleotide located at -41 base pairs relative to the TSS of Pept2 is hypomethylated in the kidney and cerebrum, whereas this site is hypermethylated in the liver. The DNA methylation status of these T-DMRtags found by D-REAM was verified in vivo in a combined bisulfite restriction analysis using HpyCH4IV as the restriction enzyme (Fig. 1B). These analyses corroborated that the T-DMRtags in Oatp1b2, Ntcp, Bsep, and Abcg5 genes are hypomethylated in the liver but hypermethylated in the kidney and cerebrum and vice versa for Pept2. The genomic region around the T-DMRtag in Abcg8 gene was not amplified after extensive PCR, and it was not possible to validate the D-REAM data with combined bisulfite restriction analysis (data not shown).
mRNA Expression Profiles of the SLC and ABC Transporters. The mRNA expression of Oatp1b2, Ntcp, Bsep, Abcg5, Abcg8, and Pept2 in the mouse liver, kidney, and cerebrum was investigated by RT-PCR (Fig. 2). The results confirmed the liver-specific expression of Oatp1b2, Ntcp, Bsep, Abcg5, and Abcg8. The expression of Pept2 was significant in both the kidney and cerebrum but was negligible in the liver.
DNA Methylation Profiles around the Transcriptional Start Site. The DNA methylation profiles in the CpG dinucleotides around the TSS of the above-mentioned transporters in mouse liver, kidney, and cerebrum were determined by bisulfite genomic sequencing (Fig. 3, A-E). The CpG dinucleotides around the TSS of Oatp1b2, Ntcp, and Bsep were overall significantly hypomethylated in the liver compared with the kidney and cerebrum (Fig. 3, A-C). The CpGs inside or adjacent to the intergenic region of Abcg5 and Abcg8 were also hypomethylated considerably in the liver but were hypermethylated in the kidney and cerebrum (Fig. 3D). The DNA methylation profiles are consistent with the liver-specific expression of the corresponding transporters. In contrast, the bisulfite sequencing for Pept2 demonstrated that the genomic region is hypomethylated in the kidney and cerebrum but hypermethylated in the liver, especially in the CpG dinucleotides located at -527, -54, -41, and +4 relative to the TSS, which is consistent with the predominant expression of this transporter in the kidney and cerebrum (Fig. 3E). These results suggest that the regions around the TSS of Oatp1b2, Ntcp, Bsep, and Abcg5/g8 can be regarded as T-DMRs and that the tissue-specific expression of these transporters is regulated through DNA methylation-mediated gene silencing.
Histone Acetylation Profiles in the Promoter Region. It is generally recognized that the DNA methylation status is linked tightly with the histone acetylation status and that activated genes are associated with acetylated histone H3. Thus, we performed ChIP assays to examine further the histone H3 acetylation profiles of the transporter genes with T-DMRs. In the liver, histone H3 in the proximal promoter of Oatp1b2, Ntcp, Bsep, and Abcg5/g8 was hyperacetylated, whereas the acetylation in the kidney and the cerebrum was minimal (Fig. 4). In contrast, acetylated histone H3 for the Pept2 upstream region was observed in the kidney and cerebrum but not in the liver. Histone H3 in the β-actin promoter was hyperacetylated in all tissues. These in vivo histone acetylation profiles were compatible with the tissue-specific expression of the transporter genes.

Comparative D-REAM and combined bisulfite restriction analysis. A, Integrated Genome Browser plots of comparative D-REAM signals. D-REAM and subsequent bioinformatics analysis were carried out and the analyzed data for Oatp1b2, Ntcp, Bsep, Abcg5/g8, and Pept2 were visualized with Integrated Genome Browser. Arrowheads and vertical bars represent T-DMRtags and HpyCH4IV sites, respectively. Comparative signals between tissues and significantly different regions are indicated as plots and gray bars, respectively. L, liver; K, kidney; C, cerebrum. B, combined bisulfite restriction analysis of T-DMRtags. The bisulfite PCR products were digested with HpyCH4IV and electrophoresed on a 2.0% agarose gel to evaluate the methylation status of T-DMRtags. The conditions of undigested (-) and digested (+) are indicated on the top of the gel. Because only unmethylated cytosine residues are converted to thymines by the bisulfite reaction, PCR fragments from unmethylated genomic DNA are resistant to HpyCH4IV digestion, whereas fragments from methylated DNA are digested. The representative image of at least two independent experiments is shown.

mRNA expression profiles of the SLC and ABC transporters. mRNA expression of Oatp1b2, Ntcp, Bsep, Abcg5, Abcg8, Pept2, and β-actin in mouse liver, kidney, and cerebrum was determined by RT-PCR using specific primers (Table 1) as described under Materials and Methods. An equal amount of total RNA was used for each sample, and distilled water was included as a negative control (NC). The identity of the PCR products is shown on the left.
Discussion
The expression of transporters is an important determinant of the physiological functions of liver and kidney, including the efficient turnover of endogenous compounds and elimination of xenobiotics. Although the involvement of epigenetic machineries in the kidney-specific expression of transporters has emerged, those in the liver-specific transporters have not been investigated. We examined comprehensively the epigenetic signatures of SLC and ABC transporters in mice, focusing particularly on those expressed exclusively in the liver.
Analysis with D-REAM and subsequent bisulfite genomic sequencing located T-DMRs that are hypomethylated in the liver but hypermethylated in the kidney or cerebrum over a wide range of genomic regions around the TSS of liver-specific transporters (Figs. 1 and 3). The histone H3 acetylation profiles neighboring the TSS also coincided with the tissue-specific expression of these transporters (Fig. 4). These results strongly suggest that the constitutive expression of liver-specific transporters involves the epigenetic signatures comprising DNA methylation and chromatin modification. Together with our previous findings (Kikuchi et al., 2006, 2007) and our ongoing study of the kidney-specific transporters (R. Kikuchi, S. Yagi, H. Kusuhara, Y. Sugiyama, K. Shiota, unpublished observations), the epigenetic regulation should be prevalent among transporters expressed in the liver and kidney.
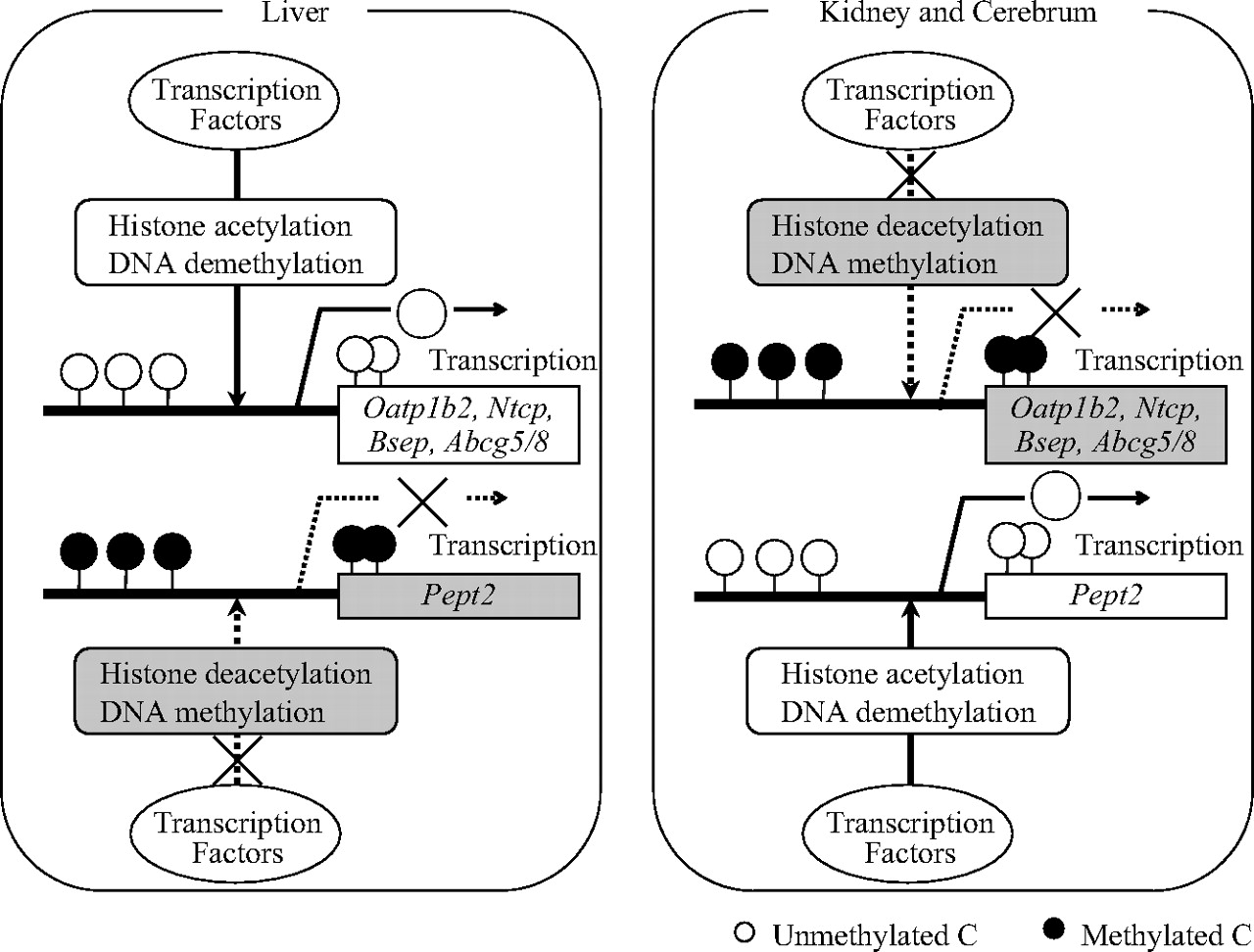
Promoter analysis of liver-specific transporters has unveiled the involvement of several transcription factors in mice, including HNF1α (Jung et al., 2001; Maher et al., 2006), glucocorticoid receptor (Eloranta et al., 2006), farnesoid X receptor (Sinal et al., 2000; Ananthanarayanan et al., 2001; Plass et al., 2002), and HNF4α (Sumi et al., 2007), which are responsible for the promoter activation of Oatp1b2, Ntcp, Bsep, and Abcg5/g8, respectively. According to the computational analysis with MatInspector (available at http://www.genomatix.de/), the position of the potential transcription factor binding motifs relative to the TSS of each transporter is as follows: HNF1-motif in Oatp1b2 promoter, -67 to -51; glucocorticoid response element in Ntcp promoter, -65 to -47; farnesoid X receptor response element in Bsep promoter, -92 to -80; and HNF4α-motif (DR-1 motif) in Abcg5/g8 intergenic region, -126 to -114 relative to the TSS of Abcg5 (-94 to -82 relative to the TSS of Abcg8). These binding motifs do not contain CpG dinucleotides but are surrounded by differentially methylated CpG dinucleotides in different tissues. Together with the different histone acetylation profiles in various tissues, it is likely that DNA methylation interrupts the binding of transcription factors indirectly through the recruitment of histone deacetylases and the subsequent chromatin condensation (Fig. 5).
It is interesting that our data emphasize that epigenetic regulation underlies the liver-specific expression of functionally cooperated transporters. Ntcp and Bsep are responsible for the sinusoidal uptake and canalicular efflux of bile acids in hepatocytes, respectively (Trauner and Boyer, 2003; Mita et al., 2006; Alrefai and Gill, 2007). Simultaneous liver-specific expression of these transporters is indispensable for the vectorial transport of bile acids under physiological conditions. Likewise, Abcg5 and Abcg8 form a heterodimer to function as the efflux transporter for sitosterols and cholesterols in the liver and to prevent the intestinal absorption of sitosterols (Hazard and Patel, 2007; Kidambi and Patel, 2008). The genes for these transporters are located in close proximity but with opposite orientation in the genome in both the human and mouse, and the intergenic region in human homologs has bidirectional promoter activity. The common regulatory mechanism, including the epigenetic traits and probably transcription factors, should lead to the same tissue distribution pattern of these two sets of functionally paired transporters.
It is noteworthy that members of the Oatp family, such as Oatp1a1, Oatp1a4, Oatp1a5, Oatp1a6, Oatp1b2, and Oatp1c1, exist in tandem on the same chromosome (Hagenbuch and Meier, 2004). Oatp1a1, Oatp1a4, Oatp1a5, and Oatp1a6 show particularly high sequence homology at the amino acid level to each other (>70%) and also show moderate sequence homology to Oatp1b2 and Oatp1c1 (>40%) (Hagenbuch and Meier, 2003), indicating that these isoforms are products of genetic duplication from the same ancestor gene. These Oatp isoforms show overlapping substrate specificities and are functionally redundant, except for Oatp1c1, which is relatively specific for thyroid hormone (Tohyama et al., 2004). Each member exhibits a unique tissue distribution (e.g., Oatp1a1 is expressed predominantly in the liver and kidney; Oatp1a4 and Oatp1a5 are expressed in the liver, kidney, and cerebrum; and Oatp1b2, Oatp1a6, and Oatp1c1 are expressed exclusively in the liver, kidney, and cerebrum, respectively). The liver-specific expression of Oatp1b2 is associated with its epigenetic signature, indicating that the difference in the epigenetic signature distinguishes each isoform of the Oatp family transporters in terms of its tissue distribution, thus conferring functional diversity from the same ancestor gene during the evolutional process. Future studies should show the in vivo relevance of epigenetic regulation for other Oatp transporters.

DNA methylation profiles around the transcriptional start site. Bisulfite genomic sequencing analysis was performed for Oatp1b2 (A), Ntcp (B), Bsep (C), Abcg5/g8 (D), and Pept2 (E) with genomic DNA extracted from mouse liver, kidney, and cerebrum as described under Materials and Methods. Top, a schematic diagram of the genomic region around the TSS. The vertical lines and numbers indicate the positions of the cytosine residues of the CpG dinucleotides relative to the TSS (+1). Bottom, DNA methylation status of individual CpG dinucleotides. The open and closed circles represent unmethylated and methylated cytosines, respectively.

Histone acetylation profiles in the promoter region. The histone H3 acetylation status of Oatp1b2, Ntcp, Bsep, Abcg5/g8, Pept2, and β-actin in the promoter region was determined by ChIP assay in mouse liver, kidney, and cerebrum as described under Materials and Methods. The closed columns represent the histone acetylation of each sample relative to its respective total input. Normal rabbit IgG was included as a negative control to verify the immunoprecipitation specificity (□). The results are presented as the mean ± S.E. of triplicate experiments.

Proposed mechanism underlying the transcriptional regulation of the tissue-specific transporters. DNA methylation neighboring the TSS evokes the condensed structure of chromatin through the deacetylation of histones, which interrupts the binding of transcription factors and consequently suppresses the expression of corresponding transporters in the tissue-specific manner.
In conclusion, our study revealed strong associations between the liver-specific expression and epigenetic profiles comprising DNA methylation and histone acetylation for Oatp1b2, Ntcp, Bsep, and Abcg5/g8 in mice. Together with our previous findings on the kidney-specific transporters, it is plausible that epigenetic regulation is a mechanism with considerable universality that underlies the tissue-specific expression of SLC and ABC transporters.
Footnotes
-
ABBREVIATIONS: SLC, solute carrier; ABC, ATP binding cassette; Bsep, bile salt export pump; ChIP, chromatin immunoprecipitation; D-REAM, tissue-dependent differentially methylated region profiling with restriction tag-mediated amplification; HNF, hepatocyte nuclear factor; Ntcp, Na+-taurocholate-cotransporting polypeptide; Oatp, organic anion transporting polypeptide; PCR, polymerase chain reaction; RT-PCR, reverse transcription polymerase chain reaction; Pept2, peptide transporter 2; T-DMR, tissue-dependent differentially methylated region; T-DMRtag, tag of the tissue-dependent differentially methylated region; TSS, transcriptional start site.
-
↵
 The online version of this article (available at http://molpharm.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://molpharm.aspetjournals.org) contains supplemental material. - Received October 7, 2008.
- Accepted December 1, 2008.

- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}