


Abstract
Cytochrome P4502C19 (CYP2C19) is an important drug-metabolizing enzyme involved in the biotransformation of, for example, proton pump inhibitors and antidepressants. Several in vivo studies have shown that the CYP2C19 activity is inhibited by oral contraceptives, which can cause important drug interactions. The underlying molecular mechanism has been suggested to be competitive inhibition. However, the results presented here indicate that estradiol derivatives down-regulate CYP2C19 expression via estrogen receptor (ER) α, which interacts with the newly identified ER-binding half site [estrogen response element (ERE)] at the position −151/−147 in the CYP2C19 promoter. In gene reporter experiments in Huh-7 hepatoma cells, the activity of the luciferase construct carrying a 1.6-kb long CYP2C19 promoter fragment cotransfected with ERα was down-regulated upon treatment with 17β-estradiol (EE) or 17α-ethinylestradiol (ETE) at half-maximum concentrations of 10−7 and 10−8 M, respectively. Mutations introduced into the ERE half site −151/−147 significantly inhibited these ligand-dependent effects. Electrophoretic mobility shift assays and quantitative chromatin immunoprecipitation experiments revealed that estrogen receptor α binds to this element. A significant suppression of CYP2C19 transcription by female sex steroids was confirmed by reverse transcription polymerase chain reaction after hormonal treatment of human hepatocytes. Inhibition experiments using a stable human embryonic kidney 293 CYP2C19 cell line revealed competitive inhibition at much higher concentrations of EE and ETE compared with those required for transcriptional inhibition. These results indicate that both EE and ETE inhibit CYP2C19 expression via an ERα-dependent regulatory pathway, thus providing a new insight into the molecular mechanism behind the inhibitory effect of oral contraceptives on CYP2C19 activity.
Introduction
Cytochrome P4502C19 (CYP2C19) is the main enzyme for the metabolism of many important drugs, including protein pump inhibitors such as omeprazole, diazepam, the antidepressant escitalopram, the platelet aggregation inhibitor clopidogrel, and the antimalaria drug proguanil (Desta et al., 2002; Kirchheiner et al., 2004; Klotz, 2009). CYP2C19 is known to be polymorphically expressed, which strongly influences the effect of drug treatment efficacy (Sim et al., 2006). The enzyme is mainly present in the liver and intestine (Klose et al., 1999; Läpple et al., 2003; Wu et al., 2006). In addition, its activity is also affected by different drugs, which may potentially lead to unwanted drug-drug interactions associated with a higher risk of drug side effects or with a failure of therapeutic efficacy (Ingelman-Sundberg et al., 2007; Ingelman-Sundberg, 2008). Several reports have shown that oral contraceptive (OC) formulations substantially decrease CYP2C19 activity (Hägg et al., 2001; Palovaara et al., 2003). The mechanism behind this has been suggested to be drug-drug interactions at the enzyme active site (Laine et al., 2000). A modulatory effect of exogenous female sex steroids is not only confirmed for CYP2C19 but has also been shown for the enzymes CYP2A6 and CYP2C9 in vitro and in vivo (Sandberg et al., 2004; Benowitz et al., 2006; Higashi et al., 2007). Despite the fact that, each year, approximately 70 million women worldwide take such pills (Belle et al., 2002), the mechanism by which estrogen derivatives affect CYP2C19 activity has not been yet elucidated. Such regulation might be also important with respect to gender differences in drug metabolism as observed with other cytochrome P450 enzymes (Wolbold et al., 2003).
17α-Ethinylestradiol (ETE) and estradiol (EE) are the major estrogens used in oral contraceptives and hormone replacement therapy. In clinical use, these estrogens are commonly combined with progestins to obtain better hormonal cycle control (Laine et al., 2003). It has been shown that OCs double the area under the curve of the CYP2C19 substrate omeprazole and the metabolic ratio of mephenytoin (Tamminga et al., 1999; Hägg et al., 2001). Both substances are the most commonly used markers of CYP2C19 activity. The inhibiting effect on CYP2C19 activity has been demonstrated to be exerted by the OC compound 17α-ethinylestradiol and not by progestins, which are also included in OC formulations (Palovaara et al., 2003).
Estrogenic and antiestrogenic effects are mediated through two related estrogen receptors ERα and ERβ (van de Stolpe et al., 2004). The ERs exert their transcriptional control upon interaction of the active receptor dimer with specific DNA binding sites, such as classic estrogen responsive elements [EREs, consensus sequence 5′-AGGTCAnnnTGACCT-3′ (Couse et al., 1997; Driscoll et al., 1998) or estrogen receptor binding half-sites (ERE-half-sites (van de Stolpe et al., 2004)]. Antiestrogens can act as pure estrogen antagonists [e.g., ICI 182,780 (fulvestrant)] or as partial agonists (also known as agonists/antagonists or selective estrogen receptor modulators) such as tamoxifen, its potent metabolite 4-hydroxytamoxifen (4-OHT), or raloxifene (R), which exert tissue-selective estrogen-like effects. The mechanism by which these compounds induce tissue- and gene-dependent effects remains to be unraveled (Fournier et al., 2001). In addition to the ligand dependent activation of ERα, ligand-independent activation pathways of ERα (most likely through phosphorylation of the N terminus of the receptor) have been described previously (Ignar-Trowbridge et al., 1992, 1996) The estrogen interaction with CYP2C19 might also be important for breast cancer and breast cancer treatment because persons carrying the CYP2C19*17 allele, which is more highly expressed than the wild-type allele (Sim et al., 2006), are protected from breast cancer (Justenhoven et al., 2009), and CYP2C19*17-carrying breast cancer patients have better prognosis after treatment with tamoxifen (Schroth et al., 2007).
In the current study, we experimentally tested the hypothesis that the decline in CYP2C19 activity in women using steroid oral contraceptives is caused by a transcriptional down-regulation of CYP2C19 gene expression rather than direct steroid-CYP2C19 protein interaction. Indeed, results from different experimental approaches indicate that both EE and ETE are able to induce binding of ligand-activated ERα to the identified specific ERE consensus half-site within CYP2C19 promoter with the ensuing inhibition of CYP2C19 expression at physiological hormone concentrations, which are significantly lower than those required for competitive inhibition using stably CYP2C19 enzyme expressing cells.
Materials and Methods
Plasmid Constructs and Chemicals.
A 1.6-kb long fragment of CYP2C19 5′-flanking region was subcloned into the MluI/XhoI site of pGL3basic vector (Promega, Madison, WI) upstream of the luciferase gene (construct 2C19wt_−1.6kb; Table 1, Fig. 1b). A construct containing destructive mutations at the possible ERE half-site at position (−151/−147) was generated from the parental 2C19wt_−1.6kb construct using GeneTailor Mutagenesis kit (Invitrogen) (construct 2C19mut_−1.6kb Table 1, Fig. 1b).
Oligonucleotides used for cloning, EMSA, and ChIP experiments
Primers for PCR cloning were designed based on the CYP2C19 sequence (GenBank accession number gi 27498938:1708014-1877575). Fragments for the reporter constructs were PCR-amplified from genomic DNA. Nonsense mutations in ERE half site I were introduced using gene tailor mutagenesis kit (Invitrogen). Mutated nucleotides in ERE half-site I are underlined. ERE half sites and mutations at ERE half sites are underlined. The ChIP primer pair generates a 305-bp PCR fragment including ERE half-site I.
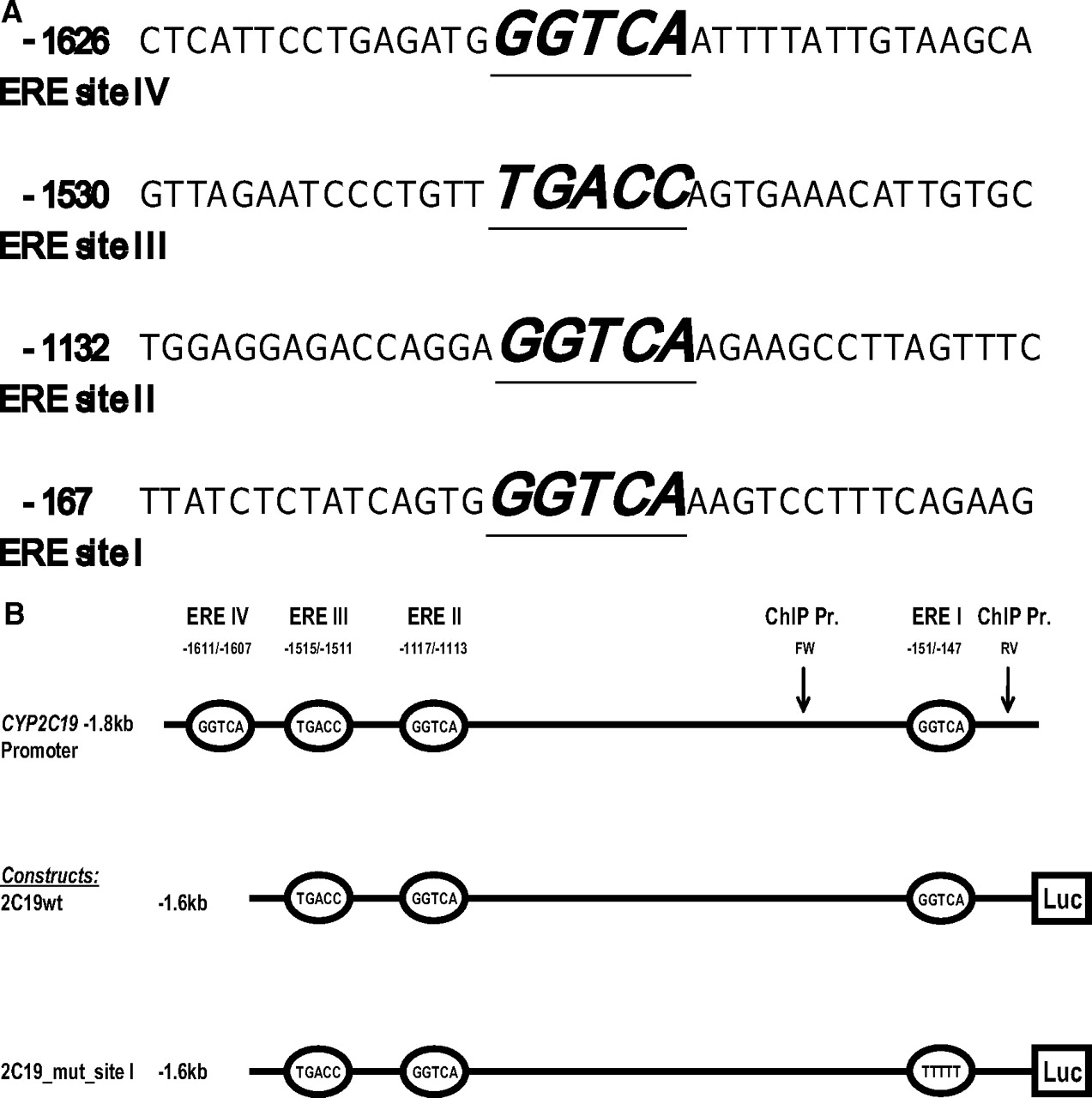
Schematic representation of ERE half-site motifs and luciferase reporter constructs. A, the DNA sequence surrounding the putative ERE half-sites of CYP2C19 gene promoter. The A of the first codon ATG is numbered as +1. The sequences of sites I to IV core regions are italic, underlined, and bold. B, schematic overview of CYP2C19 promoter fragments subcloned upstream to luciferase reporter gene. Numbers refer to the fragment length or location of putative ERE binding motifs. Mutated ERE half-site is indicated in italic. The binding sites for the forward (FW) and reverse (RV) primers used in ChIP analysis are indicated by arrows.
The expression vector encoding human estrogen receptor α (pSG5-hERα) was a kind gift of Dr. P. Chambon (Strasbourg, France). The ERα insert was recloned into the EcoRI site of pcDNA3.1 empty vector (Invitrogen). Chemicals (17α-ethinylestradiol, 17β-estradiol, 4-hydroxytamoifen, raloxifene) were purchased from Sigma (St. Louis, MO).
Electrophoretic Mobility Shift Assay.
Nuclear protein extracts were prepared from human-derived hepatoma cell lines Huh-7 and HepG2 as described previously with slight modifications (Dignam et al., 1983; Nakabayashi et al., 1991). ERα was in vitro-translated using pcDNA3.1/ERa vector in the TnT T7-Coupled Reticulocyte Lysate System (results shown in supplemental data) (Clontech, Mountain View, CA).
Three different double-stranded (ds) oligonucleotides comprising 30 bp of the CYP2C19 promoter in wild type or in mutated form at the putative ERE half-site were generated by annealing sense and antisense oligonucleotides (Table 1). Double-stranded oligonucleotides were 32P-labeled (GE Healthcare) using the T4 DNA polynucleotide kinase system (Invitrogen) in a final reaction volume of 25 μl.
The labeling reaction was carried out at 37°C for 15 min. The reaction was stopped with 5 μl of 0.2 M Na2EDTA. Binding reactions were carried out in 4% glycerol, 8 mM HEPES (Sigma Chemical Co, St. Louis, MO), pH 7.9, 0.6 mM MgCl2, 50 mM NaCl, 2 μg of poly[dI-dC]·poly[dI-DC], 12.8 fmol of 32P-labeled double-stranded probe (approximately 20,000 cpm), and 16 μg of HepG2 cell NE or Huh-7 cell NE in a total volume of 25 μl.
Mixtures with nuclear extract were preincubated at 37°C for 15 min followed by addition of the respective labeled double-stranded oligonucleotides. The complete mixture was incubated again at 37°C for 15 min. For competition experiments, 100- and 200-fold excess of the respective unlabeled double-stranded oligonucleotide was added to the probe before the addition of 32P-labeled double-stranded oligonucleotides. For supershift experiments, 2 μg of ERα or ERβ antibody (Santa Cruz Biotechnology, Santa Cruz, CA) was added to the binding reaction, and samples were incubated on ice for 45 min before the incubation with labeled oligonucleotides. Five microliters of loading buffer were added and protein-bound as well as unbound DNA was resolved on a 4% nondenaturing polyacrylamide gel. Dried gels were subjected to filmless autoradiographic analysis (BAS-1800; Fujifilm, Tokyo, Japan).
Transient Transfections.
Huh-7 cells were grown at 37°C in a humidified atmosphere with 5% CO2 in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin (Invitrogen, Carlsbad, CA). One day before transient transfection, 2 × 105 Huh-7 cells were plated into 12-well plates. Two micrograms of the pGL3basic expression vector CYP2C19–1.6kb (wild-type or mutated form) or pGl3basic were cotransfected with 0.5 μg of human pcDNA3.1-ERα or 0.5 μg of pcDNA3.1 empty vector (negative control). Transient transfections were carried out in plain medium using Lipofectamine 2000 (Invitrogen) as to manufacturer's recommendations. 24 h after transfection medium was replaced to complete medium and cells were treated (as indicated in each figure) with final concentrations of 10−10 to 10−6 M 4-OHT, R, EE, or ETE. Substrates were diluted in 95% ethanol (vehicle) and applied at a final concentration of 0.2% (v/v) vehicle per well. Cells were harvested and analyzed for luciferase activity 24 h after treatment. In each transfection mixture, 6 ng of pRL-TK vector (Promega) was included as an internal control for transfection efficiency. Luciferase activity is expressed, therefore, as a ratio of firefly luciferase activity (in arbitrary units) to the corresponding activity of the pRL-TK. All experiments were performed in triplicate and repeated three times.
ChIP Analysis.
The assay was carried out using chromatin immunoprecipitation (ChIP) assay kit (Active Motif Inc., Carlsbad, CA) according to the manufacturer's protocol. In summary, Huh7 cells were grown to 80% confluence in two 75-cm2 flasks for each treatment. Proteins were cross-linked in 1% formaldehyde, washed with phosphate-buffered saline, fixed in glycine solution, and scraped into 2 ml of phosphate-buffered saline supplemented with phenylmethylsulfonyl fluoride. Cells were lysed, homogenized, and enzymatically sheared with a subsequent immunoprecipitation using ERα antibodies (Santa Cruz Biotechnology) or control IgG (negative control). The resulting immunoprecipitates were subjected to PCR using a primer set (Table 1) that amplifies a 305-bp fragment of CYP2C19 promoter encompassing the putative ERE half-site at position −151/−147. The PCR was carried out for 35 cycles at an annealing temperature of 58°C. Quantitative evaluation of the PCR products was done by real-time PCR on the 7500 Fast Real-Time PCR system using a custom TaqMan Gene Expression assay (Applied Biosystems, Foster City, CA). The assay was constructed and validated by Applied Biosystems using the same proximal promoter fragment as a template.
Establishment of CYP2C19 Stable Cell Line.
The Flp-In system (Invitrogen) was used for the construction of the stable cell line expressing CYP2C19. Cells were maintained in high-glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin (Invitrogen) at 37°C in a humidified atmosphere with 5% CO2. Before transfections, Zeocin (Invitrogen) was added to the cell culture medium at a final concentration of 100 μg/ml. CYP2C19 cDNA was inserted into the pcDNA5/FRT plasmid and used in transfections of Flp-In- 293 cells. The CYP2C19-pcDNA5/FRT plasmid was cotransfected with the Flp recombinase expression plasmid pOG44 using Lipofectamine 2000 (Invitrogen) according to manufacturer's protocol. Single colonies resistant to hygromycin B (Invitrogen) were selected and subcultured. Selection of positive clones was performed by detection of CYP2C19 mRNA and enzyme activity as described below. Mock cells were transfected with pcDNA5/FRT and pOG44 plasmids and prepared as described above.
Hormone Treatment of Human Hepatocytes.
Human hepatocytes were obtained from CellzDirect/Invitrogen. Cells were seeded into 24-well plates at 105 cells/well using Williams' Medium E without Phenol Red supplemented with fetal bovine serum, plating cocktail solution, and dexamethasone. After 5 h, cells were treated with EE and ETE at concentrations of 10−10 to 10−6 M using ethanol as vehicle. Cells were maintained in estrogen treatment for 24 h until harvesting.
Detection of mRNA by RT-PCR.
Total RNA was isolated from cultured Flp-In- 293/CYP2C19 cells or from human hepatocytes with RNeasy Mini Kit (QIAGEN, Valencia, CA) according to the manufacturer's protocol. cDNA was prepared from the extracted RNA using reverse transcriptase reaction with SuperScript II RT (Invitrogen). mRNA levels of pcDNA5/FRT-CYP2C19 cells or human hepatocytes were determined by quantitative PCR on a 7500 fast real-time PCR system using SYBR Green (Applied Biosystems) approach with the following CYP2C19-specific primer set: forward, 5′-GAAAAATTGAATGAAAACATCAGGATTG; reverse, 5′-CGAGGGTTGTTGATGTCCATC. As internal control, TBP mRNA levels were codetermined using the primer pair 5′-GAA ACG CCG AAT ATA ATC CCA (forward) and 3′-GCT GGA AAA CCC AAC TTC TG (reverse).
Detection of Enzyme Activity by P450-Glo Assay.
Flp-In- 293/CYP2C19 cells were seeded in 96-well plates, and measurements were made at approximately 80% confluence. Cells were incubated with 20 μM Luciferin-H EGE, a CYP2C19 substrate, for 30 min at 37°C. Inhibitors were added to cells 5 min before substrate incubation. Luminescence produced was proportional to CYP2C19 activity and was measured in the cell medium according to manufacturer's protocol (P450 Glo Assay; Promega) using a TD-20/20 Luminometer (Turner Designs, Sunnyvale, CA).
CYP2C19 Enzyme Activity in Microsomes.
CYP2C19 enzyme activity and IC50 were measured in the microsomes from the insect cells overexpressing CYP2C19 using the CYP2C19/CEC High Throughput Inhibitor Screening Kit (BD Biosciences, San Jose, CA).
Statistical Analysis.
Statistical differences in reporter gene activity among CYP2C19 promoter constructs and in mRNA levels in human hepatocytes were determined by one sample t test using Prism (ver. 5.00 for Windows; GraphPad Software, San Diego, CA). A P value threshold of 0.05 was considered statistically significant in all analyses.
Results
ERα Binds Specifically to an ERE Half-Site Located at −151/−147 in the CYP2C19 promoter.
Analysis of the first 1.8 kb of CYP2C19 promoter revealed four different putative ERE half-sites at positions (−151/−147), (−1117/−1113), (−1515/1511), and (−1611/−1607) (Fig. 1A). We tested by way of EMSA whether ERα or ERβ could interact with any of these sites. Four different 32P-labeled double stranded oligonucleotides (Table 1) carrying putative ERE half-sites were incubated with nuclear extracts from Huh-7 cells and HepG2 cells. A specific protein-DNA complex was formed only with oligonucleotide 2C19_wt_site I using both HepG2 (Fig. 2A, lane 2) and Huh-7 cell nuclear extracts (Fig. 2B, lane 1). No specific complexes were observed with oligonucleotides 2C19_wt_site II to IV (data not shown). The migration of site I-specific complex was shifted using an antibody against ERα (Fig. 2, A, lane 8, and B, lane 7). An antibody against ERβ, however, failed to produce any band shifts, confirming that ERβ, which recognizes the same consensus sequence as ERα, does not bind to ERE half-site I (Fig. 2, A, lane 9, and B, lane 8). The specificity of the protein/DNA complex at locus −151/−147 (site I) was confirmed in competition experiments using nonlabeled oligonucleotides and also oligonucleotide comprising an ERE consensus cis-element (Fig. 2, A and B). Moreover, no or reduced specific binding activity could be observed after incubation of nuclear extracts from Huh-7 cells or HepG2 cells with oligonucleotides mutated at the hypothetical ERα binding half-site I (Fig. 2, A and B). ERα interaction with the CYP2C19 promoter was also confirmed in EMSA experiments in which the labeled oligonucleotide 2C19_wt_site I was incubated with the in vitro-translated receptor (results shown in the supplement). Overall, these results suggest that ERα but not ERβ is capable of binding to the potential ERE site at the position −151/−147 of CYP2C19 promoter.

ERα interacts with oligonucleotides containing ERE half-site motifs. EMSA was performed with a double-stranded oligonucleotide (Table 1) comprising the putative ERE half-site I (−151/−147). This oligonucleotide formed a positive DNA-protein-complex with nuclear extract from HepG2 (A, lane 2, white arrow) or Huh7 cells (B, lane 1, white arrow). Competition experiments were performed with the respective nonlabeled wild-type oligonucleotide (A, lanes 3 and 4; B, lanes 2 and 3) and an oligonucleotide comprising a representative ERE consensus sequence (Higashi et al., 2007), (A, lanes 5 and 6; B, lanes 4 and 5). Supershifts were observed with both HepG2 and Huh-7 NE at ERE half-site I using ERα antibody (indicated by black arrows; A, lane 8; B, lane 7). No band shifts were generated with ERβ antibody (A, lane 9; B, lane 8). −, control incubation without labeled oligonucleotide; C, DNA-protein complex; Comp, competition reactions with wild-type oligonucleotide; ERE cons, competition reactions using an oligonucleotide carrying a representative ERE consensus sequence; 2C19 mut oligo, oligonucleotide with destructive mutations in ERE half-site I; anti-ERα and anti-ERβ, antibodies against the respective receptors.
Estradiol and 17α-Ethinylestradiol Inhibit CYP2C19 Promoter Activity via the ERα Dependent Transactivation.
The functional interaction of ERα with the CYP2C19 promoter was evaluated using reporter plasmids transfected into human hepatoma Huh-7 cells. A plasmid construct carrying 1.6 kb of the CYP2C19 promoter (Fig. 1B), upstream of the reporter luciferase gene, was transiently transfected together with ERα expression vector and the cells were treated with increasing amounts of the ERα ligands ETE, EE, 4-OHT, or R. The results shown in Fig. 3 demonstrate a strong and dose-dependent down-regulation of reporter luciferase activity upon treatment with EE or ETE. The half-maximal inhibitory effect of EE and ETE was observed at 10−7 and 10−8 M, respectively, and the extent of inhibition at 10−6 M was approximately 70% using both steroids. Contrary to these results, 4-OHT and R were unable to affect the CYP2C19 promoter activity in this system (data not shown).

ERα ligands ETE and EE down-regulate the luciferase activity of CYP2C19 promoter constructs in Huh-7 cells. Relative luciferase activities of −1.6-kb CYP2C19 promoter fragment subcloned into pGL3basic vector. Data are shown as fold changes of luciferase activity relative to the activity of wild-type construct treated with vehicle after cotransfection with pcDNA3.1-ERα or mock (pcDNA3.1) and stimulation with ETE (A) or EE (B). ***, p ≤ 0.001 for 2C19_wt treated with vehicle compared with 2C19_wt treated with ETE (10−6 M, A) and EE (10−7 M, B), respectively. **, p ≤ 0.01 for 2C19_wt treated with vehicle compared with 2C19_wt treated with ETE (10−7 or 10−6 M, A) and EE (10−6 M, B), respectively. *, p ≤ 0.05 for 2C19_wt treated with vehicle compared with 2C19_wt treated with ETE (10−8 M, A) and EE (10−8 M, B), respectively. Data are presented as mean values ± S.D. of three independent experiments. Each experiment was performed in triplicate.
To investigate whether the ERE half-site I is indeed responsible for the observed estrogen-dependent down-regulation of CYP2C19 promoter activity, as suggested by EMSA experiments, a luciferase construct carrying a mutant form of ERE half-site I (Fig. 1B) was tested in the gene reporter experimental setup using ERα ligands at the highest effective concentration (10−6 M). As shown in Fig. 4, the inhibitory effect of EE and ETE was partly abolished upon transfection of the mutated construct. These results support the hypothesis of a ligand-activated ERα effect in the suppression of CYP2C19 promoter.

Mutation in the ERE half-site I partly abolishes the inhibitory effects of ETE and EE on the luciferase activity of CYP2C19 promoter construct in Huh-7 cells. Relative luciferase activities of −1.6-kb CYP2C19 promoter fragment subcloned into pGL3basic vector. Data are shown as fold changes of luciferase activity of the construct mutated at ERE half-site I relative to the activity of the wild-type construct after cotransfection with pcDNA3.1-ERα or mock (pcDNA3.1) and treatment with ETE 10−6 M (A) or EE 10−6 M (B) or vehicle. A and B: ***, p ≤ 0.001 for 2C19_wt treated with vehicle compared with 2C19_wt treated with ETE (10−6 M, A) or EE (10−6 M, B). **, p ≤ 0.01 and *, p ≤ 0.05 for CYP2C19_M1 treated with vehicle against 2C19_M1 treated with ETE (10−6 M, A) or EE (10−6 M, B). **, p ≤ 0.01 for 2C19_wt treated with ETE (10−6 M, A) or EE (10−6 M, B) compared with 2C19_M1 treated with ETE (10−6 M, A) or EE (10−6 M, B). Data are presented as mean values ± S.D. of three independent experiments. Each experiment was performed in triplicate.
Estradiol and 17α-Ethinylestradiol Suppress CYP2C19 mRNA Expression in Hepatocytes.
To further examine the effects of estrogen derivatives on hepatic CYP2C19 expression, we determined the CYP2C19 mRNA levels in dependence of estradiol or 17α-ethinylestradiol treatments using three different batches of human hepatocytes. Cells were either treated with vehicle (ethanol), EE, or ETE. As shown in Fig. 5, real-time RT-PCR revealed that CYP2C19 mRNA levels were significantly suppressed by treatments with EE at concentrations of 10−7 M. The effect of EE was found to be stronger compared with the effect seen with ETE. These results show that estrogen derivatives are indeed able to significantly inhibit CYP2C19 expression in human hepatocytes, most likely via the proposed ERα-dependent promoter-regulatory pathway, although one has to bear in mind that during cultivation, the hepatocytes lose their ability to express P450s and the system might not be optimal for this reason.

ERα ligands ETE and EE down-regulate mRNA expression of CYP2C19 in human hepatocytes. Human hepatocytes were treated with EE or ETE or with vehicle for 24 h. The expression levels of CYP2C19 mRNA were determined by real-time RT-PCR. Values of CYP2C19 mRNA levels were normalized with TBP mRNA levels and compared with the ratios obtained in vehicle-treated cells. Each column represents the mean ± S.D. of three independent mRNA measurements. **, p ≤ 0.01.
ERα Is Associated with CYP2C19 Promoter in Huh-7 Cells.
EMSA analysis and gene reporter data suggest that CYP2C19 expression might be regulated by ER-dependent transcriptional mechanisms. To investigate whether ERα is indeed associated with the putative cis-element in CYP2C19 promoter and whether ER ligands could enhance such binding as it occurs in the classic scenario of the nuclear receptor mediated modulation of transcriptional activity, we used the advantage of the ChIP technology. Genomic DNA from intact Huh-7 cells, which were either not treated or treated with ETE or 4-OHT, was isolated, cross-linked, sonicated, and immunoprecipitated with ERα antibody. The resulting DNA fragments were amplified by PCR using a primer set encompassing ERE half-site I at position (−151/−147) (Table 1, Fig. 1A). As shown in Fig. 6, top, the primer set generates a PCR product of the predicted size with DNA from vehicle and ETE-treated cells, whereas the 4-OHT treatment resulted in the loss of the band. These results were quantitatively supported by real-time PCR using a custom-made TaqMan assay that was constructed on the basis of the same promoter fragment (Fig. 6, bottom). The amount of PCR product from the ETE-treated cells was 2.3-fold higher compared with the control (vehicle) with negligible amounts detected in the 4-OHT-treated cells. This is consistent with the EMSA results that demonstrated interaction of the ERE oligonucleotides with the nuclear ERα from the intact cells. Stimulation of Huh-7 cells with ETE resulted in the 2.3-fold increase of the PCR product (Fig. 6). It is noteworthy that the 4-OHT treatment apparently abolishes the ERα binding as judged by the sharp drop in the amount of the expected PCR product. These results are in good agreement with the gene reporter and EMSA data indicating that ERα may indeed interact with CYP2C19 promoter in vivo and that this binding is regulated by ER ligands.

ERα is associated with CYP2C19 promoter at ERE half-site I in Huh7 cells. ERα binding to CYP2C19 promoter was analyzed using ChIP assay. Top, immune complexes generated by anti-ERα and control IgG were PCR-amplified (35 cycles) using the primer set for ChIP (indicated in Table 1 and Fig. 1B) targeting ERE half-site I in CYP2C19 promoter. Agarose gel electrophoresis of PCR products demonstrates a predicted amplicon size of approximately 300 bp. Bottom, real-time PCR using the same template as above and a standard cycling program suggested by Applied Biosystems for the amplifications with TaqMan probes (Materials and Methods). Results are expressed as a difference between the Ct (cycle threshold) for the PCR product from the control IgG immunoprecipitate and anti-ERα immunoprecipitate (target). Input is a positive PCR control (same fragments amplified from the starting material before the immunoprecipitation). V, vehicle; IP, immunoprecipitation.
High Concentrations of ETE and EE Directly Inhibit CYP2C19 Enzyme Activity.
Clinical data on the lower activity of CYP2C19 in women using OCs were previously interpreted as a direct inhibitory action of steroids on the CYP2C19 enzyme (Laine et al., 2000). To investigate such a scenario, we tested the effects of ETE and EE on CYP2C19 activity in human embryonic kidney 293 cells stably expressing CYP2C19. CYP2C19 expression was confirmed by both the mRNA (results not shown) and enzyme activity levels (Fig. 7, A and B). EE and ETE were found to inhibit CYP2C19 activity with IC50 values of 16 and 6.0 μM, respectively, as shown on Fig. 7C. This effect was confirmed by using CYP2C19 inhibitor screening kit from BD Biosciences. IC50 measurements using microsomes from insect cells overexpressing CYP2C19 showed lower levels (0.6 and 0.4 μM for ETE and EE, respectively; Fig. 7D), which might be connected with the better availability of inhibitors. However, both in cells and in the recombinant system, these values are 1 to 2 orders of magnitude higher compared with the above-mentioned data on the transcriptional effects of steroids.

High concentrations of ETE and EE directly inhibit CYP2C19 enzyme activity. Flp-In- 293/CYP2C19 cells were incubated with increasing concentrations of Luciferin-HEGE (CYP2C19 substrate) (A) or with 20 μM Luciferin-HEGE (measurements at various time points) (B). CYP2C19 activity in the cell medium was measured by the P450-Glo assay and expressed as mean arbitrary units of luminescence ± S.E.M. C, inhibition of CYP2C19 activity in Flp-In- 293/CYP2C19 cells by ETE or EE. As optimized in the experiments shown in A and B, the substrate concentration was set to 20 μM and incubation time to 30 min. ETE and EE were added 5 min before the enzyme assay. Enzyme activity is expressed relative to the activity of samples lacking ETE or EE. Data are presented as mean values ± S.E.M. All experiments were performed in triplicate. D, inhibition curve of CYP2C19 activity as measured by CYP2C19 High Throughput Inhibitor Screening Kit. Data are presented as in C.
Discussion
In the present study, we present evidence for that estrogen receptor α, activated by its ligands EE and ETE, is able to inhibit CYP2C19 expression via a newly identified ERE binding half-site at position (−151/−147) in the CYP2C19 promoter. This is the first study showing that estradiol and its derivatives are able to modulate CYP2C19 promoter activity, and this occurs via the classic ERE-dependent pathway. Hence, our data suggest a novel molecular mechanism for the inhibitory effect of ETE on CYP2C19 activity that has been observed in several clinical studies (Palovaara et al., 2003; Shelepova et al., 2005). This effect has been previously explained by a competitive inhibition at the active site of the enzyme (Laine et al., 2000).
In silico analysis of the first 1.8 kb of the 5′-upstream region of the CYP2C19 gene revealed four putative ERE half-sites. Although ERβ did not bind to any of the hypothetical ERE consensus sequences, which showed protein binding in EMSA analysis, a specific binding of ERα to CYP2C19 promoter at position (−151/−147) was confirmed by both EMSA and ChIP analysis using Huh-7 cells.
It is known that estrogen receptor α and β mediate their effects through at least four different pathways. In the so-called classic estrogen-dependent pathway, the hormone binds to the ER leading to conformational changes and dimerization of the receptor, which allows an interaction with comodulators and an eventual binding of ligand-receptor complex to the specific EREs in the regulatory regions of target genes (McDevitt et al., 2008). In addition, there are three other known mechanisms of ER activation, including second-messenger-dependent regulation of a membrane-associated ER and interaction with other transcription factors (McDevitt et al., 2008; Safe and Kim, 2008). Our results, especially those obtained in EMSA and ChIP, strongly support the important role of the classic regulatory pathway of estrogen-dependent CYP2C19 gene regulation. However, the destructive mutations in ERE half-site I were only partly able to abolish the inhibitory effects of ETE and EE using the reporter constructs in Huh-7 cells (see Figure 4). Therefore, we cannot completely exclude the possibility that other nonclassic ER-dependent pathways may also play a role in CYP2C19 regulation or that other, as-yet-detected ERE sites are also involved in the estrogen dependent regulation of CYP2C19.
CYP2C19 is not the only enzyme of the cytochrome P450 family that is known to be influenced by female sex steroids. For example, CYP2C9, which is very important for metabolism and dosing of, for example, warfarin and antiepileptics, is strongly inhibited by estrogen derivates (Laine et al., 2003; Sandberg et al., 2004). Indeed comparative sequencing analysis of the CYP2C9 promoter revealed that this enzyme carries an ERE similar to that identified here in the CYP2C19 gene, and it might be that the same mechanism is also relevant for estrogen-dependent CYP2C9 inhibition as here described for CYP2C19. In contrast, Benowitz et al. (2006) reported a significantly higher metabolism rate for the CYP2A6 substrates nicotine and cotinine in women compared with men and especially in women taking OCs. Similar results were obtained by Higashi et al. (2007), who proved experimentally that EE is able to induce CYP2A6 via an ERE element in CYP2A6 promoter. The modulatory factors determining enhanced or decreased promoter activity of the P450 genes in response to ER have to be identified. Our findings of an inhibitory effect of estrogens on CYP2C19 are well in line with earlier in vivo reports showing clinically relevant decreased metabolism of CYP2C19-specific substrates in women who regularly take pills that contain estradiol derivatives. This thus leads to important pharmacokinetic drug-drug interactions with other CYP2C19 substrates (Zhang et al., 2007).
As mentioned, it has been proposed that estrogen-dependent CYP2C19 inhibition may occur as a result of the competitive action of ETE on CYP2C19 enzyme (Laine et al., 2000, 2003). Here we have shown that ETE and EE indeed have the potential to directly inhibit CYP2C19 enzyme activity in intact cells overexpressing CYP2C19 or in microsomes but at much higher concentrations than required for transcriptional inhibition. These results are well in line with earlier observations (Laine et al., 2000, 2003) showing an effective in vitro inhibition of omeprazole 5-hydroxylation only at very high EE concentrations (>10−6 M). In contrast, transcriptional inhibition of CYP2C19 expression, as shown in this study, was exerted at much lower concentrations of EE and ETE, with IC50 at 10−7 to 10−8 M in luciferase gene reporter assay and maximum inhibitory effects on CYP2C19 mRNA levels at 10−7 M in human hepatocytes. This indicates that the mechanisms of OC-dependent inhibition of CYP2C19 activity in vivo would mainly be exerted by inhibition of CYP2C19 gene transcription rather than by competitive inhibition, and the results indicate for the first time an estrogen-dependent transcriptional regulation of this gene.
In conclusion, our results suggest a new regulatory mechanism by which exogenous estrogen derivatives unfold their inhibitory potential on CYP2C19. It could be also suggested that similar molecular mechanisms underlie the known inhibitory effects of female sex steroids on the activity of other cytochrome P450s (e.g., CYP2C9) (Laine et al., 2003; Sandberg et al., 2004).
Footnotes
This work was supported by Hjärnfonden, Torsten och Ragnar Söderbergs Stiftelser [Grant MT22/08]; the Swedish Research Council [Grant K2008-66X-05949-28-3]; the Danish Agency of Science, Technology and Innovation; the Lundbeck Foundation; and the Portuguese Foundation for Science and Technology [Grant SFRH/BPD/34152/2006; IBB-Institute for Biotechnology and Bioengineering/CBME-Centro di Biomedicina Molecular E Estrutural, LA, Fondo Europeo de Desarrollo Regional/POCI 2010].
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.110.065540.
-
ABBREVIATIONS:
- OC
- oral contraceptive
- ETE
- 17α-ethinylestradiol
- EE
- estradiol
- ER
- estrogen receptor
- ERE
- estrogen responsive element
- 4-OHT
- 4-hydroxytamoxifen
- R
- raloxifene
- EMSA
- electrophoretic mobility shift assay
- kb
- kilobase(s)
- NE
- nuclear extract
- ChIP
- chromatin immunoprecipitation
- PCR
- polymerase chain reaction
- bp
- base pair(s)
- RT
- reverse transcription.
- Received April 10, 2010.
- Accepted July 30, 2010.
- Copyright © 2010 The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}