


Abstract
We demonstrated recently that cytochrome b5 plays an important in vivo role in hepatic cytochrome P450 (P450) function [J Biol Chem 283:31385–31393, 2008]. We have now generated a model in which cytochrome b5 has been deleted in all tissues [cytochrome b5 complete null (BCN)], which surprisingly results in a viable mouse despite the putative in vivo roles of this protein in lipid and steroid hormone metabolism and the reduction of methemoglobin. In contrast to the liver-specific deletion, complete deletion of cytochrome b5 leads to a neonatal increase in the expression of many hepatic P450s at both the protein and mRNA level. In extrahepatic tissues, some changes in P450 expression were also observed that were isoform-dependent. In vitro cytochrome P450 activities in liver, kidney, lung, and small intestine of BCN mice were determined for a range of model substrates and probe drugs; a profound reduction in the metabolism of some substrates, particularly in lung, kidney, and small intestine, was observed. In vivo, the metabolism of metoprolol was significantly altered in BCN mice, in contrast to the previous finding in the liver-specific cytochrome b5 deletion, suggesting that extrahepatic cytochrome b5 plays a significant role in its disposition. Testicular Cyp17 hydroxylase and lyase activities were also significantly reduced by cytochrome b5 deletion, leading to significantly lower levels of testicular testosterone. The BCN mouse provides an additional model system with which to further investigate the functions of cytochrome b5, particularly in extrahepatic tissues.
Microsomal cytochrome b5 is a ubiquitous, 15.2-kDa hemoprotein associated primarily with the endoplasmic reticulum of eukaryotic cells. In conjunction with its primary electron donor cytochrome b5 reductase, this protein has been associated with many important cellular processes. Probably the most characterized is in the cytochrome P450 (P450)-mediated metabolism of xenobiotics and drugs (Porter, 2002; Yamazaki et al., 2002; Schenkman and Jansson, 2003; Yamaori et al., 2003; Zhang et al., 2005); and in cholesterol and steroid hormone homeostasis (Lamb et al., 2001; Akhtar et al., 2005). These effects on cytochrome P450 activity are very much substrate- and P450-specific, with evidence of both stimulation and inhibition of P450 turnover (Waskell et al., 1986; Porter, 2002; Schenkman and Jansson, 2003; Zhang et al., 2007, 2008). After almost 40 years of research, however, the mechanism of this interaction is still being described as “enigmatic” (Schenkman and Jansson, 2003) and “controversial” (Hildebrandt and Estabrook, 1971; Porter, 2002); and has been the focus of many, often contradictory, literature reports (Guryev et al., 2001; Yamazaki et al., 2002).
The other (non–P450-related) functions of cytochrome b5 include the reduction of heavy metals, carcinogenic arylhydroxylamines (Borthiry et al., 2007; Kurian et al., 2007), fatty acid desaturation, and methemoglobin/hemoglobin cycling (Jeffcoat et al., 1977; Umbreit, 2007). Furthermore, cytochrome b5 is also believed to play a role in the biosynthesis of plasmalogens, which are glycerol-based phospholipids, found in heart and neuronal tissues (Paltuaf et al., 1974; Woelk and Jahrreiss, 1978; Lee et al., 1991).
To establish the in vivo functions of this protein, we have generated recently a mouse in which microsomal cytochrome b5 was conditionally deleted in the liver (HBN mice) (Finn et al., 2008). HBN mice had significantly reduced in vitro rates of hepatic NADPH- and NADH-dependent metabolism of a range of model P450 substrates and probe drugs; furthermore, significant changes in drug pharmacokinetics occurred on administration of a probe drug cocktail, demonstrating that hepatic cytochrome b5 can play a significant role in in vivo drug disposition.
We have now generated a mouse model in which the cytochrome b5 has been deleted in all mouse tissues [b5 complete null (BCN)]. BCN mice were viable and fertile, and in this report, we describe the effects of cytochrome b5 deletion on the expression of drug metabolizing enzymes and on hepatic and extrahepatic foreign compound metabolism.
Materials and Methods
Chemicals.
All reagents unless stated were purchased from Sigma-Aldrich (Poole, UK). NADPH was obtained from Melford Laboratories (Ipswich, UK). 7-Benzyloxy-4-trifluoromethylcoumarin (BFC), 7-methoxy-4-trifluoromethylcoumarin (MFC), 7-hydroxy-4-trifluoromethylcoumarin, and hydroxy-tolbutamide were purchased from BD Gentest (Cowley, UK). Midazolam, 1-hydroxy-midazolam, and 4-hydroxy-midazolam were kind gifts from Roche Diagnostics, (Mannheim, Germany), and 1-hydroxy metoprolol and O-desmethyl metoprolol were generous gifts from Astra Häsle (Mölndal, Sweden). PCR primers were obtained from Eurofins MWG Operon (Ebersberg, Germany).
Generation of BCN Mice.
Cytb5lox/lox mice were generated as described previously (Finn et al., 2008) and maintained by random breeding on a 129P2 × C57BL/6 genetic background. Cytb5lox/lox mice were crossed with a transgenic mouse line expressing Cre recombinase under the control of a phosphoglycerate kinase promoter (CrePGK) (Lallemand et al., 1998) on a C57BL/6 background, and Cytb5(+/−)::CrePGK offspring were backcrossed with Cytb5(+/+) mice to generate heterozygous cytochrome b5 complete knockout mice minus CrePGK [Cytb5(+/−)], which were subsequently crossed to generate homozygous cytochrome b5 complete knockout mice [BCN, Cytb5(−/−)] and control [wild-type, Cytb5(+/+)] mice. The BCN line was thereafter maintained using a combination of crosses: Cytb5(+/−) females with Cytb5(−/−) males; Cytb5(−/−) females with Cytb5(+/−) males; and Cytb5(−/−) females with Cytb5(−/−) males. Mice carrying the null cytochrome b5 allele were identified by multiplex PCR using the following primer set: wild-type forward primer, 5′-TCCCCCTGAGAACGTAATTG-3′; null forward primer, 5′-GGTCTCTCCTTGGTCCACAC-3′; and common reverse primer, 5′-GAGTCTTCGTCAGTGCGTGA-3′ (Supplemental Fig. S1). The presence of the CrePGK transgene was determined as described previously (Henderson et al., 2003). All mice were maintained on a standard chow (RM1; Special Diet Services, Essex, UK) under standard animal house conditions, with free access to food and water and a 12-h light/dark cycle. All animal work was carried out on 10-week-old male mice, except where indicated, in accordance with the Animal Scientific Procedures Act (1986) and after local ethical review.
Preparation of Microsomes.
Microsomes were prepared from wild-type and BCN mouse tissues, using 0.3 to 0.5g of tissue, by the method of Meehan et al. (1988) for the small intestine and by a modified method of this method for all other tissues using sonication instead of mechanical homogenization (Pritchard et al., 1998). Microsomal protein concentrations were determined using the Bio-Rad Protein Assay Reagent (Bio-Rad Laboratories, Hemel Hempstead, Herts, UK). P450 oxidoreductase (POR) activity was estimated by NADPH-dependent cytochrome c reduction (Strobel and Dignam, 1978). Microsomes were stored at −70°C until required.
Quantitation of Microsomal Cytochrome b5.
The specific amount (measured in picomoles of cytochrome b5 per milligram of microsomal protein) of cytochrome b5 expressed in various tissues was determined by NADH-reduced spectrophotometry as follows: a known volume of microsomes was resuspended in 2 ml of 100 mM potassium phosphate buffer, pH 7.4, and the was suspension split between two matched quartz cuvettes. An absorbance baseline was plotted between 400 and 500 nm, after which NADH was added to the sample cuvette to a concentration of 200 μM, and a reduced spectrum was determined. The amount of cytochrome b5 in the sample was calculated by determining the absorbance difference between 426 and 409 nm using an extinction coefficient of 185,000 l/mol. Data are the mean of three animals (per tissue) ± S.D.
Immunoblotting.
Immunoblot analysis was carried our as described previously (Finn et al., 2008). Cyp17 antibody was a kind gift from Professor Mike R. Waterman (Vanderbilt University, Nashville, TN). Immunoreactive proteins were detected using polyclonal goat anti-rabbit, anti-mouse, or anti-sheep horseradish peroxidase immunoglobulins as secondary antibodies (Dako UK Ltd., Ely, UK) and visualized using Immobilon chemiluminescent horseradish peroxidase substrate (Millipore, Watford, UK) and a Fujifilm LAS-3000 mini-imaging system (Fujifilm UK Ltd., Bedfordshire, UK). Densitometric analysis was performed using MultiGauge version 2.2 software (Fujifilm UK Ltd.).
RNA Isolation and Real-Time Quantitative PCR Analysis.
Total RNA was isolated from snap-frozen liver samples using TRIzol (Invitrogen, Paisley, UK). Genomic DNA was removed using RQ1 DNase (Promega, Southampton, UK), and RNA (600 ng) was reverse-transcribed using random hexamers and Superscript II RNase H reverse transcriptase polymerase (Invitrogen). Real-Time quantitative PCR analysis was carried out with optimized gene-specific primer sets (Applied Biosystems, Warrington, UK), except in the case of cytochrome b5, in which the following primer-probe set was used: forward primer, 5′-GGGTGATGCTACCGAGAATTTT-3′; reverse primer, 5′-AGTTCCCCGATGATGTATGTTTTG-3′; and probe, [6-carboxyfluorescein]-CGTCGGGCACTCTACGGATGCAC-[5-carboxytetramethylrhodamine/ 6-carboxyfluorescein].
Reactions were carried out in triplicate and monitored by measuring fluorescence at 518 nm with excitation at 494 nm. mRNA levels were quantified using the Prism 7700 associated software and normalized to the levels 18s mRNA (Applied Biosystems).
P450 Spectral Analysis.
P450 content of mouse liver microsomes was determined by reduced CO difference spectroscopy for three samples of each genotype using the method described by Omura and Sato (1964).
Generic P450 Microsomal Incubations.
Microsomal incubations were carried out in triplicate in 50 mM HEPES, pH 7.4, and 30 mM MgCl2 containing mouse tissue microsomes and substrate prewarmed to 37°C before initiation of reaction by the addition of either NADPH or NADH to a final concentration of 0.67 mM.
Fluorogenic Assay Incubations.
Assays were performed essentially as described previously (Finn et al., 2008) using the following substrate and microsome concentrations: BFC, 40 μM substrate and 20 μg of mouse tissue microsomes; EFC, 40 μM substrate and 20 μg of mouse liver microsomes; MFC, 140 μM substrate and 20 μg of mouse liver microsomes; and ER, BR, MR, and PR, 1 μM substrate and 20 μg of mouse liver microsomes. Reactions were measured in real time for 3 min at the recommended excitation and emission wavelengths for each probe using a Fluroskan Ascent FL plate-reading fluorimeter (Thermo Fisher Scientific, Waltham, MA). Turnover rates were calculated using authentic metabolite standards (7-hydroxy-4-trifluoromethylcoumarin for BFC, EFC and MFC assays and resorufin for BR, ER, MR, and PR assays).
Recapitulation of P450 Activity.
BCN liver microsomes (20 μg) were mixed with increasing amounts of Escherichia coli membranes containing recombinant murine cytochrome b5 (3.3, 6.6, 13.2, and 26.4 pmol of cytochrome b5). Tubes were capped and incubated at room temperature for 20 min. Potassium phosphate buffer (190 μl of 100 mM, pH 7.4) containing 30 μM BFC or 0.8 μM BR was then added, and tubes were mixed before adding into the wells of a 96-well plate. Reactions were initiated with 10 μl of 10 mM NADPH and run for 3 to 5 min, and metabolites were measured and quantified as described previously. Cytochrome b5, wild-type, and wild-type plus cytochrome b5 controls were also included. Experiments using lung microsomes were carried out in the same manner except that a single concentration of cytochrome b5 was used (26.4 pmol).
BFC and MR Kinetics.
Assays to determine the apparent kinetic parameters were performed in triplicate with wild-type and HBN liver microsomes under conditions of linearity for time and protein (data not shown) using the same buffer/NADPH conditions as described above with the following concentrations of substrates: MR, 0.002 to 1 μM; and BFC, 0.01 to 100 μM, using 10 and 11 concentration points, respectively.
LC/MS-MS Assay Incubations.
Incubations were performed under the following conditions: chlorzoxazone, 1 mM substrate and 20 μg of microsomes in a final volume of 150 μl for 30 min; midazolam, 50 μM substrate and 20 μg of microsomes in a final volume of 100 μl for 9 min; metoprolol, 800 μM substrate and 30 μg of microsomes in a final volume of 100 μl for 60 min; phenacetin, 50 μM substrate and 20 μg of microsomes in a final volume of 100 μl for 9 min; tolbutamide, 800 μM substrate and 30 μg of microsomes in a final volume of 10 μl for 60 min. Assays were stopped by the addition of either 0.5 volumes (chlorzoxazone) or 1 volume (tolbutamide, midazolam, metoprolol, and phenacetin) of ice-cold methanol and incubated on ice for 10 min. Samples were centrifuged for 8 min at 16,000g to remove particulate material before HPLC (chlorzoxazone) or LC/MS-MS (tolbutamide, midazolam, metoprolol, and phenacetin) analysis was carried out as described previously (Finn et al., 2008).
Testicular Testosterone Content.
Testosterone was extracted from the testes of 10-week-old mice using the method of Zhang et al. (2003) and analyzed by reversed-phase HPLC (Paine et al., 2003).
Progesterone and 17α-Hydroxyprogesterone Incubations.
Incubations were performed in triplicate in a final volume of 150 μl of 50 mM potassium phosphate buffer, pH 7.4, containing 25 μg of testicular microsomes and either 8 μM progesterone or 15 μM 17α-hydroxyprogesterone. Reactions were incubated at 37°C and were initiated by the addition of NADPH. After 20 min, incubations were stopped by the addition of 75 μl of ice-cold methanol. Samples were incubated on ice for 10 min before being centrifuged for 8 min at 16,000g to remove particulate material before HPLC analysis.
Progesterone and 17α-Hydroxyprogesterone Assay HPLC Conditions.
Substrates and metabolites (progesterone, 17α-hydroxyprogesterone, androstenedione, and testosterone) were separated by reversed-phase HPLC using a Hypersil ODS column (5 μm; 4 × 250 mm; Agilent, Stockport, UK) maintained at 35°C with the following mobile phase conditions: 1 to 3 min, 2.5% acetonitrile/41% water/56.5% methanol; and 3 to 9 min, a linear gradient was applied to reach 5% acetonitrile/25% water/72.5% methanol. This was maintained for a further 2 min before applying a linear gradient for 50 s to reach the original solvent concentrations, which were maintained for a further 3 min. Metabolites were detected at 240 nm and quantified using authentic standards.
Statistical Methods.
Average rates of metabolism were calculated for each triplicate incubation of mouse liver microsomes from each genotype (n = 3), and these data were then used to calculate p values using an unpaired t test.
In Vitro Pharmacokinetics of Metoprolol.
Five 10-week-old male mice of each genotype were orally gavaged with metoprolol (2 mg/kg). Blood samples were taken, and the samples were analyzed by LC/MS-MS as described previously (Finn et al., 2008). Pharmacokinetic parameters were calculated using a noncompartmental model (WinNonLin version 4.1; Pharsight, Munich, Germany).
Results
Phenotype of BCN Mice.
To establish the in vivo role(s) of microsomal cytochrome b5, we generated a global deletion of this gene in the mouse. Mice lacking microsomal cytochrome b5 were unexpectedly viable, born at expected Mendelian ratios, and exhibited no gross anatomical abnormalities. BCN mice of both sexes were fertile, indicating that cytochrome b5 was not necessary for sexual maturation. The BCN mice, however, did display dry skin and an interesting fur condition in which pups and young adult mice had a characteristically dull, sparse coat, with marked piloerection, along with a degenerative nurturing phenotype probably related to changes in milk composition. This phenotype was reflected in low neonatal body weights, which improved when the pups were weaned and became normal when the animals reached 6 to 8 weeks of age. These characteristics are described in more detail elsewhere (Finn et al., 2010).
Analysis of Cytochrome P450-Dependent Mono-oxygenase Components.
The cytochrome b5 content was measured in wild-type liver (253.7 ± 47.9 pmol/mg) (Fig. 1), kidney (208.4 ± 15.1pmol/mg), and small intestine microsomes (161.4 ± 16.8 pmol/mg). Cytochrome b5 levels were not detectable in the equivalent BCN samples. Total hepatic cytochrome P450 levels were significantly increased (2-fold) in the BCN mice as measured by Fe2+-CO versus Fe2+ difference spectra (0.4 ± 0.07 versus 0.9 ± 0.4 nmol P450/mg microsomal protein; p < 0.05) (Fig. 2A). To confirm that cytochrome b5 protein had been deleted, microsomes from liver, lung, kidney, small intestine, and testes from 10-week-old male mice were analyzed by Western blotting. Cytochrome b5 protein was undetectable in all tissues of the BCN mice studied (Fig. 2, B and D). To explain the increase in P450 content, we carried out Western blotting analysis to establish which P450 isoenzymes were changed; microsomes of each genotype were analyzed for the expression of P450s, POR, and cytochrome b5 reductase (Fig. 1, B and C). It is noteworthy that hepatic P450 expression was increased in BCN for members of the Cyp2a, Cyp2b, Cyp2e, and Cyp3a subfamily members, with Cyp2b10 showing the most pronounced induction (17-fold; p < 0.001) (Fig. 2, B and C). BCN hepatic POR levels showed a small (25%) but statistically significant increase, which was reflected in an increase in the rate of reduction of the POR substrate cytochrome c compared with wild-type (130.2 ± 9.7 versus. 74.8 ± 17.3 nmol cytochrome c reduced/min/mg, respectively; p = 0.008). The increase in cytochrome c reduction was greater than the increase in POR protein. The reason for this is unclear.
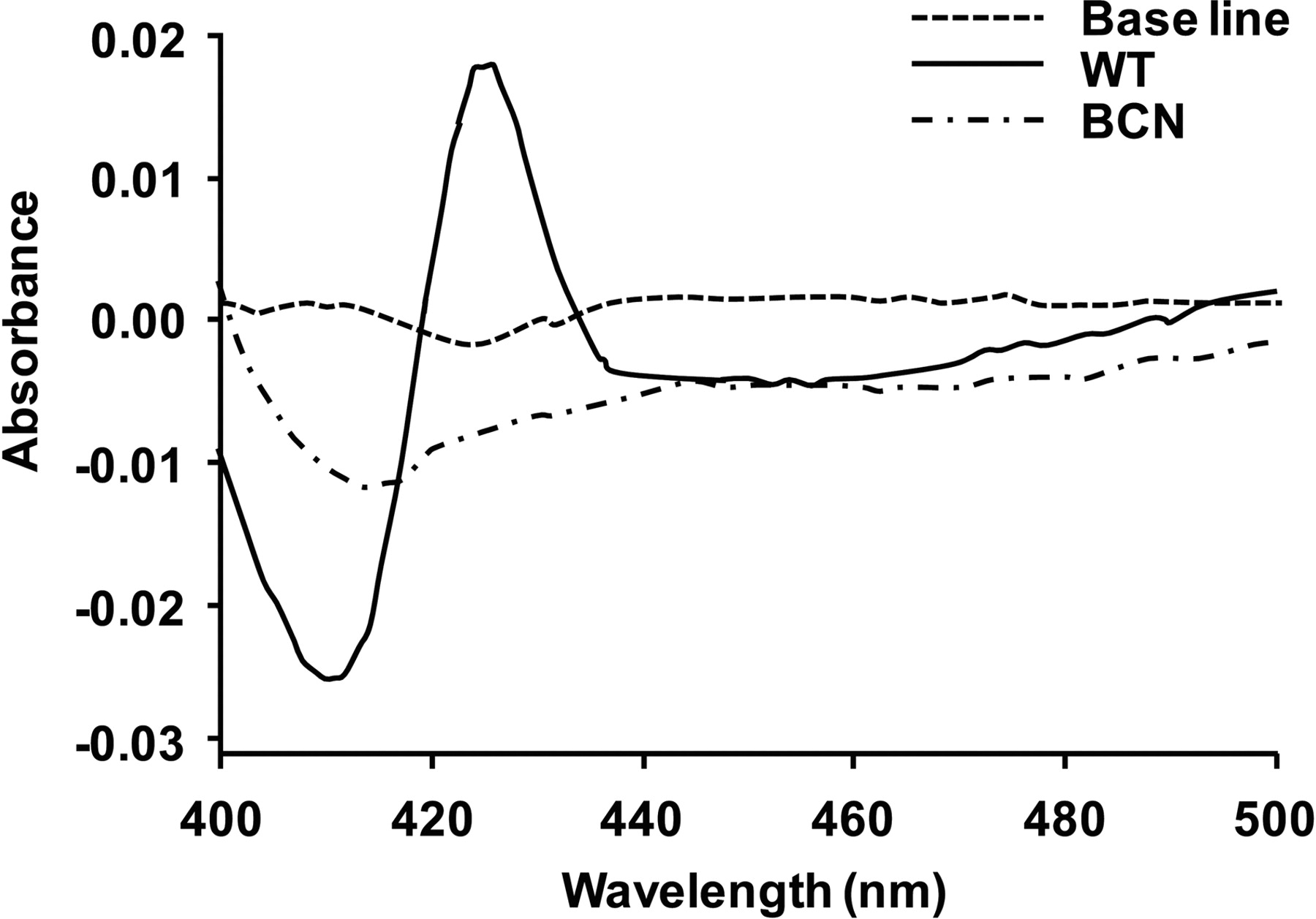
Quantitation of microsomal cytochrome b5 by difference spectrophotometry. Traces illustrate typical cytochrome b5 difference spectra obtained using hepatic microsomal fractions from Cytb5(+/+) (WT) and Cytb5(−/−) (BCN) mice. Assays were performed as described under Materials and Methods.

Characterization of cytochrome P450 mono-oxygenase components in cytochrome b5-null mice. A, CO difference spectra in hepatic microsomal fractions of Cytb5(+/+) (WT) (solid line) and Cytb5(−/−) (BCN) (dashed line) hepatic microsomes. Samples were run as described under Materials and Methods. B, immunoblot analysis of liver, kidney, lung, and small intestine microsomes (20 μg protein/lane) from Cytb5(+/+) (WT) and Cytb5(−/−) (BCN) mice, as detailed under Materials and Methods. C, densitometric analysis of liver immunoblot data (from B). Values are expressed as the fold change normalized to wild-type expression. Bars are mean ± S.D. from three samples of each genotype. ▨, wild-type values; □, BCN values. D, quantification of hepatic mRNA changes by Taqman analysis. Data are normalized to 18S expression and expressed as the fold change relative to wild-type levels. Bars are mean ± S.D. from three samples of each genotype. ▨, Cytb5(+/+) (wild type); □, Cytb5(−/−) (BCN) values. E, immunoblot analysis of cytochrome P450-dependent mono-oxygenase components in liver microsomes (20 μg protein/lane) from 1-day-old Cytb5(+/+) (WT) and Cytb5(−/−) (BCN) pups as detailed under Materials and Methods. *, p ≤ 0.05, **, p ≤ 0.01; ***, p ≤ 0.001.
In extrahepatic tissues, some changes in the levels of cytochrome P450 expression were observed: namely, an 80% decrease in the expression of Cyp2e1 was observed in the kidney (p = 0.035), a 35% increase in the expression of Cyp2b (p = 0.004) in the lung was observed, and in the small intestine we saw a 70% decrease (p = 0.003) in Cyp2d and a 40% decrease in Cyp2e (p = 0.03) (Fig. 2B). No changes in testicular P450 levels were observed (data not shown). The increases in hepatic cytochrome P450 expression were reflected in the mRNA levels for Cyp2a4, Cyp2b10, Cyp3a11, and Cyp4a10 (Fig. 2D), indicating that the observed increases in protein expression were due to increases in gene transcription or mRNA stabilization. To determine whether the changes in P450 expression observed in BCN liver were present from birth, hepatic P450 levels were measured in 1-day-old pups (two from each genotype). As in the adult mice, Cyp2a, Cyp2b, and Cyp3a levels were increased in neonatal BCN liver at this early time point, whereas Cyp2c and Cyp2d protein was marginally repressed (Fig. 2E).
In Vitro Cytochrome P450 Activities.
In vitro cytochrome P450 activities in microsomes from kidney, liver, lung, and small intestine of wild-type and BCN mice were determined using probe substrates comprising alkoxy-resorufin and alkoxy-4-trifluoromethylcoumarin derivatives (Table 1). In the case of the alkoxy-resorufins, no differences in the hepatic metabolism of ER or MR between BCN and wild-type mice were observed, whereas the BCN samples exhibited a significantly higher rate of metabolism of BR (4.1-fold) and PR (2.4-fold). These increases in turnover of PB and BR probably reflect the marked increase of Cyp2b10 expression, which is known to metabolize these compounds. Hepatic alkoxy-4-trifluoromethylcoumarin metabolism was significantly lower in the BCN samples (BFC, 4.2-fold; EFC, 5.5-fold; and MFC, 5.5-fold lower, respectively). The changes in hepatic metabolism of MR and BFC were also reflected in the kinetic parameters (Table 2). All data were monophasic and followed standard Michaelis-Menten kinetics, and in the case of MR metabolism, there was little difference in either Km or Vmax between wild-type and BCN samples. However, deletion of cytochrome b5 caused a 4.5-fold increase in Km and a 4.6-fold decrease in Vmax for the metabolism of BFC.
Effect of cytochrome b5 deletion on cytochrome P450 activities
For liver and small intestine samples, assays were performed in triplicate on three individual mouse samples, and data shown are mean ± S.D. for three mice. Because of the poor yield of microsomes from mouse lung, assays were performed in triplicate on a pool of three lung preparations in which data presented are mean ± S.D. of the triplicate determinations, precluding statistical analysis. The percentage change is shown in brackets for the BCN samples. Kidney microsomes were also tested, but no activities were detected. For BR, ER, MR, and PR, the metabolite formed is resorufin, and for BFC, EFC, and MFC, it is 7-hydroxy-4-trifluoromethylcoumarin.
Kinetic parameters for hepatic microsomal methoxyresorufin (MR) and 7-benzyloxy-4-trifluoromethylcoumarin (BFC) metabolism in Cytb5(+/+) (wild-type) and Cytb5(−/−) (BCN) liver microsomes
Assays were performed using triplicate samples for each concentration of substrate as described under Materials and Methods. Standard deviations given are from the fit of the curve as calculated using the Michaelis-Menten equation (GraFit version 5; Erithacus Software, Horley, UK).
In the lung, metabolism of the alkoxy-resorufins was reduced to undetectable levels in the BCN samples, and with the three 4-trifluoromethylcoumarins, metabolism was also profoundly reduced (BFC, 29.8-fold; EFC, 5.5-fold; and MFC, 115-fold lower). In the small intestine, BR, PR, BFC, EFC, and MFC metabolism was detected in wild-type mice, but in BCN mice, metabolism of BR and BFC was markedly reduced, and the metabolism of EFC, MFC, MR, and ER was reduced to undetectable levels. In contrast, the rate of PR metabolism in the intestine was increased in the BCN samples. No activity toward these probe substrates could be measured using either wild-type or BCN kidney microsomes. In liver microsomes from wild-type and BCN mice, NADH-dependent metabolism of BFC, EFC, and MFC was approximately 25 to 30% of that of the equivalent NADPH reactions in wild-type mice (BFC, 57 versus 275 pmol/min/mg; EFC, 61 versus 245 pmol/min/mg; and MFC, 55 versus 181 pmol/min/mg). However, no NADH-dependent metabolism could be detected in the BCN.
Effect of the Addition of Cytochrome b5.
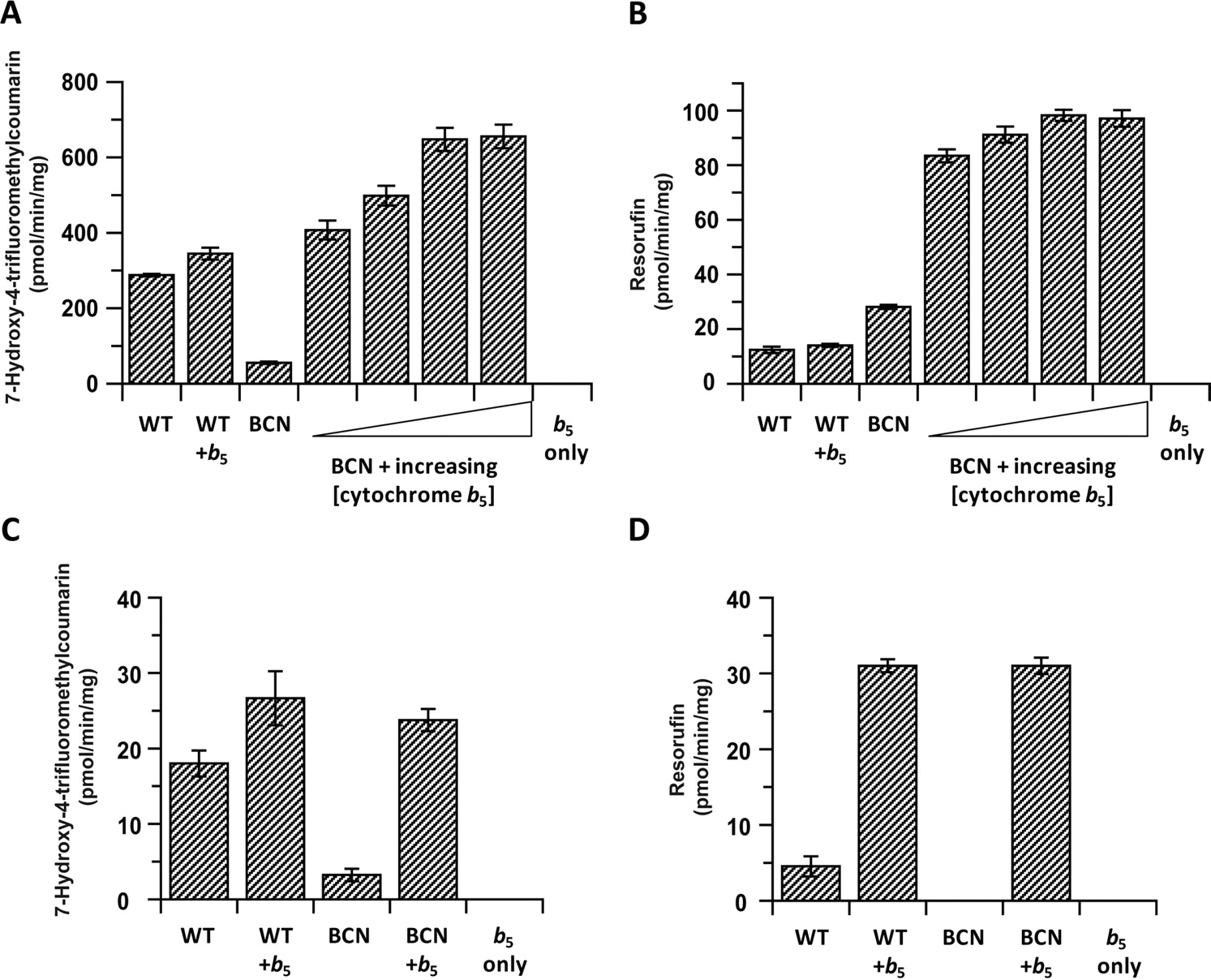
To establish whether the changes in mono-oxygenase activity could be directly attributable to the absence of cytochrome b5, E. coli membranes containing recombinant murine cytochrome b5 were added back to BCN liver microsomes. This caused a concentration-dependent increase in BFC activities, with the highest concentration of added cytochrome b5 giving approximately a 12-fold greater turnover than BCN microsomes alone and a 2.3-fold increase greater than wild-type levels (Fig. 3A). The addition of the highest concentration of cytochrome b5 to wild-type microsomes caused a comparatively small (19%) increase in activity. When this experiment was carried out using BR as a substrate (Fig. 3B), no increase in wild-type activity was observed; however, the activity in BCN microsomes (which was already approximately 2-fold more active toward BR than wild-type) was increased by 4.5-fold by cytochrome b5 addition, achieving a 7.8-fold increase over the control levels. When cytochrome b5 was added to wild-type lung microsomal incubations, activities were increased 1.5- and 7-fold for BFC and BR, respectively (Fig. 3, C and D). The addition of the same amount of cytochrome b5 back to BCN lung microsomes resulted in a 7.4- fold increase in BFC turnover (Fig. 3C) and the appearance of BR activity (Fig. 3D). For both substrates, these activities were equal to those observed when cytochrome b5 was added back to wild-type membranes. Similar experiments were performed using kidney microsomes with chlorzoxazone as substrate; in this case, the addition of cytochrome b5 to BCN microsomes brought the activity back approximately to the levels seen in wild-type microsomes, whereas it did not increase the activities of wild-type microsomes (Rates, in picomoles per minute per milligram, were the following: WT, 110 ± 5; BCN, 14 ± 1; WT + cytochrome b5, 96 ± 1; and BCN + cytochrome b5, 91 ± 2).

The effect of exogenous cytochrome b5 addition on microsomal cytochrome P450 activities. A and B, activities of Cytb5(+/+) (WT) and Cytb5(−/−) (BCN) liver microsomes toward 7-benzyloxy-4-trifluoromethylcoumarin (A) and benzyloxy-resorufin (B) in the absence and presence of increasing concentrations of recombinant cytochrome b5 (3.3, 6.6, 13.2, and 26.4 pmol of cytochrome b5 per incubation, respectively). Samples were treated as detailed under Materials and Methods, and values are presented as mean ± S.D. from triplicate incubations. C and D, activities of Cytb5(+/+) (WT) and Cytb5(−/−) (BCN) lung microsomes toward 7-benzyloxy-4-trifluoromethylcoumarin (C) and benzyloxy-resorufin (D) in the absence and presence of recombinant cytochrome b5 (26.4 pmol/incubation). Samples were treated as described under Materials and Methods, and data are presented as the mean ± S.D. from triplicate incubations.
Metabolism of Probe Drugs.
In addition to the use of model substrates, in vitro assays were performed with probe drug substrates (Fig. 4). Chlorzoxazone 6-hydroxylation was significantly reduced in liver, lung, and kidney by 4-, 2-, and 5.5-fold, respectively (Fig. 4A). In the case of midazolam, the rate of production of the 1′- and 4-hydroxy-midazolam metabolites was also much lower in the BCN samples, reaching statistical significance in the liver and kidney for 1′-hydroxy-midazolam production and in the liver for midazolam 4-hydroxylation (Fig. 4B). Metoprolol O-demethylation was significantly reduced in cytochrome b5-null liver and kidney samples, whereas no effect was observed in the lung or small intestine samples. 1′-Hydroxy-metoprolol was detected in liver, kidney, and small intestine incubations but not in the lung samples. Similarly to O-desmethyl metoprolol production, formation of this metabolite was significantly decreased in the liver and kidney samples of BCN mice (Fig. 4C). In contrast to the above substrates, metabolism of phenacetin to acetaminophen was significantly increased in the livers of BCN mice (p < 0.05), whereas in the kidney and lung, ablation of cytochrome b5 had no effect on activity (Fig. 4D). In the case of tolbutamide hydroxylation, activity was only detected in liver and kidney microsomes; however, in both cases, BCN samples showed lower activity compared with controls. The reduction was significant in the kidney (Fig. 4E).

Probe drug metabolism in microsomes from wild-type and cytochrome b5-null mice. Assays were performed in triplicate on microsomes from three mice of each genotype, as described under Materials and Methods. A, chlorzoxazone 6-hydroxylase; B, midazolam 1′- (1′-OH Midaz) and 4-hydroxylase (4-OH Midaz); C, metoprolol O-demethylase (O-DM Metop) and 1-hydroxylase (1-OH Metop); D, phenacetin O-deethylase; and E, tolbutamide hydroxylase activities. ▨, Cytb5(+/+) (wild-type); □, Cytb5(−/−) (BCN) samples. Mean values are presented ± S.D. Lv, liver; Ln, lung; K, kidney; SI, small intestine. *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001.
In Vivo Metoprolol Pharmacokinetics.
To determine whether the observed decreases in in vitro cytochrome P450 metabolism were reflected in vivo, the pharmacokinetics of orally administered metoprolol were determined in wild-type and BCN mice. The elimination profiles of metoprolol disappearance were significantly different between wild-type and BCN mice (Fig. 5). The area under the curve and terminal half-life of metoprolol were significantly increased (3.5- and 2.4-fold, respectively) and clearance decreased (3.5-fold) in the BCN animals compared with wild type. Furthermore, although not statistically significant, the metoprolol Cmax values were also increased in the BCN (4-fold). Further studies have been carried out that show a clear effect of cytochrome b5 deletion on the pharmacokinetics of a range of drugs. These data will form the basis of a further article.

In vivo pharmacokinetic profiles and parameters of the cytochrome P450 substrate metoprolol in wild-type and cytochrome b5-null mice. Metoprolol was administered orally to Cytb5(+/+) (wild-type) and Cytb5(−/−) (BCN) mice at a dose of 2 mg/kg. The data shown represent the mean ± S.E.M. of metoprolol blood concentrations at the individual time points with five animals per group. ●black circles;○, BCN. Samples were analyzed, and pharmacokinetic parameters were calculated as described under Materials and Methods. *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001.
Steroid Hormone Metabolism and Testosterone Levels.
Cytochrome b5 has been reported to be essential for the conversion of 17α-hydroxyprogesterone to androstenedione (Akhtar et al., 2005). To study the pathway of testosterone biosynthesis, we measured progesterone metabolism using testicular microsomes from wild-type and BCN mice. A significant 2-fold reduction in the rate of metabolism was measured in the absence of microsomal cytochrome b5 (Fig. 6A). When 17α-hydroxyprogesterone was used as a substrate to investigate the effect of cytochrome b5 deletion on Cyp17-lyase activities, testicular BCN microsomes had almost no activity, being 100-fold lower than wild-type controls (p < 0.001) (Fig. 6A). Because Cyp17 and 17β-hydroxysteroid dehydrogenase are both microsomal NADPH-dependent enzymes (Fig. 6C), incubations with 17α-hydroxyprogesterone also yielded the metabolite testosterone (produced by the metabolism of androstenedione by 17β-hydroxysteroid dehydrogenase) (Fig. 6C). Testicular microsomes from wild-type mice were 66-fold more active than BCN in the production of testosterone from androstenedione formed by the Cyp17 (Fig. 6A). A reflection of this reduction in levels of testosterone was also observed in vivo, because when testes from wild-type and BCN mice were analyzed for testosterone content, BCN samples contained significantly less testosterone than wild-type (approximately 1.8-fold) (Fig. 6B). There were no compensatory alterations in the levels of Cyp17 protein expressed in the testes of cytochrome b5-null mice (Fig. 6D).

The effect of cytochrome b5 deletion on steroid hormone metabolism. A, assays were performed in triplicate using testicular microsomes from three mice of each genotype as described under Materials and Methods. Activities were determined using either progesterone (Prog) or 17α-hydroxy progesterone (17α OH Prog) as substrates. ▨, Cytb5(+/+) (WT); □, Cytb5(−/−) (BCN) represent the respective levels of metabolite production (Andro, androstenedione; Test, testosterone). Each bar represents mean ± S.D. *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001. B, intratesticular testosterone content of Cytb5(+/+) (wild-type) (▨) and Cytb5(−/−) (BCN) (□) mice. Bars represent the respective levels (mean ± S.D.) for three mice of each genotype expressed per milligram of weight of tissue. C, pathway of NADPH-dependent testosterone formation from progesterone. D, analysis of cytochrome b5 and Cyp17 levels by Western blotting (20 μg protein/lane) from Cytb5(+/+) (WT) and Cytb5(−/−) (BCN), for the expression of Cyp17.
Discussion
Because of the large number of functions associated with cytochrome b5, which include fundamental processes such as steroid hormone biosynthesis and hemoglobin homeostasis (Porter, 2002; Schenkman and Jansson, 2003), it would be predicted that the deletion of this enzyme could be lethal. Unexpectedly, this was not the case, with BCN pups produced at expected Mendelian ratios with no gross anatomical abnormalities. In addition, no overt reduction in fertility was observed in either gender. Several phenotypic characteristics were identified, however, including skin defects closely resembling the human condition ichthyosis vulgaris, which were exacerbated in breeding females on the birth of successive litters.
Until the publication of our recent article detailing the conditional hepatic deletion of microsomal cytochrome b5 (Finn et al., 2008), investigation of the role of cytochrome b5 in drug metabolism was based on in vitro studies (Gruenke et al., 1995; Yamazaki et al., 1996; Yamazaki and Shimada, 2006) or heterologous overexpression of proteins (Patten and Koch, 1995). The BCN model therefore allows the role of cytochrome b5 in the in vitro and in vivo metabolism of drugs to be established.
Complete deletion of cytochrome b5 as evidenced by spectral analysis (Fig. 1) and Western blotting (Fig. 2B) resulted in significant increase in hepatic expression of Cyp2a, Cyp2b, and Cyp3a proteins. The presence of multiple bands in the Western blots for Cyp2a, Cyp2b, Cyp2c, and Cyp4a reflect the multiplicity of the proteins within each gene subfamily in the mouse, and it seems that cytochrome b5 deletion differentially affects the expression of these multiple forms. Which specific members of these gene subfamilies are up-regulated is the subject of ongoing investigations. The increases in cytochrome P450 content were already apparent from birth and were mirrored by increases in mRNA expression, suggesting a transcriptional mechanism. A similar increase in P450 expression was also observed when hepatic POR was conditionally deleted (Gu et al., 2003; Henderson et al., 2003), which in this latter case seems to be due to activation of the constitutive androstane receptor and, to a lesser degree, the pregnane X receptor by accumulated polyunsaturated fatty acids (Finn et al., 2009). No such increases were found in BCN mice, which, unlike the HRN mice, did not have a fatty liver, indicating that the activation is the result of an alternative metabolic pathway.
Investigation of the in vitro metabolism of cytochrome P450 substrates produced some unexpected and interesting results (Table 1). NADH-mediated metabolism was undetectable in BCN mice, demonstrating that cytochrome b5 is an unequivocal electron donor to cytochrome P450s. For NADPH-dependent metabolism, BCN hepatic microsomes exhibited the same magnitude of decrease in the metabolism of the alkoxy-trifluoromethylcoumarins as observed previously in HBN mice (Finn et al., 2008); however, there were some differences between these models with alkoxy-resorufin metabolism. Turnover of MR and ER in BCN hepatic microsomes was essentially unchanged from the rates observed in wild type, whereas in the HBN microsomes, ER O-deethylase activity was approximately 26% lower (Finn et al., 2008). This difference could be explained by the finding that compensatory changes in cytochrome P450 gene expression in BCN mice were not observed in HBN animals; therefore, the ratio of different P450s to each other and their absolute levels is different between the lines. These data suggest that cytochrome b5 plays little role in the metabolism of Cyp1a substrates. In support of this, the metabolism of BR and PR, traditionally attributed to Cyp2b enzymes, was increased 4.1- and 2.4-fold, respectively, correlating very well with the markedly higher levels of hepatic Cyp2b expression in BCN samples (Fig. 2, B and C). In the HBN model, BR was metabolized significantly slower (30%) of the wild-type rate (Finn et al., 2008). Indeed, Cyp2b10 levels were elevated 17-fold in BCN liver, which would therefore equate to a predicted maximal 5.1-fold increase in turnover (17 × 30% activity), close to the figure in this study (4.1-fold). The above data are interesting in that in the conditional HBN model, the effects of cytochrome b5 deletion are reflected in changes in the activities of constitutively expressed cytochrome P450 enzymes, whereas in the BCN mice, the consequence of this deletion are observed in both constitutive and inducible enzymes. It will be interesting to establish the impact of cytochrome b5 deletion after pretreatment with P450 inducers such as phenobarbital, in which it may be possible to normalize the level of P450 expression.
The activities of BCN lung microsomes toward the fluorogenic probes were more markedly reduced by cytochrome b5 deletion than the hepatic samples, showing either a complete ablation or profound reduction in turnover. Cytochrome b5 therefore seems to differentially affect the activity of cytochrome P450s in different tissues. Adding back exogenous cytochrome b5 to hepatic and pulmonary BCN incubations restored turnover to wild-type levels and then further stimulated activities in a concentration-dependent manner for both BR and BFC activity, resulting in activities 2.3- and 7.8-fold higher than wild type for BFC and BR, respectively. This “superactivation” of turnover can be attributed to the increased levels of Cyp2b (BR and BFC activity) and Cyp2a and Cyp3a (BFC) in the livers of BCN animals (Fig. 2, B and C). Furthermore, the addition of exogenous cytochrome b5 to wild-type lung microsomes, unlike hepatic microsomes, significantly stimulated turnover, most markedly during BR O-dealkylation (6.8-fold). Together, these data suggest that, in the lung, cytochrome b5 is a key component of the cytochrome P450 mono-oxygenase system (Croft et al., 1986), the constitutive level of which for the metabolism of some substrates is rate-limiting.
When the effect of cytochrome b5 deletion was investigated using a set of probe drugs, hepatic activities for chlorzoxazone (Cyp2e), midazolam (Cyp2c, Cyp3a), metoprolol (Cyp2d), and tolbutamide (Cyp2c) were markedly reduced (Fig. 4), indicating that cytochrome b5 is important for maximal turnover of these substrates. The metabolism of phenacetin, which is chiefly mediated by Cyp1a2, was either unaffected (kidney, lung) or increased (liver) in response to cytochrome b5 deletion again suggesting that cytochrome b5 is not important in Cyp1a2 function. It is noteworthy that cytochrome b5 deletion in the small intestine also caused a reduction in turnover of midazolam and metoprolol, indicating that perturbations in gut cytochrome b5 levels may have a bearing on the bioavailability of orally dosed drugs.
To investigate more fully the mechanism of cytochrome b5 interaction with cytochrome P450, the kinetic parameters of MR, which is metabolized by Cyp1a2, and is unaffected by cytochrome b5 (Table 1) and BFC, which is metabolized by multiple P450s (Cyp3a, Cyp2a, Cyp2b, Cyp2c) and is significantly affected by cytochrome b5,were determined. The Km and Vmax values for MR were not markedly different between BCN and wild type, whereas for BFC there was a 5-fold increase in Km and a 4.5-fold decrease in Vmax in the BCN samples (Table 2). The alteration in Km for BFC provides evidence to support theory that cytochrome b5 effects P450 activity through allosteric interactions with the metabolizing cytochrome P450s, causing a conformational shift in the active site, which affects substrate binding (Bridges et al., 1998; Hlavica and Lewis, 2001). Alternatively, the change in Km observed for BFC could be attributable to multiple P450s, differentially up-regulated by the deletion of cytochrome b5 contributing to the metabolism of this substrate. There is evidence to support the binding of cytochrome b5 to mammalian CYP1A2 (Shimada et al., 2005); therefore, the lack of effect on the Km for MR in the BCN samples suggests that cytochrome b5 binding does not significantly alter Cyp1a2 active site conformation. This is supported by evidence that indicates that unlike enzymes such as CYP3A4 and CYP2B4, CYP1A2 has an extremely stable and inflexible active site (Anzenbacher and Hudecek, 2001).
The generation of global and liver-specific models for cytochrome b5 deletion now allows the contribution of this protein in extrahepatic and hepatic P450 drug metabolism to be assessed in greater detail. We have shown previously that hepatic deletion of cytochrome b5 did not make a significant difference on the in vivo metabolism of metoprolol (Finn et al., 2008). In contrast, determination of the in vivo pharmacokinetic parameters of metoprolol in BCN mice provides evidence that extrahepatic tissues play a key role in its metabolism, because in this model, the area under the curve and Cmax are increased 3.5- and 4-fold, respectively, and clearance is decreased by 3.5-fold compared with wild-type mice (Fig. 5). In support of this, it has been shown recently that in mouse, intestinal P450s play important roles in drug pharmacokinetics (van Herwaarden et al., 2009). These data reinforce our previous conclusions that the level and/or the activity of cytochrome b5 can play a significant role in the metabolism, disposition, and therapeutic effectiveness of clinically used drugs.
A particularly well characterized role for cytochrome b5 is in steroid hormone biosynthesis, in which it enhances the lyase activity of Cyp17 through an allosteric mechanism but has little effect on the hydroxylase activity (Auchus et al., 1998; Akhtar et al., 2005; Naffin-Olivos and Auchus, 2006). Testicular Cyp17 activity was severely compromised in BCN samples, not only for the lyase reaction (1% of wild type) but also in progesterone hydroxylation (50% of the wild-type rate) (Fig. 6A). In addition, BCN mice had a significantly lower concentration of testicular testosterone (Fig. 6B), with no change in levels of Cyp17 protein observed (Fig. 6D). It seems, therefore, that although Cyp17 lyase activity is almost completely cytochrome b5-dependent, the presence of cytochrome b5 is not an obligatory requirement for successful testosterone biosynthesis.
In summary, we have generated a mouse model in which microsomal cytochrome b5 is completely deleted, and we characterized the effect on cytochrome P450 expression and metabolism both in vitro and in vivo. The data presented provide evidence that cytochrome b5 can have a marked effect on the rate of substrate turnover, particularly in extrahepatic tissues such as the small intestine and lung, in which it can be rate-limiting. Cytochrome b5 deletion profoundly alters the in vitro activity of Cyp17; however, this results in only a halving of testicular testosterone levels, which has no detrimental effect on reproductive ability. The BCN model provides an important tool that, in conjunction with the HBN mouse (Finn et al., 2008), will further our understanding of the many and varied physiological roles of this protein. In addition, it is known that levels of cytochrome b5 in humans are subject to significant variation (Forrester et al., 1992), which could contribute to individual differences in drug pharmacokinetics.
Acknowledgments
We thank Catherine Hughes, Susanne van Schelven, and Jennifer Kennedy for excellent technical assistance.
Footnotes
-
This work was supported by the Cancer Research UK Programme [Grant C4639/A5661].
-
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.110.064246.
-
ABBREVIATIONS:
- P450
- cytochrome P450
- BCN
- cytochrome b5 complete knockout mice, Cytb 5(−/−)
- BFC
- 7-benzyloxy-4-trifluoromethylcoumarin
- BR
- benzyloxyresorufin
- EFC
- 7-ethoxy-4-trifluoromethylcoumarin
- ER
- ethoxyresorufin
- HPLC
- high-performance liquid chromatography
- LC/MS-MS
- liquid chromatography/tandem mass spectrometry
- MFC
- 7-methoxy-4-trifluoromethylcoumarin
- MR
- methoxyresorufin
- POR
- cytochrome P450 oxidoreductase
- PR
- pentoxyresorufin
- WT
- wild type, Cytb5(+/+)
- PCR
- polymerase chain reaction.
- Received February 17, 2010.
- Accepted April 29, 2010.
- Copyright © 2010 The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}