


Abstract
The phytoestrogen resveratrol has putative health-promoting effects and is present in several dietary constituents. Resveratrol is metabolized extensively in the gut epithelium, resulting in the formation of hydrophilic glucuronic acid and sulfate conjugates. These polar resveratrol conjugates need specific transporters to cross the cell membrane. We show here that vectorial transport of some of these metabolites is mediated by multidrug resistance protein 3 (MRP3, ABCC3) and/or breast cancer resistance protein (BCRP, ABCG2) located in the basolateral and apical membranes of enterocytes, respectively. In vitro, MRP3 transports resveratrol-glucuronide (Res-3-G). The absence of Mrp3 in mice results in altered disposition of Res-3-G and its parent compound resveratrol, leading to a reduced percentage of resveratrol being excreted via the urine in Mrp3(-/-) mice. Circumstantial evidence suggests that circulating resveratrol is formed by deglucuronidating Res-3-G in vivo, providing a possible explanation for the health beneficial effects of resveratrol in vivo, despite its rapid and extensive conjugation. BCRP transports Res-3-G and resveratrol sulfates in vitro, and its absence in mice results in high plasma levels of resveratrol-di-sulfate, a resveratrol metabolite hardly detectable in the plasma of wild-type mice and in an increased disposal of resveratrol via the urine. The profound effects of ATP-binding cassette transporters on the disposal of resveratrol may be representative for the handling of several other polyphenols of dietary origin.
Resveratrol is a phytoestrogen with postulated health-beneficial effects that is present in foods such as grapes and peanuts (Baur and Sinclair, 2006). The mammalian gastrointestinal tract provides a barrier for the uptake of resveratrol and other xenobiotics of dietary origin (Kaminsky and Zhang, 2003), and important components of this barrier are the phase I and phase II metabolizing enzymes, together with associated transporters (phase III metabolism) (Xu et al., 2005). Oral resveratrol is metabolized in the gut wall (Andlauer et al., 2000; Kuhnle et al., 2000) and is found in plasma and urine mainly as sulfo- and glucuronosyl-conjugates (Walle et al., 2004; Wenzel et al., 2005).
Many of the health-promoting effects attributed to resveratrol have been found in experiments with lower organisms and/or test tubes (Howitz et al., 2003; Baur and Sinclair, 2006), in which its actions were attributable to unmodified resveratrol. Modifications such as glucuronidation and sulfation normally result in compounds that have less pharmacological activity (Tukey and Strassburg, 2000; Kaminsky and Zhang, 2003; Rimbach et al., 2004). The extensive modification of resveratrol in the gut wall has therefore led to skepticism about its therapeutic potential in humans after oral dosing (Walle et al., 2004; Wenzel and Somoza, 2005; Wenzel et al., 2005; Baur and Sinclair, 2006). Recent studies indicate, however, that dietary resveratrol can have beneficial effects in mice (Baur et al., 2006; Lagouge et al., 2006). It is therefore possible that the resveratrol metabolites that enter the circulation retain (some) pharmacological activity or that deconjugation of these metabolites occurs, thereby releasing the pharmacologically active parent compound. A better understanding of the factors involved in the tissue distribution of resveratrol and its metabolites would therefore help to identify the target organs for resveratrol and allow a better prediction of its pharmacological potency.
After their intracellular formation in enterocytes, sulfo- and glucuronic acid conjugates of resveratrol are too hydrophilic to passively diffuse over the plasma membrane, and specific transporters are implied in their extrusion from the cell (Andlauer et al., 2000; Kuhnle et al., 2000). Recently, hepatic Mrp2 was shown to mediate the canalicular efflux of resveratrol-glucuronides from rat liver (Maier-Salamon et al., 2007). Resveratrol-glucuronides are excreted over the basolateral membrane of hepatocytes as well (Maier-Salamon et al., 2007), but the responsible transporter remains to be identified. The transporters involved in the cellular extrusion of resveratrol-sulfates also are not yet known.
Two transport proteins that have been shown previously to mediate the transport of phase II metabolites are multidrug resistance protein 3 (MRP3, ABCC3) and breast cancer resistance protein (BCRP, ABCG2), both belonging to the ATP-binding cassette (ABC) superfamily of membrane transporters (Borst and Oude Elferink, 2002). Both proteins have a similar tissue distribution and are expressed in liver, kidney, and gut, organs known for their high phase II metabolizing capacity (Tukey and Strassburg, 2000). In polarized cells, MRP3 and BCRP route to the basolateral and apical membrane, respectively, as also found in the enterocytes of the gut (Scheffer et al., 2002; Haimeur et al., 2004). In vitro studies have shown that MRP3 and BCRP are typical organic anion transporters (Kool et al., 1999; Borst and Oude Elferink, 2002; Kruh and Belinsky, 2003; Haimeur et al., 2004). Substrates of both transporters include compounds conjugated to sulfate, glucuronate, or glutathione (GSH) (i.e., phase II conjugation products) (Borst and Oude Elferink, 2002; Kruh and Belinsky, 2003). MRP3 seems to have a preference for glucuronidated substrates (Borst et al., 2003; Kruh and Belinsky, 2003; Manautou et al., 2005; Zamek-Gliszczynski et al., 2006; Zelcer et al., 2006), whereas the substrate spectrum of BCRP is broader and includes many compounds containing a sulfate moiety (Borst and Oude Elferink, 2002; Schinkel and Jonker, 2003; Haimeur et al., 2004; Adachi et al., 2005). Hence, MRP3 and BCRP are involved in the protection of the body against xenobiotics by transporting xenobiotic conjugates out of the cell for subsequent urinary (MRP3) or fecal (BCRP) excretion (Schinkel and Jonker, 2003; Haimeur et al., 2004; Borst et al., 2007).
Intestinal BCRP limits the systemic exposure to several xenobiotics (including many important drugs) and xenobiotic conjugates after oral exposure (Schinkel and Jonker, 2003; Adachi et al., 2005). Less is known about the function of intestinal MRP3, but its basolateral localization implies that it transports its substrates toward the circulation for subsequent urinary excretion. Indeed, hepatic MRP3 is involved in the transport of several glucuronidated compounds to the circulation for subsequent urinary excretion (Manautou et al., 2005; Zelcer et al., 2005, 2006; Zamek-Gliszczynski et al., 2006; van de Wetering et al., 2007).
Because intestinal BCRP and MRP3 are optimally placed to handle sulfo- and glucuronic acid conjugates of resveratrol, we have used vesicular transport experiments to test the interaction of these transporters with resveratrol metabolites and studied the effect of a complete loss of Bcrp1 and Mrp3 on resveratrol pharmacokinetics.
Materials and Methods
[3H]Resveratrol (3.6 Ci/mmol) was from Moravek Biochemicals (Brea, CA). [3H]Estradiol-17β-d-glucuronide (E217βG) and [3H]methotrexate (MTX) were from PerkinElmer Life and Analytical Sciences (Waltham, MA). MultiScreenHTS MSFB filter plates (96 wells) and the MultiScreenHTS vacuum manifold were from Millipore (Amsterdam, the Netherlands). Oasis HLB solid-phase extraction cartridges were from Waters (Milford, MA). The Luna C18(2) reversed-phase column was from Phenomenex (Torrance, CA). Dulbecco's modified Eagle's medium, Sf-900 II SFM medium, Opti-MEM, penicillin/streptomycin, and fetal calf serum (FCS) were from Invitrogen (Breda, the Netherlands). Transwell Costar plates were from Corning Life Sciences (Acton, MA). All other chemicals and reagents were from Sigma Chemicals (St. Louis, MO).
Synthesis of Resveratrol Metabolites. Nonradiolabeled resveratrol metabolites were synthesized as described previously (Wenzel et al., 2005). [3H]Resveratrol-glucuronide was synthesized from [3H]resveratrol using mouse liver microsomes. Preparation of mouse liver microsomes and subsequent in vitro generation of [3H]resveratrol-glucuronide was done as described previously for morphine (van de Wetering et al., 2007). Completeness of conversion to [3H]resveratrol-glucuronide was determined by high-pressure liquid chromatography (HPLC) and liquid scintillation counting (LSC). Conversion of [3H]resveratrol to [3H]resveratrol-glucuronide after glucuronidation was complete because no radioactivity corresponding to the aglycon was detected, and apart from resveratrol-glucuronide, no additional peaks were present in the radiochromatogram (data not shown). Recovery of resveratrol-glucuronide after synthesis was 95%. [3H]Resveratrol-3-sulfate (Res-3-S) and [3H]resveratrol-di-sulfate (Res-di-S) were generated by incubation of 10 μCi of [3H]resveratrol with 450 μg of cytosolic protein from mouse liver in the presence of 100 μM 3′-phosphoadenosine-5′-phosphosulfate in 100 mM phosphate buffer, pH 7.4, for 18 h at 37°C. [3H]Resveratrol-sulfates were extracted from the reaction mix by solid-phase extraction on Oasis HLB cartridges, and [3H]Res-3-S and [3H]Res-di-S were purified by preparative HPLC on a Luna C18(2) 3.0-μm column (150 × 4.6 mm) (isocratic elution in 19% acetonitrile in 10 mM ammonium formate, pH 8.2 for 25 min). Purified [3H]resveratrol-sulfates were subsequently concentrated by freeze-drying.
Cell Lines and Culture Conditions. Sf9 insect cells in suspension were grown in Sf-900 II SFM medium containing 10% FCS. HEK293 cells expressing hMRP3 were generated by transfecting HEK293 cells with pCMVneoMRP3 (Kool et al., 1999) using calcium phosphate precipitation. After 48 h, cells were split, and G418 (800 μg/ml) was used to select for clones that express hMRP3. G418-resistant clones were tested for MRP3 expression by immunoblot analysis using the M3II9 antibody (Scheffer et al., 2002). Plasma membrane localization was confirmed by immunofluorescence microscopy using the same antibody (data not shown). HEK293 parental and HEK293-MRP3 cells were grown in Dulbecco's modified Eagle's medium supplemented with 10% FCS and 100 U penicillin/streptomycin/ml.
Preparation of Membrane Vesicles. Membrane vesicles from Sf9 cells were obtained after infection with a BCRP (Breedveld et al., 2007) or control cDNA containing baculovirus at a multiplicity of infection of 1. After incubation at 27°C for 3 days, cells were harvested by centrifugation at 500g for 5 min. The pellet was resuspended in ice-cold hypotonic buffer (0.5 mM sodium phosphate and 0.1 mM EDTA, pH 7.4) supplemented with a protease inhibitor cocktail (Roche, Basel, Switzerland) and incubated at 4°C for 90 min. The suspension was centrifuged at 100,000g for 40 min, and the pellet was homogenized in ice-cold TS buffer (50 mM Tris-HCl and 250 mM sucrose, pH 7.4) using a tight-fitting Dounce homogenizer. After centrifugation at 500g at 4°C for 10 min, the supernatant was collected and centrifuged at 4°C at 100,000g for 40 min. The pellet was resuspended in TS buffer and passed through a 27-gauge needle 25 times. The vesicles were dispensed in aliquots, snap-frozen in liquid nitrogen, and stored at -80°C until use.
Vesicles of HEK293-MRP3 and HEK293 control cells were prepared as described for the Sf9 vesicles with 1 modification: the final homogenization buffer was 10 mM Tris-HCl, pH 7.4, instead of the 50 mM Tris-HCl/250 mM sucrose, pH 7.4, used for the Sf9 vesicles.
Vesicular Transport Assays. The time- and concentration-dependent transport of various substrates into membrane vesicles was studied using the rapid filtration method as described previously (van de Wetering et al., 2007). Ki values were calculated using the following equation: Km′= Km*(1 + [I]/Ki), with Km′ being the apparent Km determined in the presence of inhibitor [I] and the concentration of the inhibitor expressed in micromoles.
Animals.Bcrp1(-/-) (Jonker et al., 2002) and Mrp3(-/-) (Zelcer et al., 2006) mice have been described previously, were on a 99% FVB background, received food and water ad libitum, and were housed in constant temperature rooms with a 12-h light/dark cycle. Mouse handling and experimental procedures were conducted in accordance with institutional guidelines for animal care and use.
Analysis of Resveratrol Pharmacokinetics in Mice. Male age- and weight-matched mice were used throughout the study and were fasted for 4 h before the onset of the experiments. Resveratrol (50 mg/kg body weight; vehicle. 0.5% hydroxypropylmethyl cellulose) was administered intragastrically by oral gavage feeding. For the time course experiments, four to five mice of each group were sacrificed at each time point by cardiac puncture and blood sampling in heparinized syringes under methoxyflurane anesthesia. Livers and kidneys were collected and immediately stored at -80°C until analysis. Plasma was obtained after centrifugation for 10 min at 1500g at 22°C and stored at -80°C until analysis. To analyze the urinary excretion of resveratrol and its metabolites, mice were placed in metabolic cages, resveratrol was administered as described above, and urine was collected for 24 h. At the end of the experiment, mice were sacrificed as mentioned above.
Quantification of Resveratrol and Its Metabolites. Resveratrol and resveratrol metabolites in plasma, urine, and tissues were determined as described previously (Burkon and Somoza, 2008).
Tissue Distribution of [3H]Resveratrol. [3H]resveratrol (50 mg/kg; 1.5-2 μCi per mouse) was administered as described above, and after 24 h, mice were sacrificed as described above, and tissues were collected and solubilized in 0.5 M NaOH/0.1% Triton X-100. After solubilization, samples were bleached using 30% H2O2. The level of radioactivity in tissue homogenates was determined by LSC using Ultima Gold scintillation fluid (PerkinElmer Life and Analytical Sciences). Urine was counted by directly adding LSC fluid without processing.
Statistical Analysis. In Figs. 1 and 2, nonlinear regression and statistical analyses were performed using GraphPad Prism software (San Diego, CA). Statistical analysis was done by testing the null hypothesis that the fits of the curves obtained in the presence of inhibitor did not significantly differ from the curve obtained in the absence of inhibitor. For statistical analysis of the pharmacokinetic experiments, one-way analysis of variance was used, including a Dunnett's multiple comparison test. Statistically significant differences from control values are indicated in the figures with *, p < 0.5, **, p < 0.01, and ***, p < 0.001.
Results
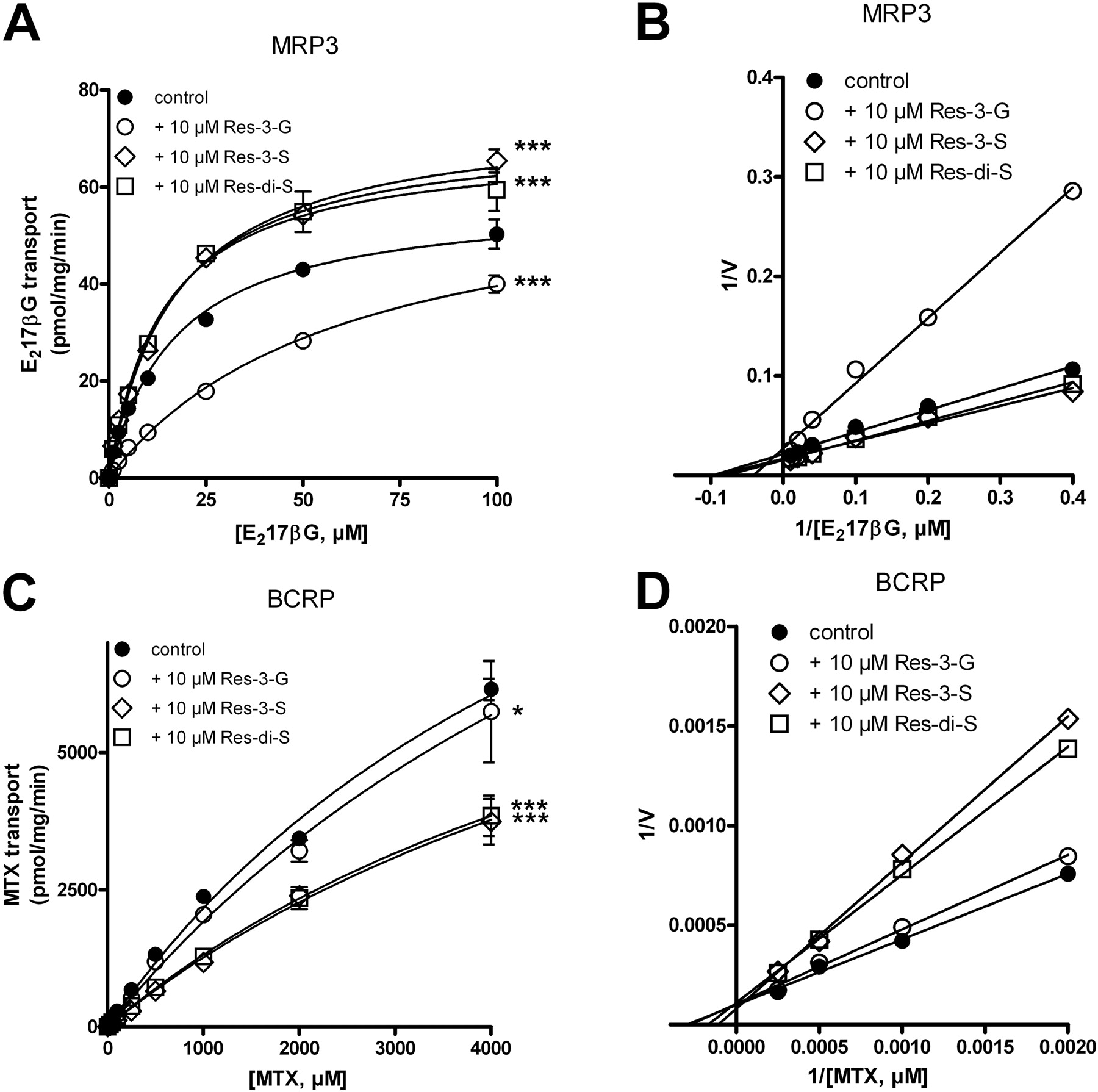
Effect of Resveratrol Metabolites on MRP3- and BCRP-Mediated Transport in Vesicular Transport Experiments. In vesicular transport experiments, competition with a known substrate is often used to screen for new substrates of membrane transporters: competitive inhibition is an indication that both compounds compete for the same binding/transport site on the transporter. We therefore studied the effects of resveratrol metabolites on MRP3-mediated transport of E217βG, a prototype MRP3 substrate. At a concentration of 10 μM, only resveratrol-3-glucuronide (Res-3-G) inhibited the transport of E217βG (Fig. 1A). Inhibition could be overcome by increasing the substrate concentration, compatible with competitive inhibition. In addition, Lineweaver-Burke transformations also indicated that inhibition of MRP3-mediated E217βG transport by Res-3-G was competitive (Fig. 1B), which was confirmed by nonlinear regression analysis yielding apparent Km values of 16.9 and 60.0 μM in the absence and presence of Res-3-G, respectively, without significant alteration of the Vmax. The Ki value is approximately 8 μM, suggesting that Res-3-G is a relatively high-affinity substrate of MRP3. It is noteworthy that both resveratrol sulfo-conjugates tested, Res-3-S and Res-di-S, moderately stimulated MRP3-mediated transport of E217βG by increasing the Vmax without significantly altering the Km (Fig. 1A). We will come back to this stimulatory effect of the resveratrol-sulfates under Discussion.
[3H]MTX was used as substrate to study the ability of resveratrol-metabolites to interact with BCRP. At a concentration of 10 μM, both Res-3-S and Res-di-S inhibited BCRP-mediated transport of MTX, whereas higher concentrations of Res-3-G were needed to inhibit transport (Fig. 1C and data not shown). Because we can not saturate MTX transport by BCRP, we were unable to determine whether inhibition of BCRP-mediated transport by resveratrol metabolites is competitive. Lineweaver-Burke transformation, however, suggested that competition by Res-3-S and Res-di-S was competitive because their curves crossed the y-axis at the same position as the curve obtained in the absence of inhibitor, whereas the position at which the x-axis was crossed differed, indicative of competitive inhibition (Fig. 1D).
Transport of Res-3-G into MRP3- and BCRP-Containing Vesicles. Because our inhibition experiments indicated that Res-3-G was a substrate of MRP3, we enzymatically synthesized [3H]Res-3-G and tested whether this could serve as a substrate for this transporter. MRP3 indeed transported Res-3-G in a time- and ATP-dependent manner (Fig. 2A). Moreover, transport followed Michaelis-Menten kinetics (Fig. 2C), and nonlinear regression analysis yielded Km and Vmax values of 14 ± 2.2 μM and 72 ± 4.5 pmol/mg/min, respectively. The Km value is in good agreement with the Ki value we found for this compound in competition experiments.
Despite the fact that Res-3-G only weakly inhibited BCRP-mediated transport of MTX (Fig. 1C), BCRP readily transported [3H]Res-3-G in an ATP- and time-dependent way (Fig. 2B). Although we could not completely saturate BCRP-mediated transport of Res-3-G at the concentration range tested (1-200 μM), the approximate Km and Vmax values were roughly 120 ± 15 μM and 340 ± 22 pmol/mg/min, respectively. This low affinity explains why Res-3-G only weakly inhibited BCRP-mediated transport of MTX.
Transport of Res-3-S and Res-di-S by BCRP. Because inhibition experiments indicated that Res-3-S and Res-di-S are substrates of BCRP, we tested whether there was direct transport of the purified [3H]Res-3-S and Res-di-S. At 37°C, the reaction rate for BCRP-mediated transport of both resveratrol sulfates into the vesicles was too fast to reliably determine initial rates of transport (data not shown), and to overcome this problem, we carried out all reactions at 21°C. Both Res-3-S and Res-di-S, were transported by BCRP in a time- and ATP-dependent manner (Fig. 3, A and B). Transport of the resveratrol sulfates by BCRP followed Michaelis-Menten kinetics (Fig. 3, C and D), with Res-3-S transport having a Km value of 5.0 ± 1.3 μM and a Vmax value of 1060 ± 69 pmol/mg/min and Res-di-S transport a Km value of 10 ± 2.0 μM and a Vmax value of 1600 ± 94 pmol/mg/min. This shows that both resveratrol sulfates are transported by BCRP with high affinity and high capacity. In vivo transport rates at 37°C will probably be even higher, because the in vitro reactions were carried out at 21°C.

Inhibition of MRP3- and BCRP-mediated transport by resveratrol metabolites. A, concentration-dependent transport of [3H]E217βGin MRP3-containing membrane vesicles in the presence or absence of 10 μM Res-3-G, Res-3-S, or Res-di-S as indicated. C, concentration-dependent transport of [3H]MTX by BCRP in the presence or absence of 10 μM Res-3-G, Res-3-S, or Res-di-S. Values are corrected for transport in the absence of ATP. Each data point and error are the mean ± S.D. of a representative experiment of two independent experiments each done in triplicate. B and D are Lineweaver-Burk transformations of the data presented in A and C, respectively.
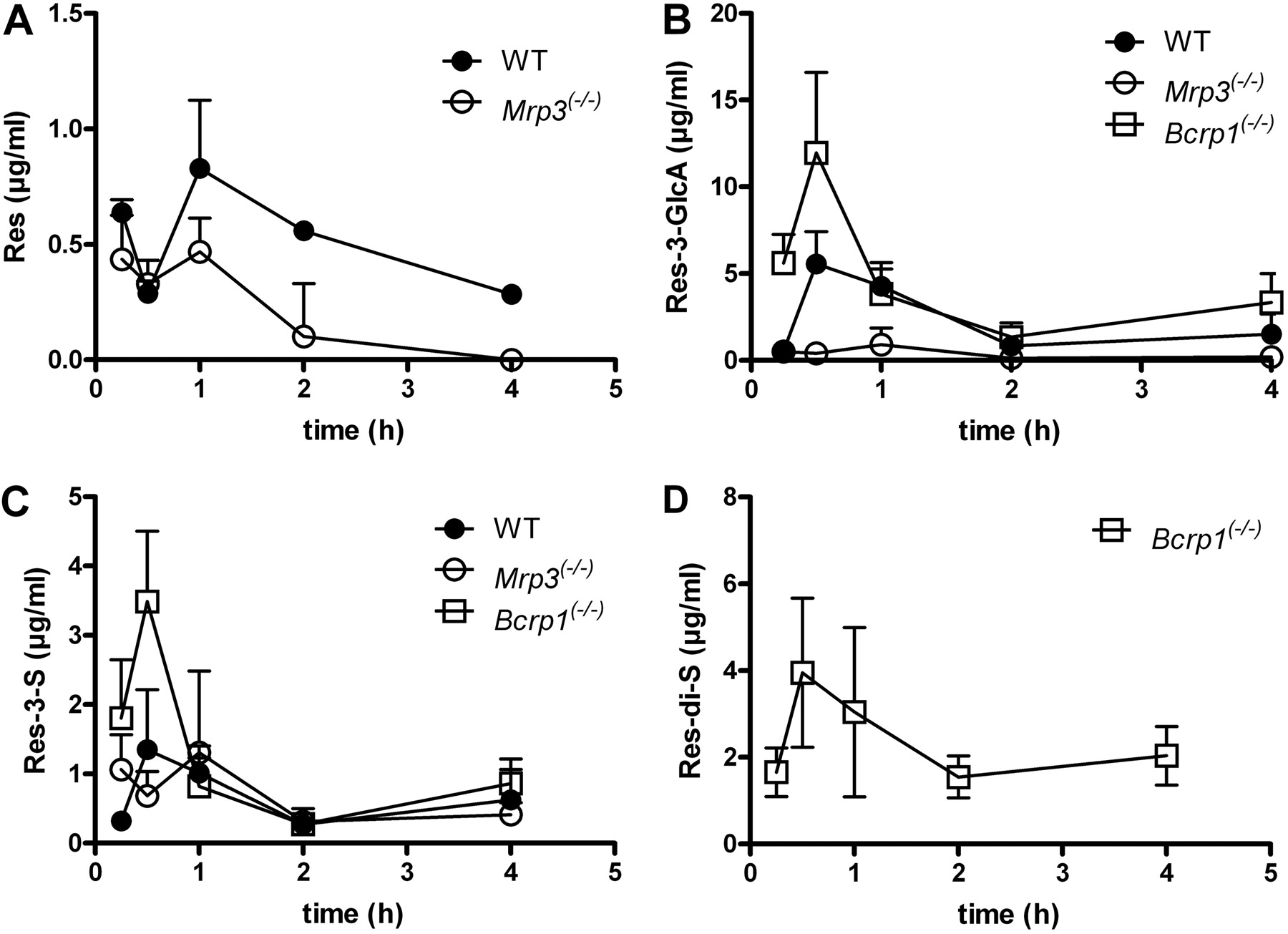
Pharmacokinetics of Resveratrol and Its Metabolites in Wild-Type, Bcrp1(-/-), and Mrp3(-/-) Mice. We chose a dose of 50 mg of resveratrol/kg body weight because resveratrol has been shown to have health-beneficial effects in mice given at daily doses ranging from approximately 40 to 400 mg/kg (Baur et al., 2006; Lagouge et al., 2006). After the oral administration of resveratrol, its main circulating metabolites in WT mice were found to be Res-3-G and Res-3-S, whereas Res-di-S was not detected in plasma and was only present in low amounts in 24-h urine samples. Plasma levels of resveratrol and its metabolites followed a biphasic elimination pattern, suggestive of enterohepatic circulation of resveratrol (Fig. 4).
In the absence of Mrp3, plasma levels of the main resveratrol metabolite Res-3-G were reduced 10-fold (Fig. 4B), and this was accompanied by a similar reduction in its urinary excretion (Fig. 5B). The plasma and urinary excretion levels of the other resveratrol metabolites were unaltered (Res-di-S) or slightly reduced (Res-3-S) in Mrp3(-/-) mice relative to wild-type mice (Figs. 4 and 5). The absence of Mrp3 in mice also resulted in an unexpected 10-fold reduction in the levels of unconjugated resveratrol in urine (Fig. 5A). Although plasma levels of resveratrol were similar in WT and Mrp3(-/-) mice for the first 30 min after resveratrol administration, subsequent time points after dosing showed lower resveratrol plasma levels in the absence of Mrp3 (Fig. 4A). These lower levels of resveratrol cannot be due to a direct effect of Mrp3 on the transport of resveratrol, because we have been unable to demonstrate the transport of resveratrol by human MRP3 in Madin-Darby canine kidney II cells (Supplemental Fig. 3B). Neither did resveratrol inhibit E217βG transport by human (Supplemental Fig. 2B) or mouse (data not shown) MRP3/Mrp3 in vesicular transport experiments.

Transport of Res-3-G by BCRP and MRP3 in vesicular transport experiments. Time-course experiments of membrane vesicles containing human MRP3 (A) or BCRP (B) with 1 μM [3H]Res-3-G and control vesicles as indicated. Transport was determined in the presence or absence of 4 mM ATP as indicated. Concentration-dependent transport of Res-3-G in MRP3-(C) and BCRP-containing (D) vesicles. Values are corrected for transport determined in absence of ATP. Each data point and error are the means ± S.D. of an experiment performed in triplicate.
BCRP is present at the apical membrane of the enterocyte and limits the uptake of its substrates from the gut (van Herwaarden and Schinkel, 2006). The absence of Bcrp1 also altered the pharmacokinetics of resveratrol metabolites in mice, resulting in slightly increased plasma concentrations of Res-3-G and Res-3-S (Fig. 4). However, the most salient finding in Bcrp1(-/-) mice was the dramatically increased plasma level of Res-di-S, a metabolite undetectable in plasma samples of WT and Mrp3(-/-) mice (Fig. 4D). This led to a more than 20-fold increase in the urinary excretion of Res-di-S in Bcrp1(-/-) mice (Fig. 5D) and an increase in the fraction of the administered resveratrol dose excreted via urine (Fig. 5E). These results indicate that Res-di-S is a transported BCRP/Bcrp1 substrate, in line with our in vitro studies showing that BCRP transports Res-di-S (Fig. 3, B and D) and that Res-di-S acts as an inhibitor of BCRP-mediated MTX transport (Fig. 1C). The elevated plasma levels of Res-3-G and Res-3-S in the Bcrp1(-/-) mice were not accompanied by higher urinary excretion of these compounds. We will return to this result under Discussion. Although BCRP is known to transport unconjugated resveratrol (Breedveld et al., 2007), we did not detect increased circulating levels of resveratrol in Bcrp1(-/-) mice or the increased urinary excretion of nonmetabolized resveratrol. On the contrary, we found somewhat reduced amounts of resveratrol excreted via the urine, and resveratrol was lower than the limit of detection in plasma samples of Bcrp1(-/-) mice.
Levels of resveratrol and its metabolites were also determined in livers and kidneys 1 h after the administration of resveratrol (Fig. 6). Although Mrp3(-/-) mice had approximately 10-fold lower plasma levels of Res-3-G than WT mice 1 h after resveratrol administration, Res-3-G levels in their livers were only 2-fold reduced (Fig. 6C). Liver levels of resveratrol were similar in WT and Mrp3(-/-) mice (Fig. 6A), even though the absence of Mrp3 led to 50% lower plasma concentrations of resveratrol 1 h after administration (Fig. 4A). No differences in liver concentrations of Res-3-S were found either (Fig. 6E). In kidneys, the concentration of resveratrol and its metabolites reflected levels found in plasma 1 h after resveratrol administration, with the parent compound resveratrol being undetectable in kidney samples of Mrp3(-/-) mice.
Excretion of [3H]Resveratrol in Wild-Type, Bcrp1(-/-), and Mrp3(-/-) Mice. We also administered [3H]resveratrol intragastrically to mice and determined the amount of radiolabel in 24-h urine and feces samples of Mrp3(-/-), Bcrp1(-/-), and WT mice (Fig. 7). As expected, Mrp3(-/-) mice excreted less radiolabel via the urine, whereas urinary excretion rates of resveratrol (metabolites) in Bcrp1(-/-) mice were increased (Fig. 7A). These alterations were mirrored by the changes in excretion via the feces: increased fecal excretion in the absence of Mrp3, and reduced fecal excretion in Bcrp1(-/-) mice (Fig. 7B). This shows that the absence of Mrp3 results in a shift from urinary to fecal excretion of resveratrol (metabolites), whereas in Bcrp1(-/-) mice, the opposite occurs: higher urinary excretion rates, and lower fecal excretion. These results are in line with analysis of the urinary resveratrol-(metabolite) profile by HPLC/DAD. Total recovery of resveratrol and its metabolites in urine was somewhat lower when analyzed by HPLC/DAD (Fig. 5E) than by LSC. We attribute this to our inability to detect some minor resveratrol metabolites by HPLC/DAD analysis such as sulfo-resveratrol-glucuronide and resveratrol-di-glucuronide. The latter was recently identified as one of the resveratrol metabolites present in human plasma (Burkon and Somoza, 2008).

Transport of Res-3-S and Res-di-S by BCRP in vesicular transport experiments. Time-course experiments of membrane vesicles containing human BCRP using 140 nM [3H]Res-3-S (A) or [3H]Res-di-S (B). Transport was determined in the presence or absence of 4 mM ATP as indicated. Concentration-dependent transport of Res-3-S and Res-di-S by BCRP is shown in C and D, respectively. Values are corrected for transport determined in the absence of ATP. Each data point and error are the means ± S.D. of an experiment performed in triplicate. Note that transport was determined at 21°C.

Pharmacokinetics of resveratrol in WT, Mrp3(-/-), and Bcrp1(-/-) mice. Groups of mice (n = 4 or 5 per time point) received a dose of 50 mg of resveratrol/kg body weight via oral gavage feeding. Plasma concentrations of resveratrol (A), Res-3-G (B), Res-3-S (C), and Res-di-S (D) were determined at the time points indicated. Values shown are means ± S.D.

Urinary excretion of resveratrol and its metabolites in WT, Mrp3(-/-), and Bcrp1(-/-) mice. Groups of mice (n = 5-9 per genotype) received an oral dose of 50 mg of resveratrol/kg body weight and the excretion of resveratrol (A), Res-3-G (B), Res-3-S (C), and Res-di-S (D) over 24 h was determined. In E, the summarized excretion of resveratrol and its metabolites is shown. Values are expressed as a percentage of the dose of resveratrol administered and are mean ± S.D.
Discussion
Our experiments identify BCRP and MRP3 as two major determinants of the pharmacokinetics of the phytoestrogen resveratrol and its metabolites. BCRP was already known to transport resveratrol itself (Breedveld et al., 2007). We show here that it also transports resveratrol sulfates and Res-3-G, albeit the latter substrate with low affinity. Res-3-G is a high-affinity substrate for MRP3, however, in line with the known preference of this transporter for glucuronidated substrates (Borst et al., 2007).
Liver, kidney, and the gastrointestinal tract are organs with a high phase II-metabolizing activity (Tukey and Strassburg, 2000; Gamage et al., 2006). In male mice, Mrp3 is present in liver and gastrointestinal tract but absent in kidney (Zelcer et al., 2006). After oral administration, resveratrol is detected predominantly in plasma and urine conjugated to glucuronic acid. Intestinal perfusion (Andlauer et al., 2000) and Ussing chamber experiments (Kuhnle et al., 2000) have shown that resveratrol is extensively glucuronidated in the gut wall. In addition, in humans, sulfotransferase activity toward resveratrol was higher in duodenum than in liver (De Santi et al., 2000). This indicates that after oral administration, the gastrointestinal tract is the major site for resveratrol metabolism.
The absence of Mrp3 or Bcrp1 has pronounced effects on the disposition of resveratrol in vivo. Mice lacking Mrp3 had up to 10-fold lower levels of Res-3-G in plasma and urine than control mice. Because resveratrol seems to be mainly glucuronidated in the gut, the decreased plasma Res-3-G must be due to the absence of Mrp3 in the basolateral membrane of the enterocytes. Absence of Bcrp1, normally located in the apical membrane of enterocytes, leads to elevated plasma levels of Bcrp1 substrates (van Herwaarden and Schinkel, 2006), as we also found for all resveratrol-metabolites in Bcrp1(-/-) mice, with the most pronounced effect on Res-di-S. The plasma concentrations of Res-3-G were also elevated. This could be due to stimulation of Mrp3-mediated transport of Res-3-G by the resveratrol-sulfates that accumulate within the enterocyte in the absence of Bcrp1. We have found that the sulfated resveratrol metabolites (Res-3-S and Res-di-S) stimulate MRP3-mediated transport of E217βGby increasing the Vmax. Stimulation has also been seen for other substrate combinations (Chu et al., 2004), and this stimulation of MRP3-mediated transport might therefore be a general feature of sulfo-conjugates.

Concentration of resveratrol and its metabolites in liver and kidney of WT and Mrp3(-/-) mice. Groups of mice (n = 4 or 5 per genotype) received an oral dose of 50 mg of resveratrol/kg body weight, and the concentrations of the indicated resveratrol (metabolites) were determined in liver (A, C, and E) and kidney (B, D, and F). Values shown are means ± S.D.

Determination of the urinary and fecal excretion of resveratrol in WT, Mrp3(-/-), and Bcrp1(-/-) mice. Groups of mice (n = 4 or 5 per genotype) received an oral dose of 50 mg of [3H]resveratrol/kg body weight, and 24 h later, the presence of [3H]label was determined in urine (A) and feces (B). In C, summarized excretion via urine and feces is shown. Values are expressed as a percentage of the dose of [3H]resveratrol administered and are the mean ± S.D.
In the Bcrp1(-/-) mice, the fraction of a dose of resveratrol excreted via the urine was increased due to the increased excretion of Res-di-S. The higher plasma levels of Res-3-G and Res-3-S in Bcrp1(-/-) mice were not accompanied by their higher urinary excretion, however. This apparent discrepancy might be explained by the presence of Bcrp1 in cells of the proximal tubule of the kidney (van Herwaarden and Schinkel, 2006), in which it might actively transport Res-3-G and Res-3-S toward the urine within the tubules, a process missing in Bcrp1(-/-) mice.
Potentially handling of resveratrol could be altered in the knockout mice by changes in the level of conjugating enzymes as a result of the absence of transporters. However, no such changes were found in microarray gene expression profiles of the knockout mice livers (data not shown).
Unmodified resveratrol was found to be substantially reduced in the plasma and urine of Mrp3(-/-) mice. Five explanations can be considered for this result. First, resveratrol is a substrate of MRP3. It does not act as a competitive inhibitor in vesicular transport experiments using vesicles containing MRP3, however (Supplemental Fig. 2B), and in transwell experiments, we did not detect transport either (Supplemental Fig. 3), clearly showing that resveratrol is not a substrate of MRP3/Mrp3. Second, the unmodified resveratrol transporter BCRP/Bcrp1 (Supplemental Fig. 3C; Breedveld et al., 2007) is more active in Mrp3(-/-) than in WT mice, resulting in less uptake of resveratrol from the gut. A major effect of Bcrp1 on uptake is improbable, however, because concentrations of resveratrol in plasma and urine of Mrp3(-/-) and Bcrp1(-/-)/Mrp3(-/-) mice were similar (data not shown). Third, decreased enterohepatic circulation (EHC) of resveratrol (metabolites) results, because less Res-3-G is available for EHC in the Mrp3(-/-) mice. If this were the case, we should also find substantially lower levels of Res-3-S and Res-di-S in plasma and/or urine of Mrp3(-/-) than of WT mice and this is not the result obtained (Figs. 4 and 5). An additional argument against reduced EHC is that the absence of Mrp3 in enterocytes is expected to result in more Res-3-G being secreted toward the intestinal lumen, in which it is available for deglucuronidation by the bacteria present in the large intestine. This should result in increased uptake of resveratrol and elevated plasma levels, not in reduced levels, as we have found. Fourth, Mrp3(-/-) mice have altered levels of an as-yet-unidentified transport protein capable of transporting unmodified resveratrol. Against this hypothesis argue our microarray data of WT and Mrp3(-/-) livers (data not shown), in which we did not find altered expression of transport proteins. Of course, this does not exclude the possibility that in other organs, including the gut, such alterations could be present. Finally, plasma levels of resveratrol decreased in the Mrp3(-/-) mouse, because it is mainly derived from deglucuronidation of Res-3-G, which is 10-fold lower in the Mrp3(-/-) mouse. This explanation is supported by several arguments. First, we find similar Res-3-G/resveratrol ratios in WT and Mrp3(-/-) mice, indicative of a similar percentage of circulating Res-3-G being deglucuronidated. Second, Bcrp1(-/-) mice do not show altered pharmacokinetics of unmodified resveratrol, even though BCRP/Bcrp1 readily transports resveratrol in vitro (Breedveld et al., 2007). This indicates that intracellular levels of unmodified resveratrol within enterocytes are too low for substantial amounts to be transported by Bcrp1 toward the gut lumen. This is in line with a study of Kuhnle et al. (Kuhnle et al., 2000) who have shown in Ussing chamber experiments that almost all resveratrol absorbed by the rat jejunum comes out basolaterally as resveratrol conjugated to glucuronic acid. We have determined whether Res-3-G is hydrolyzed in mouse plasma but found no significant resveratrol formation (data not shown). Hydrolysis might, however, occur intracellularly, followed by export of the resveratrol formed into the circulation by passive diffusion (Maier-Salamon et al., 2006) or via an as-yet-unidentified transporter. Intracellular deglucuronidation is not without precedent, because HepG2 cells can form quercetin from quercetin-glucuronide (O'Leary et al., 2003); moreover, diflunisal-glucuronides are deglucuronidated by rat liver during perfusion experiments (Brunelle and Verbeeck, 1997).
The fact that the major resveratrol metabolite, Res-3-G, is transported by BCRP and MRP3, is another illustration of the overlap in the substrate spectrum of BCRP and the members of the ABCC subfamily (Borst and Oude Elferink, 2002). MRP2 is probably a third player in the disposal of resveratrol, because we have found in vesicular uptake experiments that MRP2 transports Res-3-G as well (Supplemental Fig. 1). Indeed, experiments with Mrp2-deficient rats have shown that Mrp2 is the main transporter of Res-3-G into bile (Maier-Salamon et al., 2007). This overlap in substrate specificity of BCRP, MRP2, and MRP3 provides the cell with alternative excretion routes for the disposal of xenobiotics in case one or more transporters are blocked or overtaxed. We believe that under normal conditions, most Res-3-G formed in the enterocytes is transported by MRP3 with high affinity toward the circulation for urinary excretion. This will keep Res-3-G concentrations inside the enterocyte low and will prevent the transport of Res-3-G by BCRP or MRP2 over the apical membrane toward the lumen of the gut, in which the Res-3-G would be deglucuronidated by resident bacteria, thereby releasing resveratrol. This would result in a futile cycle of resveratrol/Res-3-G. When transport by MRP3 is hampered, however, BCRP and MRP2 might come into play transporting Res-3-G toward the lumen of the gut, thereby preventing the intracellular accumulation of Res-3-G.
The presence of Res-3-S and Res-di-S in urine samples after oral resveratrol administration implies that these metabolites are transported over the basolateral membrane of enterocytes and/or liver. The carriers responsible for this transport still need to be identified. Possible candidates that could do the job include MRP1/Mrp1 and MRP4/Mrp4, both of which transport sulfo-conjugates (Qian et al., 2001; Zelcer et al., 2003) and are present in enterocytes (Maher et al., 2005).
An intriguing observation was that Mrp3(-/-) mice develop diarrhea after oral administration of resveratrol, whereas WT mice and Bcrp1(-/-) mice do not (data not shown). The diarrhea in mice lacking Mrp3 does not seem to develop through the short-term effects of resveratrol on NaCl excretion or absorption, because we found no differences in Ussing chamber and ligated intestinal loop experiments between WT and Mrp3(-/-) mice (Supplementary Figs. 4 and 5). Although we currently have no explanation for the resveratrol-induced diarrhea in the absence of Mrp3, it might be related to the accumulation of resveratrol metabolites in colon and cecum that we detect in Mrp3(-/-) mice 24 h after resveratrol administration (data not shown).
In conclusion, our results show that the ABC transporters MRP3 and BCRP transport resveratrol metabolites in vitro and that their absence in mice results in large alterations of the pharmacokinetics of major resveratrol metabolites after an oral dose of resveratrol. It is now clear that several ABC transporters are involved in the tissue distribution and subsequent elimination of resveratrol from the body.
Acknowledgments
We thank Noam Zelcer (Howard Hughes Medical Institute, Department of Pathology and Laboratory Medicine, University of California, Los Angeles, CA), Alfred Schinkel (Division of Experimental Therapy, the Netherlands Cancer Institute), Ronald Oude Elferink (Department of Experimental Hepatology, University of Amsterdam) for critically reading this manuscript and Johanna Blitz for excellent biotechnical assistance.
Footnotes
-
This work was supported by the Nederlandse Organisatie voor Gezondheidsonderzoek en Zorginnovatie [TOP Grant 40-00812-98-07-028].
-
ABBREVIATIONS: MRP, multidrug resistance protein; ABC, ATP-binding cassette; BCRP, breast cancer resistance protein; DAD, diodearray; E217βG, estradiol-17β-d-glucuronide; EHC, enterohepatic circulation; FCS, fetal calf serum; HEK, human embryonic kidney; HPLC, high-pressure liquid chromatography; LSC, liquid scintillation counting; MTX, methotrexate; Res-3-G, resveratrol-3-glucuronide; Res-3-S, resveratrol-3-sulfate; Res-di-S, resveratrol-di-sulfate; Sf9, Spodoptera frugiperda 9; WT, wild type; TS buffer, Tris-HCl and sucrose.
-
↵
 The online version of this article (available at http://molpharm.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://molpharm.aspetjournals.org) contains supplemental material. - Received September 19, 2008.
- Accepted December 29, 2008.

- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}