


Abstract
Discovery of the multidrug resistance protein 1 (MDR1), an ATP-binding cassette (ABC) transporter able to transport many anticancer drugs, was a clinically relevant breakthrough in multidrug resistance research. Although the overexpression of ABC transporters such as P-glycoprotein/ABCB1, MRP1/ABCC1, and MXR/ABCG2 seems to be a major cause of failure in the treatment of cancer, acquired resistance to multiple anticancer drugs may also be multifactorial, involving alteration of detoxification processes, apoptosis, DNA repair, drug uptake, and overexpression of other ABC transporters. As a tool for the study of such phenomena, we designed and created a microarray platform, the ABC-ToxChip, to evaluate relative levels of transcriptional activation among genes involved in the various mechanisms of resistance. In the ABC-ToxChip, a comprehensive set of genes important in toxicological responses (represented by 2200 cDNA probes) is complemented with probes specifically matching ABC transporters as well as oligonucleotides representing 18,000 unique human genes. By comparing the transcriptional profiles of KB-3-1 and DU-145 parental cells with resistant derivatives selected in colchicine (KB-8-5), and 9-nitro-camptothecin (RCO.1), respectively, we demonstrate that ABC transporters (ABCB1/MDR1 and ABCC2/MRP2, respectively) show dramatic overexpression, whereas the glutathione S-transferase gene GST-Pi shows the strongest decrease in expression among the 20,000 genes studied. The results were confirmed by quantitative reverse transcription-polymerase chain reaction and immunohistochemistry. The custom-designed ABC-Tox microarray presented here will be helpful to elucidate mechanisms leading to anticancer drug resistance.
Mechanisms leading to multidrug resistance (MDR) include increased drug efflux (Shen et al., 1986b; Gottesman et al., 2002), decreased drug uptake (Shen et al., 2000), recruitment of drug-processing and metabolizing enzymes (Schuetz et al., 1996), the activity of conjugating enzymes (Goto et al., 2002), alteration of the DNA repair activity, reduction of cell susceptibility to apoptosis (Reinhold et al., 2003), and mutation of specific drug targets (Urasaki et al., 2001). In vitro-selected cell lines most likely use more than just one of the pathways cited above (Gottesman et al., 2002).
Tools, such as the oligonucleotide-based microarrays, cDNA-based microarrays, or serial analysis of gene expression (Velculescu et al., 1995) are relevant methods to screen for multifactorial mechanisms of drug resistance. However, microarray technology has several inherent problems that hamper definitive interpretation. It is generally necessary to validate results using a complementary technology, such as a different microarray platform or RT-PCR (Lee et al., 2003). Furthermore, the lack of sensitivity can prevent detection of infrequent transcripts (Evans et al., 2002) and lead to biased conclusions. In research aimed at the elucidation of mechanisms underlying acquired drug resistance, the principal shortcomings of arrays available at the time of our study were the low representation of genes of interest and the general lack of specificity of the probes. That was especially true for the ATP-binding cassette (ABC) superfamily, which has 48 members sharing high sequence similarity with highly divergent functions and/or specificities.
Like endogenous and exogenous toxic compounds, anticancer drugs diffuse through the cell membrane. One of the most interesting features of ABC transporters is that these proteins can transport many substrates across the membrane, including anticancer drugs, ions, and peptides. MDR1, or P-glycoprotein, was the first ABC transporter found to confer MDR in resistant tumor samples and in in vitro-selected cell lines (Shen et al., 1986a). With cell lines selected in various anticancer drugs, overexpression of further ABC transporters, such as ABCC1-MRP1 (Cole et al., 1992) and ABCG2-MXR-BCRP (Doyle et al., 1998; Miyake et al., 1999), was clearly shown to be associated with drug export and resistance. In experiments performed in vitro, other members, such as ABCA2 and ABCB4 (MDR3), were shown to actively transport cytotoxic drugs (Laing et al., 1998; Smith et al., 2000). Among the members of the MRP (ABCC) subfamily, MRP2 (ABCC2, canalicular multispecific organic anion transporter) transports GSH-S-conjugates (e.g., leukotriene C4 and 2,4-dinitrophenyl-S-GSH), oxidized GSH, glucuronide conjugates (e.g., glucuronidated bilirubin and bile salts), and sulfate conjugates of certain bile acids (e.g., 3-sulfato-lithocholytaurine) (Muller et al., 1994). By their ability to transport nucleoside analogs, ABCC3-MRP3 and ABCC5-MRP5 are thought to cause certain forms of drug resistance (Reid et al., 2003). ABCC11-MRP8 has also been associated with anticancer drug export (Guo et al., 2003; Szakács et al., 2004).
Although microarrays make possible the screening of thousands of genes at a time, attempts aiming to address anticancer drug resistance have been biased in their interpretation by use of microarrays bearing low numbers of ABC transporter superfamily probes (Lamendola et al., 2003). Herein, we report the design and the application of a dedicated ABC-ToxChip in which we combine probes for a comprehensive set of detoxifying genes with probes specifically matching ABC transporters. Instead of using the short 25-mer probe technology (Affymetrix, Santa Clara, CA), not optimized for detecting infrequent transcripts, we printed a combination of longer 70-mer oligonucleotide probes and 200- to 500-base pair fragments, thereby obtaining higher sensitivity because of the longer complementary sequences.
Camptothecin (CPT) derivatives such as CPT-11 (Irinotecan) and Hycamtin (Topotecan) are increasingly used in antitumor therapy against colon and lung cancers (Kudoh et al., 1998). 9-Nitro-camptothecin (9NC) has recently been studied in phase II trials for pancreatic cancer and is now in phase III trials (Pantazis et al., 2003). Many factors, such as specific mutations in topoisomerase I, complemented by a general alteration of apoptotic regulation, have been proposed to explain the phenotype of camptothecin resistance in a prostate cancer cell line (RCO.1) selected for resistance to 9NC (Chatterjee et al., 2001; Urasaki et al., 2001; Reinhold et al., 2003). To address specifically the roles of detoxifying enzymes and ABC transporters in camptothecin resistance, we compared the transcriptional profiles of DU145 prostate cancer cells and a 9NC-selected derivative cell line, RCO.1. Herein, we show that in RCO.1 cells, 9NC resistance is accompanied by the differential expression of ABCC2-MRP2 and enzymes regulating glutathione metabolism.
Materials and Methods
Taq polymerase and RT-PCR reagents were from Invitrogen (Carlsbad, CA). Reagents for quantitative RT-PCR were from Roche Diagnostics (Indianapolis, IN). High-density microarrays were printed in the National Institute of Environmental Health Sciences facility. 9NC was purchased from LKT Laboratories (St. Paul, MN).
Cell Lines and Cell Culture. DU-145 and a 9NC-selected derivative, RCO.1, were a generous gift from Dr. P. Pantazis (University of Miami, Coral Gables, FL). Cells were cultured in RPMI 1640 medium (Invitrogen) supplemented with 10% tetracycline-approved fetal bovine serum (Hyclone Laboratories, Logan, UT) and 2 mM l-glutamine (Quality Biological, Inc., Gaithersburg, MD) at 37°C in 5% CO2. Resistance to camptothecin analogs of RCO.1 was maintained with passage in camptothecin-containing medium (0.1 μM) every 3 months.
RNA Preparation. Total RNA was purified as described by Reinhold et al. (2003). For preparation of the RNA used in the microarray analysis or RT-PCR, cells were cultured in nonselective medium.
Microarray Design. A 20,000 human oligonucleotide/cDNA hybrid chip printed at the National Institute of Environmental Health Sciences (Microarray Group, National Institute of Environmental Health Sciences) was used for gene expression profiling experiments. The chip contained three categories of probes.
-
Ninety-six ABC-specific probes (matching 36 ABC transporters), consisting of either cDNA probes developed in-house or oligonucleotide probes obtained from QIAGEN Operon (Alameda, CA). To create the cDNA-probes, we performed PCR amplifications using specific primers on full-length cDNA sequences (ABCB1-MDR1, ABCC1-MRP1, ABCC2-MRP2, ABCG2-MXR, ABCC7-CFTR, and ABCB11) or on cDNA prepared from cells rich in a given transporter (J.P. Annereau, G. Szakacs, T. Fojo, M. M. Gottesman, unpublished data). We used a BLAST-based algorithm to increase specificity. First, we aligned the targeted transporter with members of its own subfamily to locate a candidate region representing minimal overlap. Specificity was then verified by blasting the candidate sequence against the whole nonredundant human database. The probes synthesized by RT-PCR were TA cloned (Invitrogen), as detailed in Table 1. ABC transporters are named using the HUGO Gene Nomenclature Committee nomenclature (http://nutrigene.4t.com/humanabc.htm). When more than one probe was used for an ABC transporter, the probe name was chosen according to the respective Ref-Seq entries of the National Center for Biotechnology Information database (i.e., ABCC3 (NM_003786, AJ294547.1, AJ294559.1, and AJ294558.1). Exons matching the probe sequences are indicated in Table 1.
-
Probes (2200) matching genes important in toxicological responses (cDNA clones from the National Institute of Environmental Health Sciences ToxChip version 3.0) (Nuwaysir et al., 1999). The National Institute of Environmental Health Sciences set contains approximately 2200 known human genes involved in pathways such as response to estrogens, polycyclic aromatic hydrocarbons, peroxisome proliferators, DNA damage, and oxidant stress, as well as genes involved in apoptosis, cell cycle, tumor suppression, signal transduction, and transcription.
-
Eighteen thousand genome-wide 70-mer probes obtained from QIAGEN Operon. A complete listing of the two sets (human oligonucleotide gene-set and the human TOX V1.0-set) printed on the ABC-ToxChip is available at http://dir.niehs.nih.gov/microarray/annereau/home.htm.
ABC probes printed on the ABC-ToxChip
Oligo indicates that the probe is a single-stranded, 70-mer oligonucleotide; cDNA indicates the probe is a fragment of double-stranded DNA amplified by PCR, ABC transporters are named using the HGNC nomenclature (http://nutrigene0.4t.com/humanabc.htm). The probes are named after matching RefSeq entries [National Center for Biotechnology Information, e.g., C3 (NM-003786, AJ294547.1, AJ294559.1, AJ294558.1)]. The exon most closely matching the sequence is indicated.
Gene Representation on the Microarray. The gene ontology classification (http://www.geneontology.org/#ontologies) annotates each gene in three categories: molecular function, biological process, and cellular component. To search the gene ontology (GO) reference for each gene represented on our microarray, we used the human genome annotation (HUGO) reference converted from the gene accession number by the source database (http://genome-www5.stanford.edu/cgi-bin/SMD/source/sourceSearch) and the Match-Miner database (http://discover.nci.nih.gov/matchminer/html/index.jsp). GO annotation keyword entries were performed using the QuickGO server at the European Bioinformatics Institute (http://www.ebi.ac.uk/ego/).
Microarray Spotting. The oligonucleotides were resuspended in ArrayIt spotting solution plus buffer (Telechem, San Jose, CA) and spotted at a concentration of 40 μM onto poly-l-lysine-coated glass slides using a modified, robotic DNA arrayer (Beecher Instruments, Bethesda, MD). The spotting was performed in an environmentally controlled chamber at 25°C and 40% relative humidity. After printing, the arrays were cross-linked in a Stratalinker at a power of 300 mJ and blocked with succinic anhydride/1-methyl-2-pyrrolidinone (protocol available at http://dir.niehs.nih.gov/microarray/methods.htm).
Microarray Hybridizations. Each total RNA sample (25 μg) was labeled with cyanine (Cy)3 or Cy5-conjugated dUTP (Amersham Biosciences, Inc., Piscataway, NJ) by a reverse transcription reaction using the reverse transcriptase SuperScript II (Invitrogen) and Oligo dT primer (Amersham Biosciences, Inc.). Each RNA pair was hybridized to a total of four arrays using a fluor reversal accomplished by labeling the control sample with Cy3 in two hybridizations and with Cy5 in the other two hybridizations. After hybridization, arrays were washed with Telechem wash buffers A, B, and C (Telechem International Inc., Sunnyvale, CA) for 2, 5, and 5 min, respectively. The hybrid chips were scanned with a scanner (Agilent Technologies, Wilmington, DE) using independent laser excitation of the 2 fluors at 532 nm and 635 nm wavelengths for the Cy3 and Cy5 labels, respectively. Results are available at http://dir.niehs.nih.gov/microarray/annereau/home.htm.
Outlier Filtering. Raw pixel intensity images were analyzed using ArraySuite version 2.0 extensions of the IPLab's image processing software package (Scanalytics, Fairfax, VA). The program uses methods that were developed and described previously by Chen et al. (2002) to locate targets on the array, measure local background for each target and subtract it from the target intensity value, and to identify differentially expressed genes using a probability-based method. The data were filtered with a cut-off at an intensity level just above the buffer blank measurement values to remove genes having one or more intensity values in the background range. After pixel intensity determination and background subtraction, the ratio of the intensities of the treated cells and control was calculated. The ratio intensity data from all probes on the human oligonucleotide chip was used to fit a probability distribution to the ratio intensity values. The resulting probability distribution was used to calculate a 95% confidence interval for the ratio intensity values. Genes having normalized ratio intensity values outside of that interval were considered to show significant differential expression.
Statistical Analysis and Filtering of Outliers. For each of the four replicate arrays for each sample, lists of differentially expressed genes at 95% confidence levels were created and deposited into the National Institute of Environmental Health Sciences MAPS database (Bushel et al., 2001). Genes that showed fluorescence bias or high variation were not considered for further analysis. Assuming that the replicate hybridizations are independent, a calculation using the binomial probability distribution indicated that the probability of a single gene occurring on this list when there was no real differential expression was <0.00048. The entire data set is available at http://dir.niehs.nih.gov/microarray/annereau/home.htm.
Identification of Relevant Biological Processes. The GoMiner software (Zeeberg et al., 2003), which can be downloaded at the web site of the Genomics and Bioinformatics Group, National Cancer Institute (http://discover.nci.nih.gov) is a computational resource for identifying biological processes that are altered in a disease phenotype. The algorithm maps genes with altered expression to the gene ontology. The gene ontology database lists three structured, controlled vocabularies (ontologies) that describe gene products in terms of their associated biological processes, cellular components, and molecular functions in a species-independent manner. The GO entries are hierarchically linked, thus allowing pooling and construction of cluster genes of crossed pathways. The bias in the gene representation (only a part of the genome is represented on the chip) could potentially lead to misleading conclusions. To evaluate the false discovery rate (FDR) for each potentially interesting GO category, we have used the high-throughput GoMiner that implements a correction for multiple comparisons testing (B. R. Zeeberg and H. Qin, in preparation). The statistical relevance of a candidate process found to be altered with drug resistance is therefore calculated in comparison with the overall processes that could be theoretically identified.
Quantitative RT-PCR. Real-time PCR was performed with a Light Cycler RNA SYBR Green kit (Roche Diagnostics). The reaction was performed in a 20-μl final volume with 0.150 μg of purified total RNA, 4 μl of PCR mix provided by the manufacturer, 4 μl of MgCl2 (25 mM), 2.5 μl of each primer (2 μM), and 0.4 μl of enzyme mix and diethyl pyrocarbonate-H2O. Specific primers were designed to produce unique bands for the 47 ABC transporters (Szakács et al., 2004). The reverse transcriptase reaction was performed at 55°C for 20 min. cDNA generated by the reverse transcriptase step was denatured at 95°C for 20 s. Amplification of the cDNA was achieved in 45 cycles of 95°C, 5 s; 58°C, 10 s; and 72°C, 13 s. Fluorescence was recorded during the elongation phase at 72°C.
Western Blots. After SDS-PAGE on 6% acrylamide SDS-PAGE gels (Invitrogen), samples (50 μg) were electroblotted on nitrocellulose membranes (Invitrogen). Membranes were saturated with a 10%, fat-free milk solution (Giant Foods, Inc., Landover, MD) and then incubated overnight in the presence of an anti-MRP2 primary antibody (M-8316; Sigma-Aldrich, St. Louis, MO) at 1:1000 dilution. A horseradish peroxidase-conjugated secondary antibody (anti-rabbit goat IgG (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) was applied for 1 h at 1:2000 dilution. Enhanced chemiluminescence reagents were purchased from Amersham Biosciences UK, Ltd. (Little Chalfont, Buckinghamshire, UK).
For glyceraldehyde-3-phosphate dehydrogenase and GST-Pi, 12% SDS-PAGE gels were used (Invitrogen). Primary antibodies were anti-glyceraldehyde-3-phosphate dehydrogenase RDI-TRK-5G4-6C5 (Research Diagnostics, Flanders, NJ) at 1:1000 dilution and anti-GST-P IgG1 at 1:1000 dilution (BD Biosciences PharMingen, San Diego, CA). The secondary antibody was goat-IgG anti-IgG mouse horseradish peroxidase-conjugated (IR-115-035-164; Jackson ImmunoResearch Laboratories Inc.) at 1:2000 dilution.
Results
Development of a Hybrid cDNA/Oligo ABC-ToxChip to Study Drug Resistance. The ABC-ToxChip, one section of which is shown in Fig. 1A, is a dedicated microarray designed for the study of the drug resistance of cancer cells. The chip includes three distinct sets of probes (Fig. 1A). The first set of probes targets 36 ABC transporters (70-mer oligonucleotides and cDNA probes). Specificity and sensitivity were ensured by targeting each transporter with multiple probes (Table 1). A second set of probes was included because multidrug resistance can be regarded as altered detoxification. The TOX probe set (Nuwaysir et al., 1999) covers known detoxification genes involved in response to estrogens, polycyclic aromatic hydrocarbons, peroxisome proliferators, DNA damage, and oxidant stress, as well as genes involved in apoptosis, cell cycle, tumor suppression, and signal transduction and transcription. To reduce the problem of sequence unreliability encountered with commercially available cDNA clones (Knight, 2001), the probes (200-500 base pairs) of the TOX-set were derived from sequence-verified plasmids originally compiled at the National Institute of Environmental Health Sciences. To minimize the risk of a bias in the analysis inherent in highly specialized microarrays, we completed the chip with a third probe set, the 18,000 probes (GEN-set) that target the human transcriptome (QIAGEN Operon). To minimize cross-hybridization, we verified the specificity of the cDNA and 70-mer oligonucleotide sequences by extensive in silico analysis (data not shown).

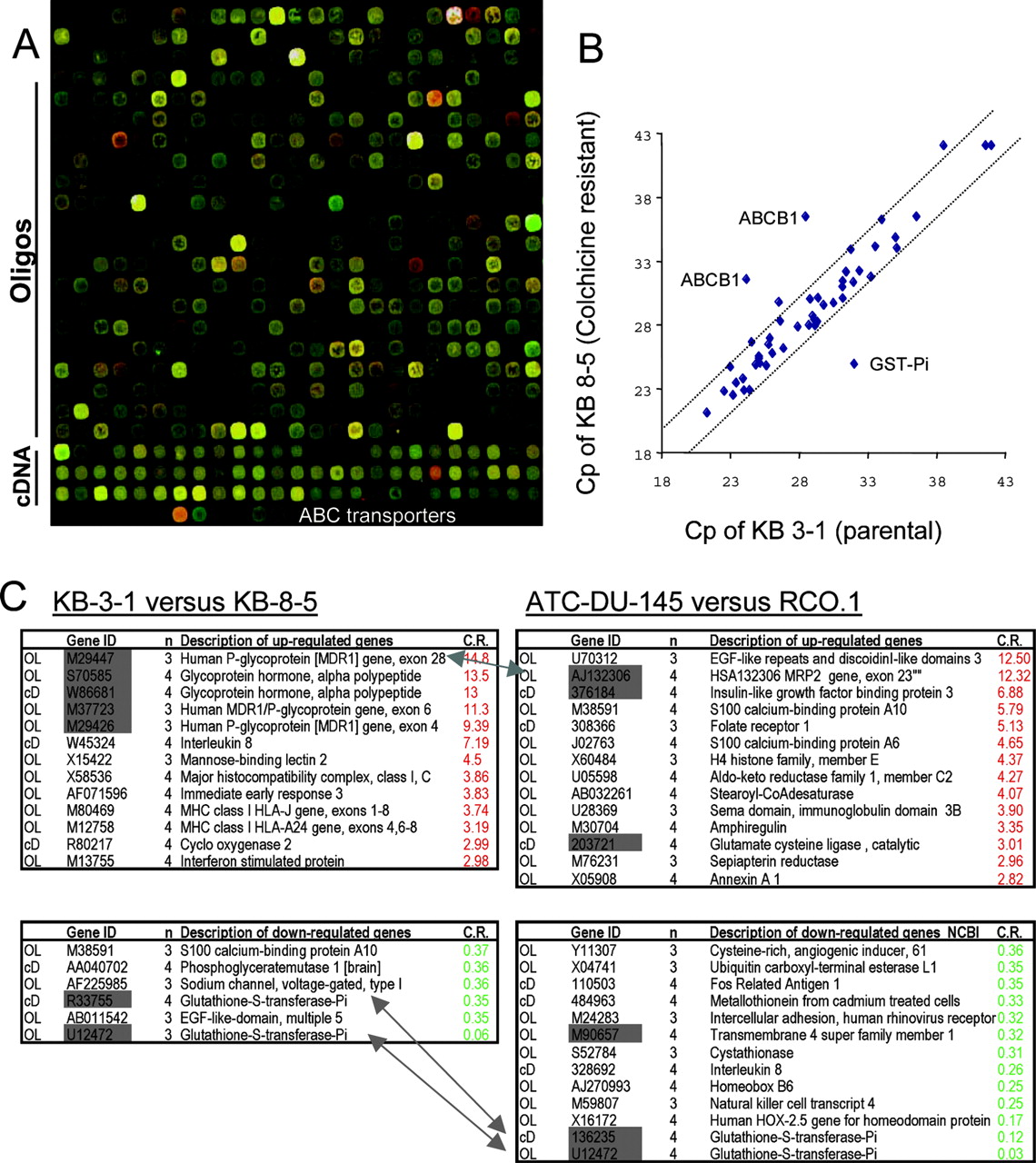
Creation and application of the ABC-ToxChip. A, details of a scan of the ABC-ToxChip. The section shown is one of the 32 blocks printed. The platform has 4 × 8 such blocks, each containing 625 (25 × 25) probe locations, yielding in total 20,000 locations available for printing. The upper part of the block contains the oligonucleotide set, and the lower part bears the cDNA-probe detoxification set. The probes specifically matching the ABC transporters are printed in the lowest row. B, quantitative RT-PCR to follow transcriptional changes of ABC transporters and GST-Pi. Crossing point values of amplification for 47 ABC transporters and GST-Pi in the two samples are shown in a scatter plot. Deviations greater than 2 cycles are considered significant. Results demonstrate activation of MDR1 and down-regulation of GST-Pi with colchicine resistance. C, significant outliers (>2.7-fold) identified by the microarray experiments. ABC transporters occur as major outliers in both analyses, suggesting their strong contribution to the MDR phenotype. Genes for which the variation was confirmed by more than one probe are indicated in gray. Arrows indicate the most drastic changes discussed in the text (i.e., changes in GST and ABC transporters). OL and cD refer to oligonucleotide and cDNA probes, respectively. n defines the number of experiments (out of four replicates) in which gene expression was found to be significantly altered. C.R. (calibrated ratio) indicates the -fold difference in expression between the resistant cell line versus the parental cell line.
Of the 18,398 probes represented by 16,329 oligonucleotides and 2069 cDNA that matched a valid gene reference, 11,813 were annotated with HUGO references, using the Source (Diehn et al., 1993) (http://source.stanford.edu/cgibin/source/sourceSearch) and Matchminer databases (Bussey et al., 2003) (http://discover.nci.nih.gov/matchminer). The HUGO identifiers were subsequently used as input for analysis by High-Throughput GoMiner. The cDNA, or detoxification, set includes 1394 probes with valid GO annotations, of which 1152 are also represented in the GEN-set. Analysis of the distribution of gene ontology entries confirmed that the cDNA set is indeed enriched in genes related to detoxification processes. For example, genes related to drug stress and DNA repair are highly represented in the cDNA TOX set (1.3 and 3.3% of the total genes, respectively), compared with the GEN-set (0.3 and 0.6%, respectively).
Validation of the ABC-ToxChip Using an Established Model of Resistance. To validate the chip with respect to markers of resistance and sensitivity to colchicine, we compared two cell lines using the ABC-ToxChip. KB-8-5 cells were derived from parental KB-3-1 cells by selection in colchicine (Akiyama et al., 1985). RNA samples from the two lines were reverse-transcribed, labeled with Cy3/Cy5, and hybridized to four ABC-ToxChip slides. After filtering the data to find the most reliably differentially expressed genes (with a statistical type I error of 5%), we identified 241 significant differences (Table S1). Among the four cross-labeled experiments, 98 hits (40.6%) occurred in all four replicate scans. The rest (59.4%) were significant in only three of the four replicate slides.
In total, we identified 13 differentially expressed genes associated with colchicine resistance with ratios above 2.7 (Fig. 1C). Among the 20,000 genes, ABCB1/P-gp showed the highest difference (10-fold). Closely related members of the same subfamily (ABCB2-3-4) showed only moderate (about 2-fold) changes, presumably caused by cross-hybridization, confirming the improved specificity of our dedicated microarray for ABC transporters. To verify the patterns of ABC expression detected by the microarray analysis, we measured the expression of 47 ABC transporters by real-time RT-PCR. The analysis confirmed that ABCB1 (MDR1) was strongly up-regulated, whereas the expression of the other members was virtually unchanged (Fig. 1B). Comparison of KB-3-1 cells with KB-C1, a derivative showing extreme resistance (1 μg/ml) to colchicine showed the highest change in expression of the same genes (log2 ratio of 6.32 and -4.2 for ABCB1/MDR1 and GST-Pi, respectively; data not shown).
Increased expression levels of MDR1 (ABCB1) and the glycoprotein hormone subunit (GPH) were revealed by several independent probes (M29447, M37723 and S70585, W86681 for GPH and ABCB1, respectively). The list of other up-regulated genes includes interleukin-8, MHC-I, COX-2, and the interferon-stimulated protein, whereas GST-Pi, calcium-binding protein A10, voltage-gated sodium channel, and epidermal growth factor-like domain 5 are markedly down-regulated in KB-8-5 cells compared with the parental cell line. We used the information provided by HUGO to process the ab initio and hypothesis-free GoMiner algorithm to detect biological processes that are represented by the differentially expressed genes and could possibly explain mechanisms leading to colchicine resistance.
Among 241 statistically significant outlier genes (see Materials and Methods), we identified 125 unique HUGO entries (52%). Because the dedicated design of the microarray can introduce a bias in analysis, with, for example, an over-represented class of detoxifying genes printed, we evaluated the significance of the results in the light of the global representation of the GO ontology annotation entries. The GoMiner analysis indicates that the `antigen presentation' and `carbohydrate metabolism' GO categories are enriched in genes that exhibit altered expression in the colchicine-resistant cell line (P < 0.0001). In Table S2, we present the statistically significant biological processes that can be associated with colchicine resistance. In Table S3, we present the genes that are members of the altered gene ontology categories related to antigen presentation, carbohydrate metabolism, transport, and oxidoreduction.
Application of the ABC-ToxChip to Study 9-Nitro-camptothecin Resistance. We next turned our attention to RCO.1, a prostate cell line selected in 9NC, for which mechanisms leading to the resistant phenotype have not been fully elucidated. This cell line was generated by selecting parental DU-145 cells in 0.1 μM 9NC (Urasaki et al., 2001). RCO.1 presents a unique profile of multidrug resistance with a selective resistance to camptothecin analogs NB-506 (an inhibitor of topoisomerase I) and cisplatin, whereas being sensitive to drugs known to be exported by ABCB1-MDR1 or ABCC1-MRP1 (Chatterjee et al., 2001). Several laboratories have attempted to explain this atypical profile of resistance: a mutation of topoisomerase I, preventing the binding of camptothecin analogs (Chatterjee et al., 2001) and alteration of the apoptotic pathway (Chatterjee et al., 2001; Reinhold et al., 2003) have been proposed. However, an exhaustive analysis of ABC transporters or genes involved in detoxification has not been reported so far for those cells. To explore the role of ABC transporters and other detoxifying genes in 9NC resistance, we compared the mRNA expression profiles of DU-145 and RCO.1 cells using the ABC-ToxChip. Total cellular RNA was extracted from both cell lines. Because the phenotype of acquired 9NC resistance is stable without the pressure of selection, RNA was collected from cells cultured in drug-free medium. Microarray experiments were performed as described previously for the KB cells. To control for labeling differences, reactions were carried out in quadruplicate, and the fluorescent dyes were switched to minimize dye bias.
The genes most significantly overexpressed in RCO.1 are shown in Fig. 1C and in Table S4. Among the 20,000 genes, ABCC2-MRP2 shows one of the highest differences. First named the canalicular multispecific organic anion transporter, ABCC2-MRP2 is a 190-kDa phosphoglycoprotein localized in the canalicular (apical) membrane of hepatocytes. It is involved in the transport of organic anions, including sulfated and glucuronidated bile salts. Overexpression of MRP2 has been suggested to confer resistance to anticancer drugs such as cisplatin, anthracyclines, and methotrexate, and animal models have shown reduced hepatic transport of camptothecins (Horikawa et al., 2002).
γ-Glutamylcysteine synthetase (GCS), also known as glutamate cysteine ligase, a key enzyme in glutathione metabolism, was coordinately up-regulated with MRP2, in keeping with the finding that MRP2 exports glutathione conjugates (Paulusma et al., 1996). In contrast, despite its general up-regulation in cancer cells, GST-Pi, which catalyzes the conjugation of glutathione to electrophilic carcinogens, was significantly down-regulated in RCO.1 cells, as it was in the colchicine-selected KB cells.
Additional changes that might contribute to the pattern of drug resistance in RCO.1 cells were also observed. Up-regulation of various histones in the selected cells may provide a means for the cells to adapt to the 9NC-mediated DNA insult. Further changes associated with drug resistance include reduction in the tumor-associated antigen L6 [transmembrane (TM) superfamily 4 or TM6]. TM6 has also been found to be down-regulated in cisplatin-resistant cells (KB/cDDP) (Higuchi et al., 2003), indicating that the loss of TM6 expression is potentially associated with resistance to camptothecin and cisplatin. Of the genes presented in Table S4, GST-Pi, NKT4, and interleukin-4 were underexpressed in RCO.1 cells. The same was seen in an independent microarray analysis (Table 2), which failed, however, to detect the overexpression of ABCC2-MRP2.
Differentially expressed genes identified by two independent microarray analyses (ATC-DU-145 versus RCO.1)
Although 85 of the 125 outliers (68%) have a HUGO reference, the GoMiner algorithm did not suggest specific biological processes to be significantly linked to camptothecin resistance (see Tables S5 and S6 for the biological processes and their associated genes linked to camptothecin resistance.)
We next sought to determine whether the differential expression of ABCC2-MRP2 and GST-Pi could be confirmed by quantitative RT-PCR. The results show that among the 47 ABC transporters, ABCC2-MRP2 is overexpressed in the resistant cell line, whereas expression of the other members is unchanged (Fig. 2A). Because mRNA levels may not reflect protein expression (as a result of modulation of translation or inhibition of protein degradation), we analyzed expression of MRP2 and GST-Pi at the protein level. Figure 2C shows that protein expression levels of these two genes follow the pattern predicted by the RNA analyses: in the RCO.1 cells, GST-Pi is down-regulated and ABCC2-MRP2 is up-regulated compared with the parental line.

Overexpression of ABCC2 and down-regulation of GST-Pi with 9NC selection. A, confirmation of the overexpression of ABCC2 and down-regulation of GST-Pi with 9NC selection by quantitative RT-PCR. Crossing point values of amplification for 47 ABC transporters and GST-Pi in the two samples are shown in a scatter plot. B, illustration of the role of ABCC2, GST, and GCS in the glutathione-related detoxification pathway. The concentration of glutathione is regulated by its metabolism (GSH-GSX, GSH-oxidized GSH) and biosynthesis, catalyzed by GST and GCS, respectively, and the activity of ABCC2. C, correlation between the mRNA and protein levels. The symbols S and R correspond to the sensitive (ATC-DU-145) and resistant (RCO.1) cell lines, respectively. The signal for GST-Pi in the resistant cell line was below the level of detection.
Discussion
In this report, we present a new microarray, the ABCToxChip, for analysis of multifactorial drug resistance. We designed and synthesized probes to match 36 of the 48 ABC transporters and printed them in an array enriched in genes involved in detoxification, as well as a general set of 18,000 human gene probes. To increase at the same time sensitivity and specificity for genes involved in drug resistance, we combined on the same chip both cDNA and oligonucleotide probes. This approach provides an example of how existing microarray platforms may be modified to target a gene family with high sensitivity and specificity to study multidrug resistance of cancer. Using the ABC-ToxChip in the present study, we found, consistent with expectation from previous observations, that ABCB1-MDR1 is overexpressed in colchicine-resistant KB-8-5 cells. KB-8-5 cells express ABCB1-MDR1 at a level comparable with that found in clinical samples. We had made several earlier attempts to identify the molecular signature of colchicine resistance, using a 9000-clone cDNA microarray platform (UniGemV2; Advanced Technology Center, National Institutes of Health, Gaithersburg, MD). The UniGemV2 chip failed to identify overexpression of ABCB1-MDR1; ABCB2, a protein closely related in sequence to ABCB1 was the main outlier (data not shown). ABCB2-TAP1 is a component of the endoplasmic reticulum transport system for peptide antigen presentation and is not believed to play a role in the efflux of cytotoxic compounds. According to the current study, it is not actually overexpressed in KB-8-5 cells and probably appeared so in the previous work because of cross-hybridization of its oligonucleotide probe with ABCB1.
Our analysis of a prostate cancer cell line (RCO.1) indicated a possible role of ABCC2/MRP2 in 9NC resistance. Previous studies have shown that the resistance of RCO.1 cells could also be explained by a mutation in the Topoisomerase 1 gene (Urasaki et al., 2001) and/or by a defect in apoptosis pathways (Chatterjee et al., 2001; Reinhold et al., 2003), suggesting that resistance to camptothecin is multifactorial. These previous attempts to elucidate mechanisms leading to camptothecin resistance included microarray analyses of parental DU145 cells and RCO.1 cells (Reinhold et al., 2003), using a UniGem microarray. Despite the high expression of ABCC2/MRP2 in the resistant cells (Fig. 1C), its overexpression was not detected in these earlier studies. Together, these findings suggest that the specific targeting of ABC transporters using a dedicated, custom-designed microarray may improve on the specificity and sensitivity of earlier generation microarrays.
Shortcomings typical of microarray platforms include imperfect clone annotation and the lack of specificity of probes targeted at overlapping or homologous sequences of closely related proteins, such as ABCB1 and ABCB2. Those considerations prompted us to create the ABC-ToxChip as a platform dedicated to study of drug resistance and related phenomena. To ensure specificity, we designed probes uniquely matching the target transporters. The ABC-ToxChip analysis of cells selected in colchicine and in 9NC demonstrates the elevated expression of ABCB1 and ABCC2, respectively. Because gene expression levels obtained by even the most carefully designed microarrays must be validated by independent methods, we also designed specific primers for the human ABC transporters (Szakács et al., 2004). Quantitative RT-PCR data confirmed the pattern of ABC transporter expression and suggested that there are no further ABC transporters differentially expressed in the cells analyzed in this study. Through the systematic analysis of ABC transporters and other genes involved in detoxification, our data provide novel information about the effect of 9NC selection on gene expression.
Although the GoMiner algorithm did not identify major biological processes linked to 9NC resistance, the list of differentially expressed genes may provide some insight into mechanisms underlying (or accompanying) resistance. An interesting example is Radixin, which is overexpressed in the 9NC selected cell line. Radixin, which belongs to the ERM (Ezrin-Radixin-Myosin) protein complex, is involved in localization of integral membrane proteins. Radixin -/- mice have Dubin-Johnson-like symptoms (Kikuchi et al., 2002), because MRP2 is not properly localized to the plasma membrane.
The coordinated expression of phase II (conjugating) and phase III (efflux) systems has been shown to improve cellular detoxification (Morrow et al., 2000). We found significant changes in the expression of two glutathione metabolism-related genes. GST-Pi is involved in coupling electrophilic drugs to reduced glutathione, and GCS is the rate-limiting enzyme for glutathione synthesis. Neither GST-Pi mRNA nor the protein is detectable in the resistant cell line. This observation is striking given the high GST-Pi levels found in several resistant tumors and cell lines (Tew, 1994; Liu et al., 2001). In contrast, GCS is overexpressed in the camptothecin-resistant RCO.1 cells, as shown by both oligonucleotide and cDNA probes (Fig. 1C). The changes in GST-Pi, GCS, and ABCC2 expression suggest a putative “metabolic switch” necessary for resistance. As shown in Fig. 2B, these enzymes play roles in the pathway of glutathione-mediated detoxification. Two further genes belonging to the “cysteine metabolism” ontology group (Table S4) consistently show differential expression (cystathionine γ-lyase and glutamate cysteine-ligase; Table S6).
Elevated expression of ABCG2-MXR (Brangi et al., 1999) has been observed in DX-8951f, a derivative of the human ovarian cancer cell line A2780 selected for resistance by exposure to an analog of camptothecin (van Hattum et al., 2002). Our results do not show elevated ABCG2 expression in RCO.1 cells. They do, however, suggest involvement of ABCC2 in the resistant phenotype. Consistent with our findings, previous studies have reported ABCC2 as a detoxifying transporter for camptothecin in animal models (Arimori et al., 2003) and cell lines transfected with antisense to ABCC2 (Koike et al., 1997). Because resistance of RCO.1 to 9NC was shown to be partially mediated by a mutation in topoisomerase I, we speculate that the elevated expression of ABCC2 may play a role during the initial steps of the selection process. This early adaptation may provide the background for evolution of further resistance mechanisms, such as mutation or down-regulation of topoisomerase 1 (Urasaki et al., 2001).
Retroviral transfer of ABCC1 (MRP1) has been shown to result in decreased intracellular glutathione levels and increased sensitivity to BSO (Rappa et al., 2003). Furthermore, in vitro reversal of MRP1-mediated resistance and in vivo potentiation of the cytotoxicity of doxorubicin in MRP1-overexpressing tumors by BSO have previously been reported (Rappa et al., 2003). In addition, consistent with the capacity of ABCC2 to export glutathione conjugates (Konig et al., 1999), RCO.1 cells have proved hypersensitive to BSO treatment (data not shown). It is possible that MRP2 and GCS are up-regulated in a coordinated way as the cells adapt to the cytotoxic stress. Because maintenance of glutathione levels is critical to survival of the cells, overexpression of GCS may represent an adaptive response compensating for the loss of intracellular glutathione.
In conclusion, we designed and printed a novel ABC-Tox-Chip and then used it to assess the molecular concomitants of drug resistance in two cell culture model systems. In both, we found that two of the ABC transporters are prominently overexpressed (in relation to 18,000 other genes) at the level of transcription. These results reinforce our focus on the expression of ABC transporters as specific markers of acquired drug resistance. The ABC-ToxChip should be a helpful tool for assessing the roles of ABC transporters. It should also be helpful in studies of the linkage between expression of the ABC transporters and expression of other detoxification genes in drug resistance and in various pathophysiological processes. We expect it to be useful not only for cell culture models but also for clinical samples.
Acknowledgments
We thank Dr. Pantazis for the generous gift of the RCO.1 cell lines. We are also grateful to George Leiman for editorial assistance.
Footnotes
-
This work was supported by the National Institutes of Health.
-
J.-P.A. and G.S. contributed equally to this work.
-
ABBREVIATIONS: MDR, multidrug resistance; RT-PCR, reverse transcription-polymerase chain reaction; ABC, ATP-binding cassette; GST, glutathione S-transferase; MRP, multidrug resistance-associated protein; CPT, camptothecin; 9NC, 9-nitro-camptothecin; PCR, polymerase chain reaction; GO, gene ontology; Cy, cyanine; PAGE, polyacrylamide gel electrophoresis; GCS, γ-glutamylcysteine synthetase; TM, transmembrane; BSO, l-buthionine-[S,R]-sulfoximine; HUGO, Human Genome.
-
↵s⃞ The online version of this article (available at http://molpharm.aspetjournals.org) contains supplemental Tables S1-S6.
-
↵1 Current address: Laboratoire de Biologie Moléculaire et Structurale, Centre National de la Recherche Scientifique, Gif Sur Yvette, France.
- Received July 14, 2004.
- Accepted August 30, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}