


Abstract
The aryl hydrocarbon receptor (AhR) is a ligand-activated transcription factor responsible for mediating the cellular response to the toxic compound 2,3,7,8,-tetrachlorodibenzo-p-dioxin. An essential role for the AhR in cellular biology has been established previously, but no high-affinity endogenous ligand has yet been identified. We have confirmed the presence of a putative endogenous ligand(s) in CV-1 cells through transient transfection with various cytochrome P450 isoforms. Expression of cytochromes P450 1A1, 1A2, or 1B1 reduced AhR-mediated luciferase reporter activity, whereas cytochrome P450 2E1 exhibited no significant effect. Studies with 2,4,3′,5′-tetramethoxystilbene, a potent and specific inhibitor of cytochrome P450 1B1, was able to partially block cytochrome P450 1B1-mediated reduction in reporter gene activity. These results provide evidence of the existence of a possible feedback mechanism in which AhR-regulated cytochromes P450 from the CYP1A and CYP1B families are able to metabolically alter putative endogenous ligand(s). Several experiments were performed to provide initial characterization of these putative endogenous ligands, including electrophoretic mobility shift assay analyses, which demonstrated that these ligands directly activate the AhR. Soluble extracts from various C57BL/6J and Ahr-null mouse tissues were also analyzed for the presence of AhR activators. Studies revealed that Ahr-null mouse lung tissue had a 4-fold increase in AhR-mediated reporter activity in cells. Quantitative polymerase chain reaction analysis revealed that lung tissue exhibits relatively high constitutive CYP1A1 mRNA levels. These results suggest that there is an autoregulatory feedback loop between the AhR and cytochrome P450 1A1 in mouse lung.
The aryl hydrocarbon receptor (AhR) is a ligand-activated basic helix-loop-helix transcription factor and member of the basic helix-loop-helix/Per-Arnt-Sim family of DNA binding regulatory proteins (Burbach et al., 1992; Ema et al., 1992; Schmidt and Bradfield, 1996). In the absence of ligand, the Ah receptor resides in the cytosol in an oligomeric 9 S protein complex. Contained in this core complex are the AhR ligand-binding subunit, a dimer of the 90-kDa heat shock protein, and a single molecule of the immunophilin like X-associated protein 2 and p23 (Chen and Perdew, 1994; Meyer et al., 1998; Hollingshead et al., 2004). Ligand binding induces receptor translocation into the nucleus followed by dissociation of the 90-kDa heat shock protein. Once in the nucleus the ligand-activated AhR can dimerize with aryl hydrocarbon receptor nuclear translocator (ARNT), forming a high-affinity DNA binding complex capable of binding specific target DNA sequences known as dioxin-responsive elements (DRE) in the regulatory region of responsive genes (Fukunaga and Hankinson, 1996). Interaction with these response elements results in an alteration of gene expression. Genes transcriptionally regulated by the AhR are primarily involved in foreign chemical metabolism and include the xenobiotic-metabolizing cytochrome P450 enzymes from the 1A and 1B families (Gonzalez et al., 1984; Sutter et al., 1994). Additional AhR-regulated genes include glutathione transferase Ya subunit (Telakowski-Hopkins et al., 1988), NAD(P)H-menadione oxidoreductase (Favreau and Pickett, 1991), UDP-glucuronosyltransferase (Lamb et al., 1994), aldehyde-3-dehydrogenase (Vasiliou et al., 1996), and prostaglandin endoperoxide H synthase-2 gene (Vogel et al., 1998).
Before any of the DRE-responsive genes can be activated, the AhR signal transduction cascade must be initiated through ligand binding. Many types of compounds, ranging from widespread and potentially toxic synthetic environmental contaminants to dietary metabolites formed in the acidic environment of the stomach, can serve as regulators of the AhR. Halogenated aromatic hydrocarbons and polycyclic aromatic hydrocarbons are two of the most potent classes of AhR ligands known, and they include such carcinogenic compounds as polychlorinated biphenyls, polychlorinated dibenzo-p-dioxins and benzo[a]pyrene. These compounds are produced through various industrial processes, including chlorine bleaching of wood pulp, waste incineration, incomplete combustion of fossil fuels, metal production, and synthesis of organochlorine products (Denison and Heath-Pagliuso, 1998). In addition to synthetic exogenous ligands, it is well documented that dietary derived indole-containing compounds, including indole-3-carbinol (Bjeldanes et al., 1991), oxidized carotenoids such as canthaxanthin and astaxanthin (Gradelet et al., 1997), and heterocyclic amines produced during the cooking of meat (Kleman et al., 1992), can all function as AhR ligands. Perhaps even more significant is the formation of indolo[3,2-b]carbazole, a metabolite of indole-3-carbinol that is produced via acid condensation in the stomach and that is capable of functioning as a potent AhR ligand, possessing an affinity for the receptor only 1 order of magnitude weaker than the highly toxic carcinogen 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) (Bjeldanes et al., 1991; Perdew and Babbs, 1991). Any toxicological or biological significance of long-term exposure to these rapidly metabolized diet-derived ligands of the AhR remains to be determined.
An important question that remains unresolved is the identity of key endogenous regulators of the AhR. Several endogenous compounds possessing low affinity for the receptor have been identified, including the heme breakdown products bilirubin and biliverdin (Sinal and Bend, 1997; Phelan et al., 1998); 7-ketocholesterol, a cholesterol derivative (Savouret et al., 2001); several arachadonic acid metabolites, including lipoxin A4 (Schaldach et al., 1999); and various prostaglandins (Seidel et al., 2001). However, no high-affinity endogenous AhR ligand has yet been characterized that can activate the AhR at physiologically relevant conditions, despite the accumulation of evidence supporting their existence. For example, activation of AhR-dependent processes in the absence of exogenously added ligand has been reported with animal cells in culture (Sadek and Allen-Hoffmann, 1994; Ma and Whitlock, 1996; Weiss et al., 1996; Chang and Puga, 1998). Assuming that AhR activation requires ligand, these observations can be interpreted as indirect evidence for the existence of an endogenous ligand(s), present at sufficient concentrations to lead to significant levels of AhR activation. Quite possibly the most conclusive evidence supporting an endogenous biological role for the receptor has been derived from studies using Ahr-null mice. Three independent laboratories have each generated Ahr-null mice, and although differences in phenotype have been reported, all of these animals share certain common defects. In general, these mice displayed multiple hepatic defects, including a decrease in liver size and weight as a result of a decrease in the cytoplasmic volume of hepatocytes along with mild bile duct fibrosis (Fernandez-Salguero et al., 1995; Lahvis et al., 2000). Other aberrations included compromised immune system function due to a decrease in the number of splenocytes (Fernandez-Salguero et al., 1995), reproductive defects (Abbott et al., 1999), and anomalies of the eye and kidney, including persistent neonatal vascular structures (Lahvis et al., 2000). In addition to the aforementioned defects, mice harboring Ahr-null alleles exhibited decreased constitutive expression of the xenobiotic-metabolizing enzyme cytochrome P450 1A2, along with a complete loss of cytochrome P450 1A1 induction normally seen in response to dioxin exposure (Lahvis and Bradfield, 1998). This specific phenotype has resulted in benzo[a]pyrene no longer being able to mediate skin carcinogenesis in these animals (Shimizu et al., 2000). Taken together, these observations provide support for the existence of high-affinity endogenous AhR ligands. However, it is also possible that the AhR is activated in a ligand-independent manner. Nevertheless, it is probable that the AhR plays a crucial biological role in normal cellular metabolism and that one or more high-affinity endogenous ligands exist to modulate the timing, duration, and magnitude of its function in the cell. Elucidating the structure of such a ligand(s) would enable a biological role for this enigmatic orphan receptor to be more precisely determined. Subsequently, the significance of excessive Ah receptor activation by environmental compounds could be more thoroughly evaluated and new molecular targets for therapeutic drugs might emerge. In this report, we establish the presence of putative high-affinity endogenous Ah receptor ligand(s) isolated from CV-1 cells. In addition, we establish that the compounds are also a substrate for AhR-regulated cytochromes P450, and we provide evidence for an autoregulatory feedback loop between the AhR and cytochrome P450 1A1 that is most active in lung.
Materials and Methods
Reagents. Glycerol, acrylamide, and bisacrylamide were purchased from Research Organics (Cleveland, OH). Optima grade high-purity organic solvents meeting American Chemical Society standards were purchased from Thermo Fisher Scientific Co. (Waltham, MA), and they were used wherever applicable throughout the course of these studies. Glacial acetic acid and powdered anhydrous sodium sulfate, manufactured by EMD Chemical, were obtained from VWR (West Chester, PA). Anhydrous dimethyl sulfoxide (DMSO) at +99.9% purity was purchased from Sigma-Aldrich (St. Louis, MO). TCDD was a generous gift from Steve Safe (Texas A&M University, College Station, TX).
Plasmids and Bacterial Strains. The construct pcDNA3-βmAhR was used to express the murine AhR, whereas pGudLuc 6.1 was used for DRE-driven luciferase reporter gene expression in cells as described previously (Murray et al., 2005). β-Galactosidase was expressed using the pDJM2-β-gal construct, and it served as a control for transfection efficiency (Adra et al., 1987). Ectopic expression of cytochrome P450 isoforms 1A1, 1A2, 1B1, and 2E1 in CV-1 cells was achieved with the expression vectors pCMV4-hCYP1A1 and pCMV4-hCYP1A2 obtained from Dr. Robert Tukey (University of California, San Diego, La Jolla, CA), pRcCMV-hCYP1B1 obtained from Dr. William F. Greenlee (Chemical Industry Institute of Toxicology, Research Triangle Park, NC), and pCI-hCYP2E1 supplied by Judy Raucy (Puracyp Inc., Carlsbad, CA). The hCYP2E1 cDNA was subcloned from the EcoRI site of pCR2.1 to pCI. Plasmids were transformed into Escherichia coli DH5α-competent bacterial cells (PGC Scientifics, Frederick, MD). Transformed bacteria were grown overnight in Luria broth supplemented with the appropriate antibiotic for selection.
Cell Lines and Cell Culture. The CV-1 cell line was obtained from the American Type Culture Collection (Manassas, VA). Trypsin-EDTA, PBS, α-modified minimal essential media (α-MEM), penicillin, and streptomycin were all obtained from Sigma-Aldrich (St. Louis, MO). Fetal bovine serum was purchased from Hyclone Laboratories (Logan, UT). Opti-MEM was purchased from Invitrogen (Carlsbad, CA). CV-1 cells were grown in (α-MEM) supplemented with 10% fetal bovine serum (v/v), 100 IU/ml penicillin, and 0.1 mg/ml streptomycin at 37°C in a humidified atmosphere containing 6% CO2, 94% room air.
Transient Transfection, Luciferase, and β-Galactosidase Assays. Transfection of CV-1 cells was performed in six-well tissue culture plates, with a total of 2.5 μg of plasmid DNA per well and delivered using Lipofectamine reagent (Invitrogen) according to the method described by the manufacturer. Transient transfections used 500 ng of pcDNA3/βmAhR; 200 ng of pGudLuc 6.1, a DRE-driven reporter construct; and pDJM-βgal, the β-galactosidase control plasmid, along with varying amounts of a cytochrome P450 expression vector or control plasmid. Upon completion of the transfection, Lipofectamine complexes were removed and replaced with α-MEM containing 10% fetal bovine serum. Twenty-four hours after transfection, cells were thoroughly rinsed with PBS and lysed in 1× cell culture lysis buffer (2 mM trans-1,2-diaminocyclohexane-N,N,N′,N′-tetraacetic acid, 2 mM dithiothreitol, 10% glycerol, and 1% Triton X-100). Lysates were centrifuged at 18,000g for 15 min. Cytosol was assayed for luciferase activity using the Promega luciferase assay system (Promega, Madison, WI) as specified by the manufacturer. Light production was measured for 15 s at room temperature using a TD-20e luminometer (Turner Designs, Sunnyvale, CA). Cytosolic protein concentration was determined using the bicinchoninic acid assay (Pierce Chemical, Rockford, IL). β-Galactosidase assays were performed and spectrophotometrically monitored at 420 nm using the Promega β-galactosidase assay system (Promega). Luciferase activity was expressed relative to β-galactosidase activity or protein concentration.
Inhibition of CYP1B1 by 2,4,3′,5′-Tetramethoxystilbene. CV-1 cells were grown to ∼80% confluence in six-well tissue culture plates, and then they were transfected with plasmid DNAs using the Lipofectamine (Invitrogen) method, as described by the manufacturer. Control cells were transfected with 500 ng of pcDNA3-βmAhR, 250 ng of pDJM2-β-gal, 200 ng of pGudLuc 6.1 and pCI vector, for a total of 1.5 μg of DNA per well. Experimental cells received the same transfected DNA profile except pCI vector was replaced with 400 ng of pRcCMV-CYP1B1. Transfected cells were allowed 16 h of recovery in complete medium before being dosed with concentrations of 2,4,3′,5′-tetramethoxystilbene (TMS), ranging from 50 nM to 10 μM, for an additional 8 h before harvest. Cells were thoroughly rinsed with PBS before lysis in 1× cell culture lysis buffer. Cell lysates were centrifuged at 18,000g for 15 min. Cytosol was assayed for luciferase activity, and activity was expressed as relative light units per units of β-galactosidase activity.
Initial Ligand Characterization. C18 Solid Phase Extraction columns (Phenomenex, Torrance, CA) were initially used in attempts to concentrate the putative AhR ligand from CV-1 cells. Eventually, solid phase extraction methods were abandoned in favor of liquid extraction methods using high-purity Optima grade organic solvents (Fisher Scientific Co.). Performing multiple (two to three) extractions of CV-1 cytosol in series with an excess (2.0–2.5 volumes) of organic solvent proved to be the most effective method for extracting the AhR-modulating compounds. Cytosol was mixed together with organic solvent [hexane/ethyl acetate, 1:1 (v/v)] in a 15- × 130-mm round-bottomed glass centrifuge tube. This solvent was used for all extractions. The tubes were capped with a solvent-washed silicon stopper, and the mixtures were vortexed vigorously for several minutes. To facilitate separation of the aqueous and organic phases, the samples were centrifuged at 1600g for 10 min using a Beckman S4180 rotor in a Beckman Allegra 21 tabletop centrifuge (Beckman Coulter, Fullerton, CA) at 15°C. After extraction, the organic phase was removed and dried under reduced pressure in a CentriVap (Labconco Corporation, Kansas City, MO) under argon gas sparge or by hand under a continuous gentle stream of filtered argon gas. Immediately upon reaching dryness, the samples were carefully resolubilized in DMSO, and they were assayed using the HepG2 40/6 reporter cell line, which contains a stably integrated pGudLuc 6.1 vector (Long et al., 1998). Each bar represents the mean of three separate determinations ± S.D.
Electrophoretic Mobility Shift Assays. DRE-specific electrophoretic mobility shift assays were performed using in vitro-translated AhR and ARNT proteins. Expression vectors for these proteins were translated using a TNT-coupled transcription and translation rabbit reticulocyte lysate kit (Promega). Varying amounts of CV-1 cytosol were extracted with hexane/ethyl acetate [1:1 (v/v)], stripped of residual water through incubation with anhydrous sodium sulfate, evaporated to completion under reduced atmospheric pressure using a speed vac, and resuspended in 1.5 μl of DMSO. Proteins for the transformation reactions were mixed together at a 2:1 M ratio in HEDGE buffer (25 mM HEPES, 1 mM EDTA, 10 mM sodium molybdate, and 10% glycerol, pH 7.5), followed by addition of either DMSO-solubilized organic extract of CV-1 cytosol or TCDD. All transformation assays were incubated for 90 min at room temperature, followed by the addition of oligonucleotide buffer [42 mM HEPES, 0.33 M KCl, 50% glycerol, 16.7 mM dithiothreitol, 8.3 mM EDTA, 0.125 mg/ml CHAPS, and 42 ng/μl poly(dI:dC)]. After 15 min, 200,000 cpm of 32P-labeled wild-type DRE was added to each reaction for an additional 15-min incubation. A portion of each sample was removed and electrophoresed on a 6% nondenaturing polyacrylamide gel. Wild-type DRE oligonucleotides made up of the sequence 5′-GATCTGGCTCTTCTCACGCAACTCCG-3′ and 3′-ACCGAGAAGAGTGCGTTGAGGCCTAG-5′ were gifts from Dr. M. S. Denison (University of California, Davis, CA).
Proteinase K Treatment. Approximately 2.6 mg of CV-1 cytosol was incubated in the presence of proteinase K [1:1 (w/w)] for 90 min at room temperature. An equal amount of CV-1 cytosol, serving as a control, was left untreated, and it was incubated under the same conditions. Upon completion of proteinase K digestion, 20-, 30-, and 40-μg aliquots of cytosol were removed from both reactions. These samples were mixed with an equal volume of 2× Tricine sample buffer, heated at 95°C for 5 min, and resolved by Tricine SDS-polyacrylamide gel electrophoresis (3.96% stacking gel/7.92% running gel) to confirm complete digestion of cytosolic protein. The remaining cytosol from both the treated and untreated reactions was divided into 125-, 250-, and 500-μg aliquots. Samples were then extracted with 3 volumes of hexane/ethyl acetate [1:1 (v/v)]. The organic phase was removed, and the extract was dried over anhydrous Na2SO4, followed by evaporation to completion under argon gas. The extracts were then dissolved in a minimal volume of DMSO and applied to HepG2 40/6 cells for bioassay analysis. After a 6-h incubation, cells were harvested and assayed for luciferase reporter gene activity. Results are displayed in bar graph, and each bar represents the mean of four separate determinations ± the standard deviation.
Mice. Adult wild-type C57BL/6J mice (∼2 month) were purchased from The Jackson Laboratory (Bar Harbor, ME). Ahr-null mice in a C7BL/6J background were obtained from Dr. Bradfield (McArdle Laboratory for Cancer Research, University of Wisconsin-Madison Medical School, Madison, WI). The mice had free access to water and diet, and they were maintained in a temperature- and light-controlled facility. The standards for animal use and care set by The Pennsylvania State University's animal research program were followed for all experiments. Carbon dioxide inhalation method was used to sacrifice the mice. Tissue samples were collected, frozen in liquid nitrogen, and stored at –80°C until processed further.
Murine Tissue Extraction. Mouse tissues were mechanically homogenized in buffer and cytosolic extraction-isolated as described previously (Hollingshead et al., 2006). The protein concentration of cytosolic preparations was determined using the bicinchoninic acid assay (Pierce Chemical). Based on the calculated protein values for each tissue, 1.85 mg of heart, liver, lung, and kidney cytosolic preparation along with 1.23 mg of spleen were used in the organic extraction process. Tissue-derived cytosolic samples were extracted twice using 3.0 volumes of hexane/ethyl acetate [1:1 (v/v)]. To facilitate separation of the aqueous and organic phases, sample emulsions were centrifuged at 1600g for 10 min at 15°C. After phase separation, the organic component was removed and stripped of residual water through incubation with anhydrous sodium sulfate, and then it was evaporated to completion under a continuous gentle stream of filtered argon gas. Immediately upon reaching dryness, the samples were resolubilized in DMSO, and they were assayed using the HepG2 40/6 reporter cell line as described above.
Real-Time Quantitative Polymerase Chain Reaction. Total RNA was isolated from mice tissues using TRI Reagent (Sigma-Aldrich), and then it was reverse transcribed using the High Capacity cDNA Archive kit (Applied Biosystems, Foster City, CA) according to the respective manufacturer's protocols. The cDNA made from 25 ng of RNA was used for each qPCR reaction. qPCR was performed on DNA Engine Opticon system using the IQ SYBR Green qPCR kit purchased from Bio-Rad Laboratories (Hercules, CA). Data were analyzed and plotted using Prism 4 software (GraphPad Software Inc., San Diego, CA.
Results
CV-1 Cells Contained a Putative Endogenous Ligand That Appeared to Be Metabolized by AhR-Regulated Cytochrome P450 Monooxygenase Enzymes. Previous studies examining AhR nuclear translocation and transcriptional activity revealed that potentially high concentrations of a putative AhR endogenous regulator existed in the CV-1 cell line (Chang and Puga, 1998). The CV-1 cell line is an immortalized animal cell line derived from the kidney epithelium of the African green monkey, a tissue type that in vivo expresses significant levels of AhR. After transient cotransfection of these cells with an AhR expression vector and a DRE-driven luciferase reporter gene construct, high levels of luciferase activity can be observed in the absence of any exogenous ligand. However, based on the fact that AhR activation may not solely be dependent upon ligand, these data do not thoroughly substantiate the existence of an endogenous AhR ligand. Further experimentation was needed to confirm the existence of a high-affinity endogenous AhR ligand in the CV-1 cell line. After confirming the observations of Chang and Puga (1998) in our own laboratory using the CV-1 cell line, we expanded upon this initial discovery through a series of transient transfections to determine the effect of various ectopically expressed cytochrome P450 isoforms on DRE-driven reporter gene activity. The goal was to confirm the presence of a putative endogenous Ah receptor ligand and to test our hypothesis of the existence of a potential AhR-mediated cytochrome P450 autoregulatory pathway.

Transient transfection of cultured CV-1 cells with AhR-regulated cytochrome P450 isoforms leads to a reduction in AhR activity. Expression vectors for CYP1A1, CYP1A2, and CYP1B1 were transiently cotransfected into cultured CV-1 cells, along with an AhR expression vector (pcDNA3/βmAhR) and the DRE-driven luciferase reporter (pGudLuc 6.1). The subsequent effect of P450 expression on DRE-driven luciferase reporter activity was assayed (A). The effect of CYP2E1 on reporter activity served as a negative control (B), whereas transfection of cells with 800 ng of CYP1B1 followed by a 12-h induction with 10 nM TCDD served as a control (C). Each data point represents the mean ± S.D. of four separate determinations. Values are presented as the percentage of change from the negative control.
Expression vectors for cytochrome P450 isoforms CYP1A1, CYP1A2, CYP1B1, and CYP2E1 were individually cotransfected into CV-1 cells, along with an AhR expression vector and a DRE-driven luciferase reporter gene construct. The potential of the CYP1 family members to metabolically alter putative ligand activity was the primary focus of this experiment. CYP2E1 served as a control because it is capable of metabolizing substrates such as ethanol, which are smaller than known AhR ligands (i.e., mol. wt. > 220). Transfection of all cytochrome P450 isoforms was performed in quadruplicate at concentrations between 50 and 400 ng of the appropriate expression vector. Changes in putative endogenous ligand concentration were determined through changes in DRE-driven luciferase reporter activity. Luciferase values were normalized to protein concentration. The results of this experiment (Fig. 1A) suggest the existence of a putative endogenous AhR ligand in CV-1 cells capable of undergoing CYP1 metabolism. In a control experiment, cells transfected with plasmids expressing AhR, CYP1B1 and DRE-driven reporter followed by treatment with TCDD demonstrated that transient expression of CYP1B1 did not lead to inactivation of the AhR pathway (Fig. 1B). The results clearly indicate that TCDD induction leads to a dramatic increase in AhR transcriptional activity, even in the presence of CYP1B1 expression. Expression of CYP2E1 protein in CV-1 cells resulted in a lack of specific metabolic activity toward the putative endogenous AhR ligand (Fig. 1C). Conversely, expression of CYP2E1 resulted in a significant increase in AhR activity with the lower levels of transiently transfected expression vector. The reason for this is not known. Thus, it seems as though specificity exists between this putative AhR ligand and the P450s that are capable of metabolically altering its activity. These results provide evidence suggesting the existence of a possible feedback mechanism in which AhR-regulated cytochrome P450 isoforms from the CYP1A and CYP1B families are able to metabolically alter putative endogenous ligand structure.
Inhibition of CYP1B1 by TMS Rescued AhR-Mediated Transcriptional Activity. To confirm that a direct metabolic alteration of the putative endogenous ligand by CYP1B1 was responsible for the reduction in DRE-driven luciferase reporter gene activity, studies were performed with a cytochrome P450 1B1-specific inhibitor. TMS, a methoxy derivative of 2,4,3′,5′-tetrahydroxystilbene has been shown to be a potent and specific inhibitor of cytochrome P450 1B1 (Chun et al., 2001). CV-1 cells were cotransfected with an AhR expression vector, a DRE-driven luciferase reporter construct and a β-galactosidase-expressing control vector. Transfections proceeded for 12 h before Lipofectamine complexes were removed, and the cells were allowed to recover in complete medium. Eight hours before harvest, cells were treated with TMS in concentrations varying from 50 nM to 10 μM. Treatment with 50 nM TMS induced an approximate 40% recovery in DRE-driven luciferase reporter gene activity relative to cells not cotransfected with pRc/CMV/CYP1B1. Increasing the TMS concentration increased the reporter gene response in a dose-dependent manner up to 750 nM, suggesting that inhibition of CYP1B1 activity leads to an accumulation of an AhR ligand (Fig. 2A). In the absence of ectopic CYP1B1 expression, TMS treatment of CV-1 cells resulted in a dose-dependent induction of DRE-driven luciferase gene activity. This further increase in reporter gene activity seems to be mediated either by the ability of TMS to serve as an AhR agonist or the possibility that TMS blocks endogenous metabolism of the putative ligand. To differentiate between these two possibilities, we tested the ability of TMS to act as an AhR agonist. Bioassay studies were performed using the HepG2 40/6 reporter cell line, and the results of this experiment indicate that TMS does possess weak AhR agonist activity (Fig. 2B). However, an elevation in reporter activity was observed only for TMS concentrations of 750 nM or greater. This result may indicate that, in CV-1 cells at concentrations of TMS below 750 nM, the increase in reporter activity observed may be due to an inhibition of putative AhR ligand metabolism. In addition, the ability of TMS to reverse CYP1B1-mediated inhibition of DRE-driven reporter activity occurs at 50 nM TMS, well below the concentration needed to induce AhR-mediated reporter activity in the HepG2 40/6 cell line. These results taken together indicate that, at low concentrations, TMS is capable of inhibiting CYP1B1 metabolism, thus allowing for a recovery in DRE-driven reporter gene activity.

Inhibition of CYP1B1 activity leads to accumulation of an AhR ligand(s). CV-1 cells were cotransfected with an AhR expression vector, a DRE-driven luciferase reporter construct and a β-galactosidase expressing control vector both in the presence and absence of pCMV/hCYP1B1 vector for 12 h. After transfection and recovery, the cells were treated with the CYP1B1-specific inhibitor TMS (A). The ability of TMS to act as an AhR agonist was also examined via bioassay in the human-derived HepG2 40/6 reporter cell line (B). Each data point represents the mean ± S.D. of four separate determinations. Values are presented as relative luciferase units, and they have been normalized to protein concentration.
Ligand Characterization. To identify the putative endogenous AhR ligand(s) from CV-1 cells, a purification strategy needed to be developed, in the absence of any information on the chemical characteristics of the molecule. This necessitated making several initial assumptions, many of which were ultimately verified through experimental testing. We rationalized that an endogenous ligand for the AhR, like most xenobiotic ligands for this receptor, would be relatively hydrophobic in nature; thus, it would be able to partition into hydrophobic stationary phase or organic solvent system. It was further hypothesized that to activate the AhR in vivo, the endogenous ligand must be capable of partitioning into the cytoplasm. Thus, the cytosolic fraction seemed to be the optimal source of starting material in the search for endogenous AhR ligands. Furthermore, to choose a fraction that contained membranes would lead to organic extracts laden with high concentrations of neutral lipids, known to interfere with our assay system.
First, solid phase extraction methods were used to concentrate and purify the active metabolite. After testing several hydrophobic sorbent materials, it was determined that a C18 reverse phase resin was the most efficient, confirming our assumption that the active molecule was relatively hydrophobic. All preliminary purification attempts focused on the use of C18 solid phase extraction columns to concentrate the active compounds before high-performance liquid chromatography (HPLC) analysis, but this method was later replaced by liquid extraction using high-purity organic solvents after discovering the C18 sorbent material itself possessed significant AhR activity (data not shown). Various solvents and solvent combinations were tested to determine an efficient solvent system for the liquid-liquid extraction method. Preliminary data demonstrated that several solvents, including hexane, ethyl acetate, dichloromethane, and a mixture of hexane/ethyl acetate [1:1 (v/v)] were effective in extracting the endogenous AhR ligand activity from CV-1 cytosol (data not shown). It was also discovered that using excess anhydrous sodium sulfate to remove any residual trace of water in the organic extract greatly aids in preserving the integrity of the active compound (Fig. 3A). Furthermore, it was determined that solvent extraction efficiency of cytosol is enhanced through acidification of cytosolic samples before organic extraction (data not shown). This result presumably indicates the presence of one or more ionizable functional groups present in the structure of the putative endogenous ligand. Thus, throughout the experiments presented in this article, a hexane/ethyl acetate [1:1 (v/v)] solvent system was used to extract acidified cytosolic fractions derived from either cultured cells or tissue samples. Although the initial information on the chemical properties of the putative, endogenous AhR ligand contained in CV-1 cells is strictly qualitative, it has proven extremely valuable, especially in performing large-scale, high-yield, organic extractions of cytosolic fractions.

Organic solvent extractions of CV-1 cytosol can activate the AhR. Both hexane and ethyl acetate extracts of CV-1 cytosol were tested for DRE-driven luciferase activity in the HepG2 40/6 reporter cell line (A). Values are presented as relative luciferase units, and they have been normalized to protein concentration. A dried sample is defined as having any aqueous residue removed through the use of anhydrous sodium sulfate. DNA mobility shift assays were performed with the organic extracts. A mixture of in vitro-translated AhR and ARNT was mixed with or without the addition of a ligand, and then it was subjected to electrophoresis on a nondenaturing polyacrylamide gel (B). The control (–) represents a control translation and an AhR translation mixed together and treated with TCDD (minus ARNT).
Electrophoretic Mobility Shift Assays Confirmed the Presence of an Endogenous AhR Ligand Present in CV-1 Cell Extracts. DNA mobility shift assays were performed to demonstrate that the ability of these CV-1 extracts to induce reporter activity in HepG2 40/6 cells was indeed the direct result of AhR activation by a true high-affinity endogenous ligand. Liquid phase organic extracts of CV-1 cytosol were prepared by extracting the required volume of cytosol with 3 volumes of hexane/ethyl acetate [1:1 (v/v)]. The organic phase was removed to a clean vessel and stripped of any residual water through a brief incubation with anhydrous sodium sulfate. The extract was then aliquoted, dried under reduced atmospheric pressure, and solubilized in DMSO. Vectors for the AhR and ARNT proteins were in vitro-translated in rabbit reticulocyte lysate. Translated AhR and ARNT proteins were mixed together in a 2:1 M ratio. A solvent blank made up of evaporated hexane/ethyl acetate served as a background control. The mobility of the 32P-DRE-AhR:ARNT complex from in vitro translation mixture-transformed AhR was compared with that of TCDD-induced AhR/ARNT. The results of Fig. 3B demonstrate that an organic extract of CV-1 cytosol contains a true endogenous AhR ligand capable of activating the AhR signaling pathway apparently through direct binding to the AhR.
Proteinase K Treatment Did Not Alter DRE-Driven Luciferase Reporter Activity. To differentiate between a low-molecular-weight endogenous ligand and the possibility of a bioactive peptide modulator, a protease treatment experiment was performed. CV-1 cytosol was incubated at room temperature in the presence or absence of protease. The protease treatment resulted in digestion of protein into amino acids and small peptides (Fig. 4A) Protease treatment failed to abolish or significantly alter DRE-driven luciferase activity in organic solvent extracts from protease-digested cytosol samples (Fig. 4B). This suggests that the large -fold induction in reporter gene activity seen with solvent-extracted CV-1 cytosol is presumably not due to a peptide ligand or a modulatory protein molecule.
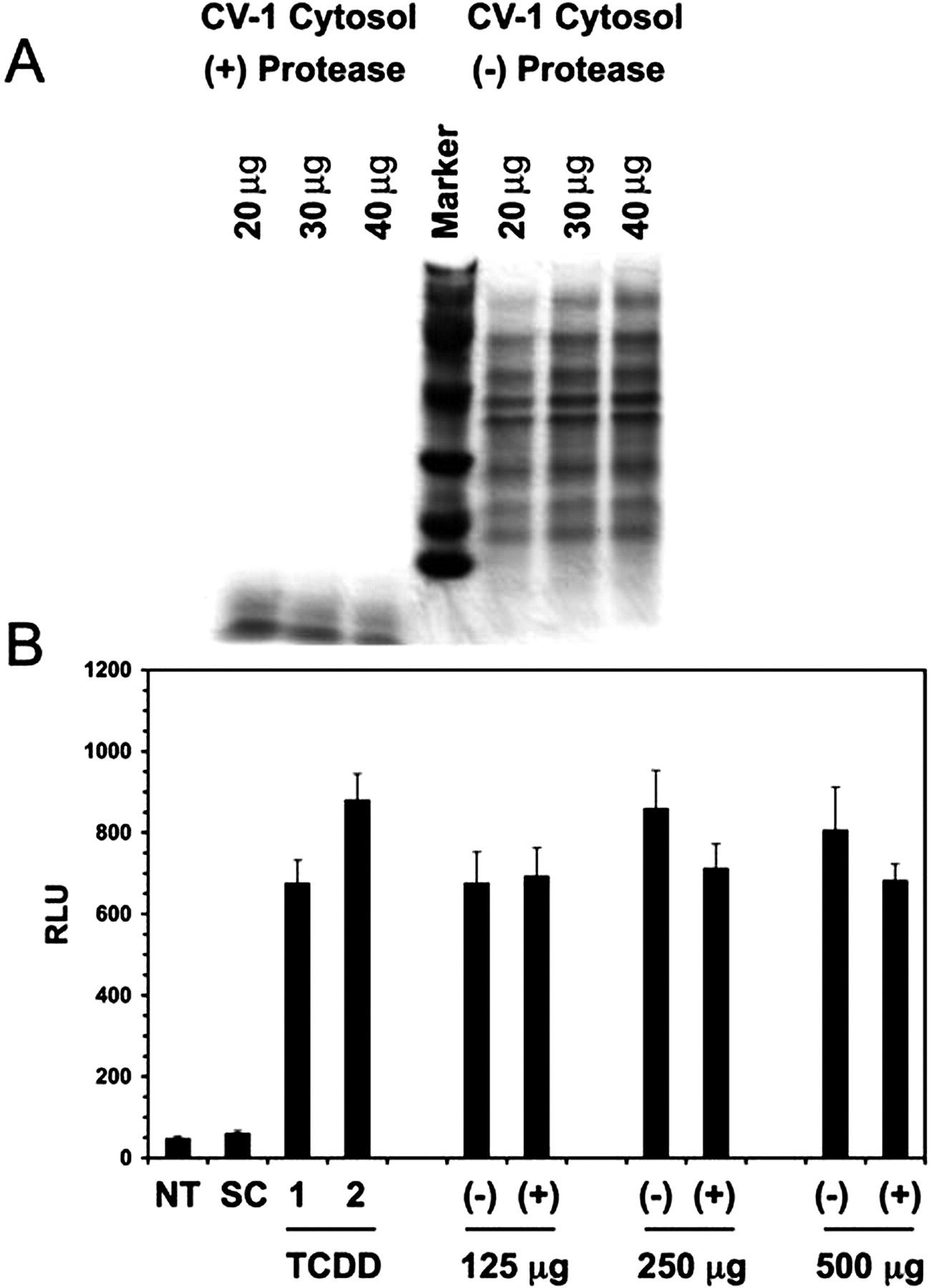
Treatment of CV-1 cytosol with proteinase K does not alter DRE-driven luciferase activity. Cytosolic protein was treated with protease for 90 min at room temperature. Aliquots from both protease-treated and untreated samples were resolved by Tricine SDS-polyacrylamide gel electrophoresis (A). Organic extractions of both cytosolic samples were tested for biological activity in the HepG2 40/6 reporter cell line (B). Each data point represents the mean ± S.D. of three separate determinations. Values are presented as relative luciferase units, and they have been normalized to protein concentration.
HPLC Fractionation of CV-1 Cytosol Indicated a Complex Mixture of AhR-Modulating Compounds. First, the fractionation of cytosolic samples was performed using reverse phase HPLC. However, this approach was eventually replaced by a classic normal phase HPLC methodology to alleviate stability problems associated with exposing the unstable active samples to solvent systems containing water. The separation revealed the presence of multiple peaks with AhR activation potential and a wide range of retention times (Fig. 5). These results confirm the complexity of activity in the organic extract, presumably because of the presence of multiple AhR ligands. On the other hand, this activity profile may also be indicative of unresolved stability issues involving a single ligand that is degrading into several active products. Nevertheless, the activity is capable of being separated into distinct peaks, allowing a high level of purity to be potentially obtained.
Endogenous Ligands for the AhR Accumulated in the Lung of Ahr-Null Mice. Based on the results obtained in CV-1 cells, we hypothesized that in the absence of AhR expression, certain AhR ligands would accumulate in tissues because they would be unable to induce their own cytochrome P450-mediated metabolism. To test this hypothesis cytosolic extracts from various C57BL6/J and Ahr-null mouse tissues were extracted with organic solvent. The level of AhR transcriptional activity induced by these extracts was assessed using the HepG2 40/6 reporter cell line. Of the Ahr-null tissues tested, only lung seems to significantly accumulate AhR ligands, where a 4-fold increase in AhR transcriptional activity was observed (Fig. 6A). A decrease in AhR activity was actually seen in the heart of Ahr-null mice, which was an unexpected result. However, the dramatic increase in the presence of AhR ligands in lung supports our hypothesis that the absence of AhR would lead to the accumulation of ligands in Ahr-null mice that are normally metabolized by enzymes regulated by the AhR.
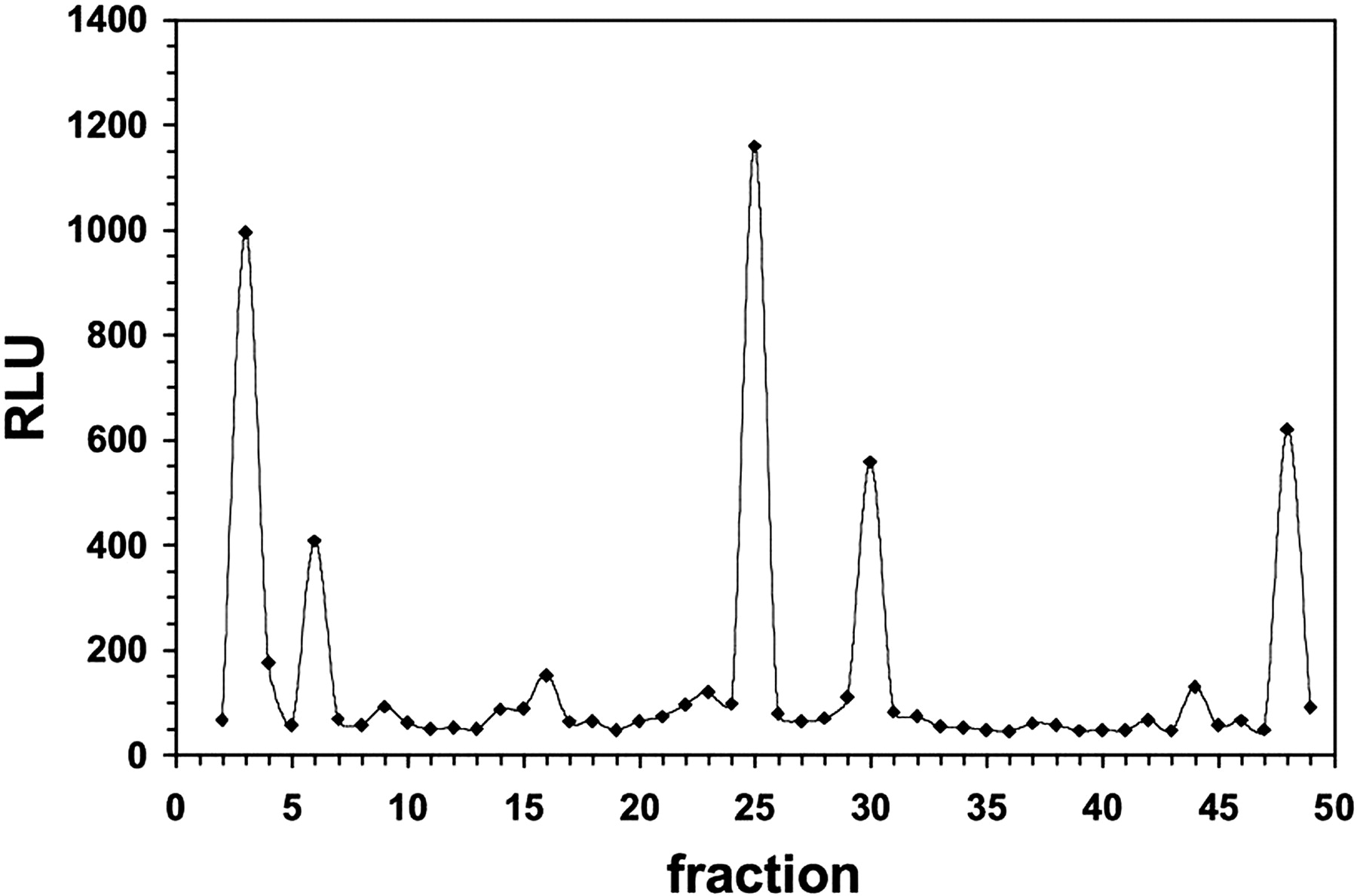
HPLC fractionation of putative endogenous AhR ligands from CV-1 cells. Fractionation of organic extracts derived from CV-1 cytosol was achieved using a Phenomenex Luna semipreparative (10 × 250 mm) silica column (10 μm) with a mobile phase solvent system made up of hexane/acetic acid [99.9:0.1 (v/v)]. Multiple fractions were collected and analyzed for biological activity in the HepG2 40/6 reporter cell line. DRE-driven luciferase reporter activity is plotted as a function of time.
Mouse Lung Exhibited Relatively High Constitutive CYP1A1 Expression. The constitutive level of CYP1A1 and CYP1B1 mRNA was determined in various mouse tissues isolated from C57BL6/J and Ahr-null mice. It is noteworthy that lung exhibited 4-fold more CYP1A1 mRNA than the heart, the tissue with the second highest level of CYP1A1 mRNA in C57BL6/J mice (Fig. 6B). In contrast, the CYP1B1 levels were relatively low in lung. The level of constitutive CYP1A1 mRNA expression was almost totally dependent on AhR expression. It was surprising that CYP1B1 mRNA expression was essentially the same in C57BL6/J and Ahr-null mice, indicating that constitutive expression of CYP1B1 is not dependent on the AhR. The CYP1A1 results would suggest that the AhR is constitutively active in mouse lung.

Examining the level of constitutive Ah receptor transcriptional activity and AhR ligands in various tissues from AhR+/+ and AhR–/– mice. Hexane/ethyl acetate [50:50 (v/v)] extraction of cytosol prepared from various mouse tissues produces organic extracts capable of driving the expression of a DRE-controlled luciferase reporter gene. Extracts derived from the tissues of both C57BL6/J and Ahr-null mouse strains are examined for differences in their ability to modulate AhR activity (A). The constitutive expression of CYP1A1 and CYP1B1 mRNA in Ahr-null and wild-type mouse tissues is examined (B and C). RNA obtained from different mouse tissues was reverse transcribed and analyzed for Cyp1A1 and Cyp1B1 mRNA abundance using real-time reverse transcription-polymerase chain reaction. The graphs represent data obtained from two animals of each genotype (wild-type and Ahr-null mice). The experiment was repeated twice, independently, and both replicates revealed the same results. Each bar represents the mean of three separate determinations ± S.D. Closed bars represent wild-type mice, whereas open bars represent Ahr-null mice.
Discussion
This study is one of the first attempts to isolate putative endogenous ligands from biological extracts under gentle extraction conditions. The results provide strong evidence supporting the existence of a putative high-affinity endogenous ligand(s) for the AhR using CV-1 cells as a model, and they are consistent with previously published observations (Chang and Puga, 1998). These ligands appear to be capable of modulating receptor activation through direct binding, and they are able to induce DRE-driven luciferase reporter gene expression to a level greater than or equal to a saturating dose of TCDD. Furthermore, it seems as though the putative ligand(s) is capable of relatively high-affinity interaction with the AhR, allowing the receptor to mediate the metabolism of its own putative endogenous ligand(s). This would suggest the existence of a negative feedback autoregulatory pathway in which ligand activation of the AhR would lead to activation of genes in the Ah battery, including cytochrome P450 1A1, 1A2, and 1B1 (Fig. 7). Expression of these P450 enzymes could then mediate metabolism of the putative endogenous ligands, resulting in the attenuation of the AhR pathway. In addition, considering the modest amount of transfected plasmid needed in Fig. 1 to almost completely suppress Ah receptor constitutive activity, it would seem that even a modest increase in P450 expression may be capable of completing this feedback loop, especially CYP1A1, which seems to exhibit the highest efficiency of ligand metabolism. An autoregulatory feedback mechanism can occur upon induction of oxidant stress leading to repression of CYP1A1 gene expression, through down-regulation of NF1 activity (Morel et al., 1999). Oxidant stress can occur through cytochrome P4501A1-mediated metabolism of AhR ligand such as benzo[a]pyrene. The ability of CYP1A1 to autoregulate its own expression in response to exogenous AhR ligands, such as polycyclic aromatic hydrocarbons, has been well documented (RayChaudhuri et al., 1990; Schaldach et al., 1999), so it is reasonable to expect that this type of regulation could occur in the context of cellular homeostasis.
Derived from the kidney epithelium of the African green monkey, the CV-1 cell line is an immortalized cell line exhibiting minimal AhR expression. Therefore, the levels of AhR-regulated cytochromes P450 would be extremely low, leading to a subsequent accumulation of cellular metabolites, including potential intracellular endogenous AhR ligands. Thus, an increased level of a relatively high-affinity endogenous ligand may explain the elevated levels of DRE-driven luciferase reporter gene activity observed in these cells when transfected with an AhR expression vector and a DRE-driven luciferase reporter gene construct. This may also explain the ability of purified cytosolic extracts to induce luciferase reporter gene expression to a level equal to or greater than that observed for a saturating dose of TCDD (data not shown). Similar observations have also been reported in bioassay experiments using purified prostaglandins, especially prostaglandin G2 (Seidel et al., 2001). It is also possible that, in addition to being a potent high-affinity AhR agonist, the putative endogenous ligand in CV-1 cells may also be capable of stimulating the AhR pathway by an indirect mechanism.
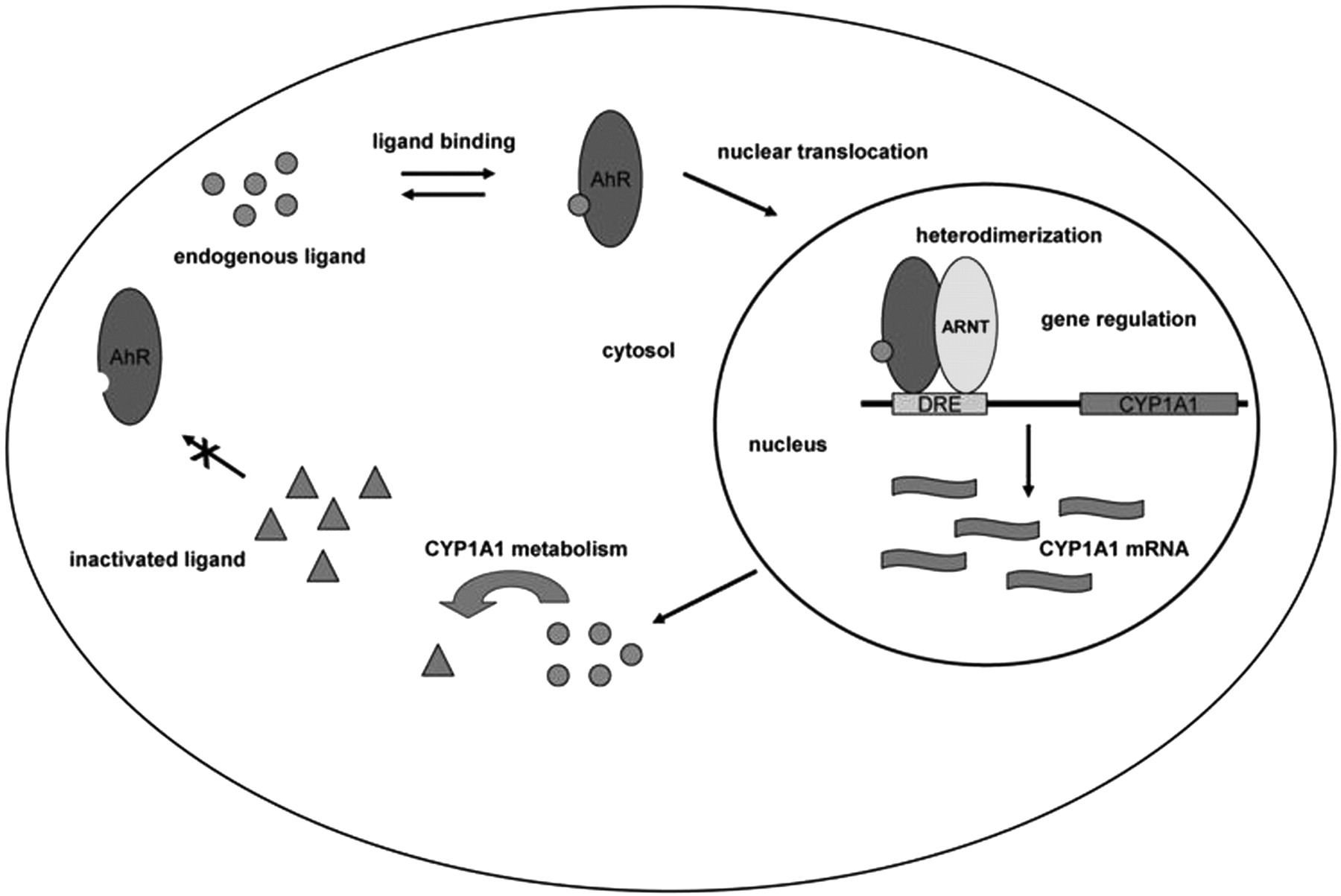
An overall scheme depicting the proposed negative feedback loop mechanism in mouse lung.
The possibility of a cell culture media contaminant producing the observed induction in DRE-driven luciferase reporter activity upon treatment with organic solvent extract of CV-1 cell cytosol was addressed in several ways. All bioassay experiments performed have included as one of several controls a media blank in which HepG2 40/6 cells receive no treatment other than the cell culture media in which they are routinely grown. No induction in reporter gene activity has ever been observed with media incubation alone (data not shown). Second, a time course assay was performed using the HepG2 40/6 reporter cell line. Media were incubated in a cell culture incubator for 48 h before the start of the experiment. Every 6 h for an 18-h period, media were removed and added to HepG2 40/6 cells. An induction in reporter gene activity was never observed with conditioned media. In a similar experiment to test the possibility of a secreted metabolite, media conditioned for 48 h by actively growing CV-1 cells was added to the HepG2 40/6 reporter cell line every 6 h for an 18-h period. Once again, no induction was observed in DRE-driven luciferase reporter gene activity.
Our results have revealed the presence of a relatively potent, putative endogenous ligand in CV-1 cells capable of activating the AhR pathway by directly binding and transforming the receptor to its DNA-binding form. In addition, this putative endogenous ligand can be metabolized by AhR-regulated cytochrome P450s. This nonpeptide ligand is easily extracted into aprotic solvents after acidification, suggesting that the compound is hydrophobic in nature with one or more ionizable acidic groups. In addition, the presence of multiple peaks of activity upon HPLC fractionation of extracts from CV-1 cell cytosol would suggest that more than one AhR ligand exists in CV-1 cells. Attempts to further purify these individual peaks have been complicated by their apparent instability (data not shown).
To further explore the significance of the data obtained in CV-1 cells, we wanted to test whether an AhR-cytochrome P450 autoregulatory loop exists in vivo. To accomplish this, cytosolic samples of five tissues from “wild-type” and Ahr-null mice were extracted with organic solvent and tested for their ability to activate the AhR in a reporter cell line. It is noteworthy that lung extracts from Ahr-null mice revealed a 4-fold increase in AhR activation potential compared with wild-type mice. This observation, coupled with the relatively high AhR-dependent CYP1A1 constitutive activity seen in lung, 50-fold higher than any other tissue tested, would support the assertion that an autoregulatory loop between the AhR and CYP1A1 exists. It is interesting to note that the constitutive level of CYP1B1 in lung seems to be quite low. These data would indicate that lung from Ahr-null mice would be a rich source of putative endogenous AhR ligands. Whether this activity is due to the previously identified compound 2-(1′H-indole-3′-carbonyl)-thiazole-4-carboxylic acid methyl ester from porcine lung will require further investigation (Song et al., 2002). Considering that the lung is a complex tissue made up of a number of cell types, future studies will focus on the specific cell type(s) in Ahr-null mice that accumulate AhR ligands. Although no quantitative differences in the presence of AhR ligands was seen in other tissues, it is possible that the method chosen to assess the presence of ligands may not be examining all AhR ligands, such as more polar compounds. The ability of AhR ligands to accumulate in tissues of Ahr-null mice may be attenuated by the presence of constitutively expressed CYP1B1, CYP1A2, or other P450s that are not totally dependent of AhR to mediate expression. These observations also suggest that exogenous AhR ligands in the diet are not accumulating in Ahr-null mice. If this were the case, we would expect to detect increases in AhR ligands in Ahr-null mouse tissues such as liver. In summary, the in vivo data clearly support the presence of an AhR-mediated cytochrome P450 autoregulatory loop in mouse lung.
Efforts to identify the compound using various complementary methods of mass spectrometry and tandem mass spectrometry are underway in both CV-1 cells and in Ahr-null mouse lung. Results of these experiments should reveal the identity of this putative AhR ligand(s) and provide insight into the types of compounds that can serve as high-affinity endogenous ligands for the AhR. This information will greatly improve our understanding of the physiological role of the AhR and the subsequent biochemical mechanism of action of this receptor.
Acknowledgments
We thank Drs. Steve Safe, William Greenlee, Mike Denison, Judy Raucy, and Robert Tukey for TCDD, pRcCMV-hCYP1B1, pGudLuc 6.1, pCR2.1-hCYP2E1, pCMV4-hCYP1A1, and pCMV4-hCYP1A2, respectively.
Footnotes
-
This work was supported by the National Institute of Environmental Health Science grant ES09878 (to C.B.M.) and ES04869 and ES11834 (to G.H.P.).
-
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
-
doi:10.1124/mol.107.038968.
-
ABBREVIATIONS: AhR, aryl hydrocarbon receptor; Ah, aryl hydrocarbon; ARNT, aryl hydrocarbon receptor nuclear translocator; DRE, dioxin responsive element(s); TCDD, 2,3,7,8-tetrachlorodibenzo-p-dioxin; DMSO, dimethyl sulfoxide; PBS, phosphate-buffered saline; MEM, minimal essential media; CHAPS, 3-[(3-cholamidopropyl)dimethylammonio]propanesulfonate; qPCR, quantitative polymerase chain reaction; HPLC, high-performance liquid chromatography.
- Received June 13, 2007.
- Accepted August 24, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}