


Abstract
Resveratrol undergoes extensive metabolism to form biologically active glucuronides in humans. However, the transport mechanisms for resveratrol glucuronides are not fully established. Here, we aimed to characterize the efflux transport of resveratrol glucuronides using UGT1A1-overexpressing HeLa cells (HeLa1A1 cells), and to determine the contribution of multidrug resistance–associated protein (MRP) 4 to cellular excretion of the glucuronides. Two glucuronide isomers [i.e., resveratrol 3-O-glucuronide (R3G) and resveratrol 4′-O-glucuronide (R4′G)] were excreted into the extracellular compartment after incubation of resveratrol (1–100 μM) with HeLa1A1 cells. The excretion rate was linearly related to the level of intracellular glucuronide, indicating that glucuronide efflux was a nonsaturable process. MK-571 (a dual inhibitor of UGT1A1 and MRPs) significantly decreased the excretion rates of R3G and R4′G while increasing their intracellular levels. Likewise, short-hairpin RNA (shRNA)–mediated silencing of MRP4 caused a significant reduction in glucuronide excretion but an elevation in glucuronide accumulation. Furthermore, β-glucuronidase expressed in the cells catalyzed the hydrolysis of the glucuronides back to the parent compound. A cellular pharmacokinetic model integrating resveratrol transport/metabolism with glucuronide hydrolysis/excretion was well fitted to the experimental data, allowing derivation of the efflux rate constant values in the absence or presence of shRNA targeting MRP4. It was found that a large percentage of glucuronide excretion (43%–46%) was attributed to MRP4. In conclusion, MRP4 participated in cellular excretion of R3G and R4′G. Integration of mechanistic pharmacokinetic modeling with transporter knockdown was a useful method to derive the contribution percentage of an exporter to overall glucuronide excretion.
Introduction
Resveratrol is a natural polyphenol widely distributed in plant kingdom (e.g., in grapes, peanuts, and berries) (Shishodia and Aggarwal, 2006). It possesses numerous health-promoting effects including antioxidant, anti-inflammatory, chemopreventive, and cardioprotective properties (Bishayee, 2009; Brisdelli et al., 2009; Jasiński et al., 2013; Poulsen et al., 2013; Szkudelski and Szkudelska, 2015). However, bioavailability of resveratrol is exceedingly low (<1%) after oral uptake (Walle et al., 2004; Wenzel and Somoza, 2005; Walle, 2011). Extensive glucuronidation is found to be the main cause of low oral bioavailability (Walle et al., 2004; Walle, 2011). Glucuronidation results in high levels of circulating glucuronides [resveratrol 3-O-glucuronide (R3G) and resveratrol 4′-O-glucuronide (R4′G)] that are suggested to be one source of the beneficial effects of resveratrol (Walle et al., 2004; Walle, 2011).
Efflux transporters such as breast cancer resistance protein, multidrug resistance–associated protein (MRP) 2, and MRP3 have been identified as contributors to intestinal/hepatic excretion of resveratrol glucuronides (Adachi et al., 2005; Maier-Salamon et al., 2006; van de Wetering et al., 2009). However, it remains unknown whether the MRP4 transporter (with significant expression in the liver and kidney) is involved in transport of resveratrol glucuronides (Russel et al., 2008). The objective of the present study was to determine the role of MRP4 in excretion of resveratrol glucuronides using UGT1A1 stably transfected HeLa cells. Contribution of MRP4 to glucuronide excretion was quantified by coupling short-hairpin RNA–mediated silencing with mechanistic pharmacokinetic modeling.
Materials and Methods
Materials.
Expressed human UGT1A1 was purchased from BD Biosciences (Woburn, MA). Anti-β-glucuronidase (GUSB) and anti-β-actin antibodies were purchased from Abcam (Cambridge, MA). MK-571 was obtained from Sigma-Aldrich (St. Louis, MO). Resveratrol was purchased from Aladdin Reagents (Shanghai, China). R3G and R4′G were purchased from Cayman Chemical (Ann Arbor, MI).
Establishment of UGT1A1- and GUSB-overexpressing HeLa Cells.
UGT1A1- or GUSB-overexpressing HeLa cells were generated using the lentiviral transfection method as previously described (Quan et al., 2015; Sun et al., 2015). Western blotting against GUSB was performed as previously described (Quan et al., 2015).
Glucuronide Excretion Experiments.
Glucuronide excretion experiments were performed following the published procedures (Quan et al., 2015). Resveratrol and its glucuronides (R3G and R4′G) were quantified using the Waters ACQUITY ultra-performance liquid chromatography system (Milford, MA).
Statistical Analysis.
Data are expressed as mean ± S.D. Statistically significant differences were analyzed by unpaired Student’s t test. The level of significance was set at P < 0.05 (*), P < 0.01 (**), or P < 0.001 (***).
Results and Discussion
Effects of MK-571 on Disposition of Resveratrol Glucuronides.
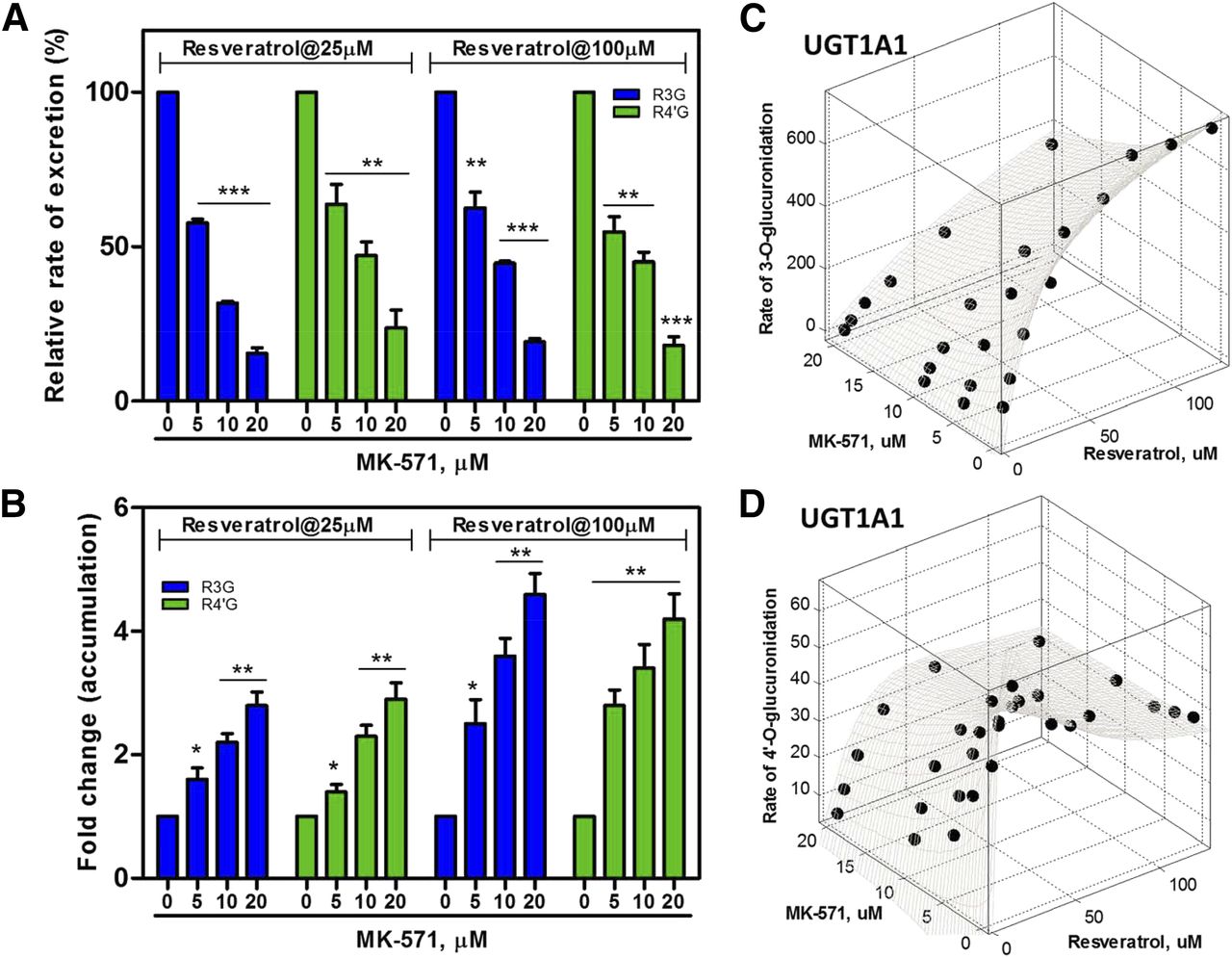
MK-571 (a pan-MRP inhibitor) caused a significant reduction (>36%, P < 0.01) in excretion of resveratrol glucuronides (Fig. 1A). A maximal reduction was associated with the highest MK-571 concentration of 20 μM (Fig. 1A). By contrast, MK-571 led to a significant elevation (>140%, P < 0.05) in glucuronide accumulation (Fig. 1B). The extent of elevation was positively correlated with the MK-571 concentration (Fig. 1B). The results suggested that MRP family proteins may be responsible for excretion of resveratrol glucuronides.

Effects of MK-571 on disposition of resveratrol glucuronides after incubation of HeLa1A1 cells with resveratrol (25 and 100 μM). (A) Effects of MK-571 on the excretion rates of resveratrol glucuronides. (B) Effects of MK-571 on the intracellular levels of resveratrol glucuronides. Each data point was the average of three determinations with the error bar representing the standard deviation (*P < 0.05; **P < 0.01; ***P < 0.001 compared with the control group). (C) Kinetic modeling of the effects of MK-571 on R3G formation by UGT1A1. (D) Kinetic modeling of the effects of MK-571 on R4′G formation by UGT1A1.
Effects of MK-571 on Resveratrol Glucuronidation by UGT1A1.
The effects of MK-571 on glucuronidation of resveratrol were determined using expressed UGT1A1 enzyme. Formation of both R3G and R4′G was significantly inhibited by MK-571 (Fig. 1, C and D). The inhibition kinetics for R3G formation was best described by the competitive inhibition model (Fig. 1C; Supplemental eq. 4; Supplemental Table 1). The inhibition kinetics for R4′G formation was well fitted by a two-site inhibition model (Fig. 1D; Supplemental eq. 10; Supplemental Table 1). The models consistently suggested that MK-571 inhibited glucuronidation of resveratrol through its binding to the reaction site of UGT1A1. Since the enzyme activity was inhibited by MK-571, reduced glucuronide excretion may not be ascribed to suppression of MRP transporters by MK-571 (Fig. 1A). However, the significant increase in glucuronide accumulation caused by MK-571 was strong evidence that one or more MRP family proteins contributed to cellular excretion of resveratrol glucuronides (Fig. 1B). This was because elevated glucuronide accumulation must result from blocked glucuronide efflux though the action of MK-571 on MRPs.
Effects of MRP4 Knockdown on Glucuronide Disposition.
MRP4 was knocked down by short-hairpin RNA–mediated gene silencing. The selected short-hairpin RNA was able to decrease the expression of MRP4 by 65% in HeLa cells (Quan et al., 2015). Knockdown of MRP4 caused a significant reduction (P < 0.05) in excretion of resveratrol glucuronides (Supplemental Fig. 1A). On the contrary, MRP4 knockdown led to a significant elevation (P < 0.05) in intracellular glucuronide (Supplemental Fig. 1B). The alterations in glucuronide disposition indicated that the MRP4 transporter was involved in excretion of resveratrol glucuronides. Transport of the resveratrol glucuronides by MRP4 was further confirmed by the vesicular transport assays (Supplemental Fig. 2). It was a novel finding that MRP4 was involved in the transport of resveratrol glucuronides. Previous studies indicated that breast cancer resistance protein, MRP2, and MRP3 were responsible for excretion of resveratrol glucuronides (Adachi et al., 2005; Maier-Salamon et al., 2006; van de Wetering et al., 2009). Breast cancer resistance protein and MRP2 (two apical exporters) facilitated elimination of the glucuronides via biliary and intestinal routes. By contrast, MRP3 and MRP4 (two hepatic basolateral exporters) contributed to systemic exposure of the glucuronides via basolateral excretion.
GUSB-Mediated Hydrolysis of Resveratrol Glucuronides.
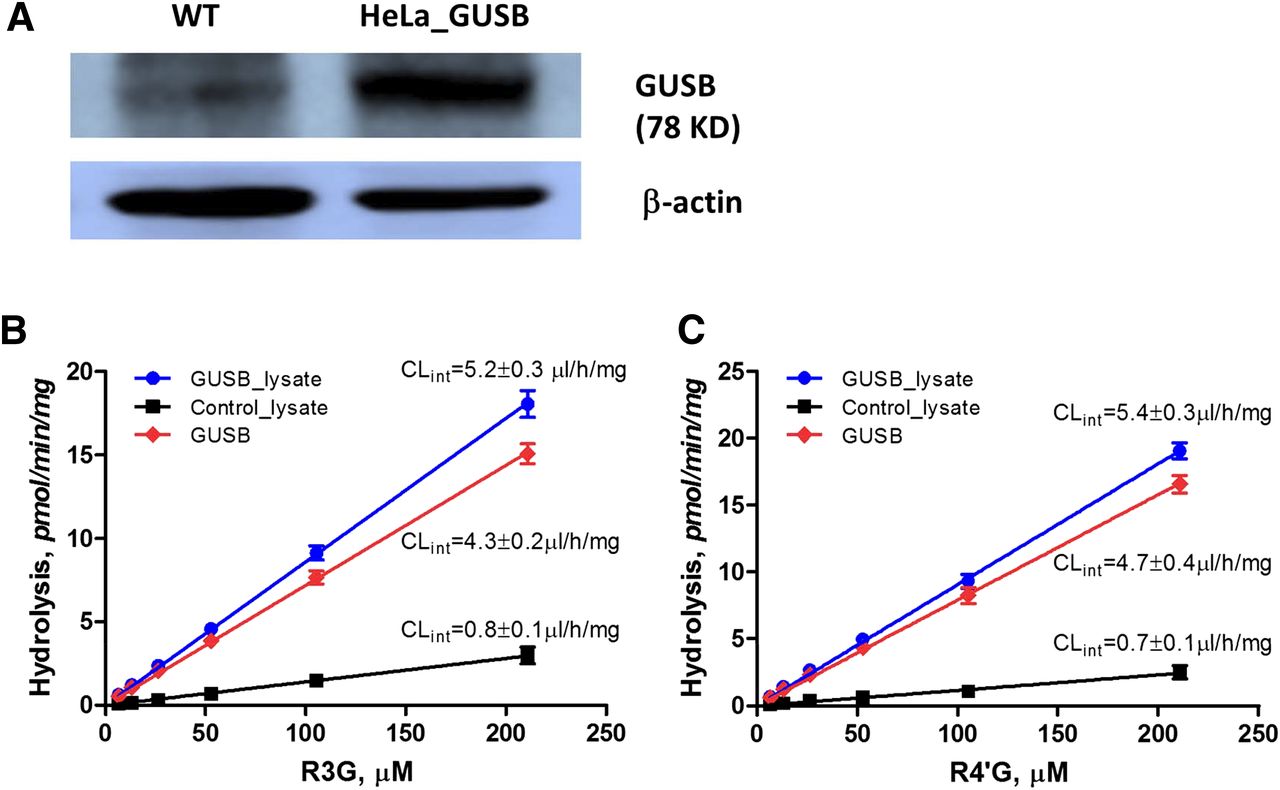
The hydrolysis potential of resveratrol glucuronides (R3G and R4′G) in HeLa cells was determined using the lysate preparations from HeLa (control lysate) and GUSB-overexpressing HeLa cells (GUSB lysate). Overexpression of GUSB was confirmed by western blotting (Fig. 2A). Both the control lysate and GUSB lysate were able to catalyze hydrolysis of resveratrol glucuronides (R3G and R4′G) (Fig. 2, B and C). As expected, the GUSB lysate showed a much higher hydrolysis activity toward the glucuronides compared with the control lysate (Fig. 2, B and C). The intrinsic clearance for the hydrolysis reaction was estimated by performing linear regression (the intrinsic clearance equaled the slope). The GUSB enzyme possessed similar activities toward the two glucuronide isomers (4.3 versus 4.7 μl/h/mg, P > 0.05) (Fig. 2, B and C).

Functional characterization of GUSB in hydrolysis of resveratrol glucuronides. (A) Protein expression of GUSB in wild-type HeLa (WT) and GUSB-overexpressing HeLa cells (HeLa_GUSB). (B) Hydrolysis kinetics of R3G by HeLa lysate (control) and HeLa_GUSB lysate. (C) Hydrolysis kinetics of R4′G by HeLa lysate (control) and HeLa_GUSB lysate. The hydrolysis activity of GUSB enzyme was derived by subtracting the hydrolysis rate of HeLa cell lysate from that of HeLa_GUSB lysate. Each data point was the average of three determinations with the error bar representing the standard deviation and Clint representing the intrinsic clearance.
Mechanistic Pharmacokinetic Modeling of Resveratrol Disposition in Control versus MRP4 Knockdown Cells.
The concentration-time profiles were determined for resveratrol and its glucuronides in both extracellular and intracellular compartments after administration of 50 nmol resveratrol. A cellular pharmacokinetic model was used to describe the data (Supplemental Figs. 3 and 4). Mechanistic fitting was performed by fixing the binding affinity parameters [the Michaelis-Menten constant (Km) and substrate inhibition constant (Ksi), independent of the model system] to their corresponding values derived from an in vitro glucuronidation assay (Supplemental Tables 2 and 3). The coefficients of variation for all fitted parameters were less than 20%, suggestive of adequate fitting of the model to the data (Supplemental Table 2). The estimated values for all parameters except the efflux rate constants (Kef,1 and Kef,2) were similar (P > 0.05) between the control and MRP4 knockdown cells. The Kef,1 and Kef,2 values were reduced to 72% ± 4.2% and 70% ± 3.6% of the control, respectively. Based on the relative expression level of MRP4, it was estimated that MRP4 contributed 43% ± 1.6% of R3G excretion and 46% ± 2.4% of R4′G excretion in HeLa1A1 cells.
The established pharmacokinetic model (Supplemental Fig. 3) was scientifically solid. First, the model assumed that resveratrol passively diffused across cell membranes. Passive diffusion as the main transport mechanism for resveratrol has been documented in the literature (Henry et al., 2005). Second, the model included a deglucuronidation process. This was necessary because GUSB enzyme expressed in the cells was able to hydrolyze resveratrol glucuronides back to the parent molecule. Third, the model assumed that glucuronide efflux was a nonsaturable process. This assumption appeared to be valid because the excretion rate of glucuronide was linearly related to the intracellular glucuronide (Supplemental Fig. 5). Fourth, it was assumed that intracellular binding of glucuronides was negligible. This was well supported by the fact the hydrophilic glucuronides hardly bound to lysate proteins (Supplemental Fig. 6).
In summary, this study has demonstrated for the first time that MRP4 participates in cellular excretion of resveratrol glucuronides and that β-glucuronidase is a key determinant to total cellular glucuronidation. Furthermore, integration of mechanistic pharmacokinetic modeling with transporter knockdown is a useful method to derive the contribution percentage of an exporter to overall glucuronide excretion in a cell system expressing multiple transporters.
Authorship Contributions
Participated in research design: Wang, Li, Dong, Wu.
Conducted experiments: Wang, Li, Quan, Dong.
Contributed new reagents or analytic tools: Dong.
Performed data analysis: Wang, Li, Wu.
Wrote or contributed to the writing of the manuscript: Wang, Li, Wu.
Footnotes
- Received October 7, 2015.
- Accepted January 8, 2016.
↵1 S.W. and F.L. contributed equally to this work.
This work was supported by the National Natural Science Foundation of China [Grant 81573488]; the Young Scientist Special Projects in Biotechnological and Pharmaceutical Field of 863 Program [Grant 2015AA020916]; and the Outstanding Youth Fund from the Natural Science Foundation of Guangdong Province [Grant 2014A030306014].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- GUSB
- β-glucuronidase
- MRP
- multidrug resistance–associated protein
- R3G
- resveratrol 3-O-glucuronide
- R4′G
- resveratrol 4′-O-glucuronide
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}