


Visual Overview

Abstract
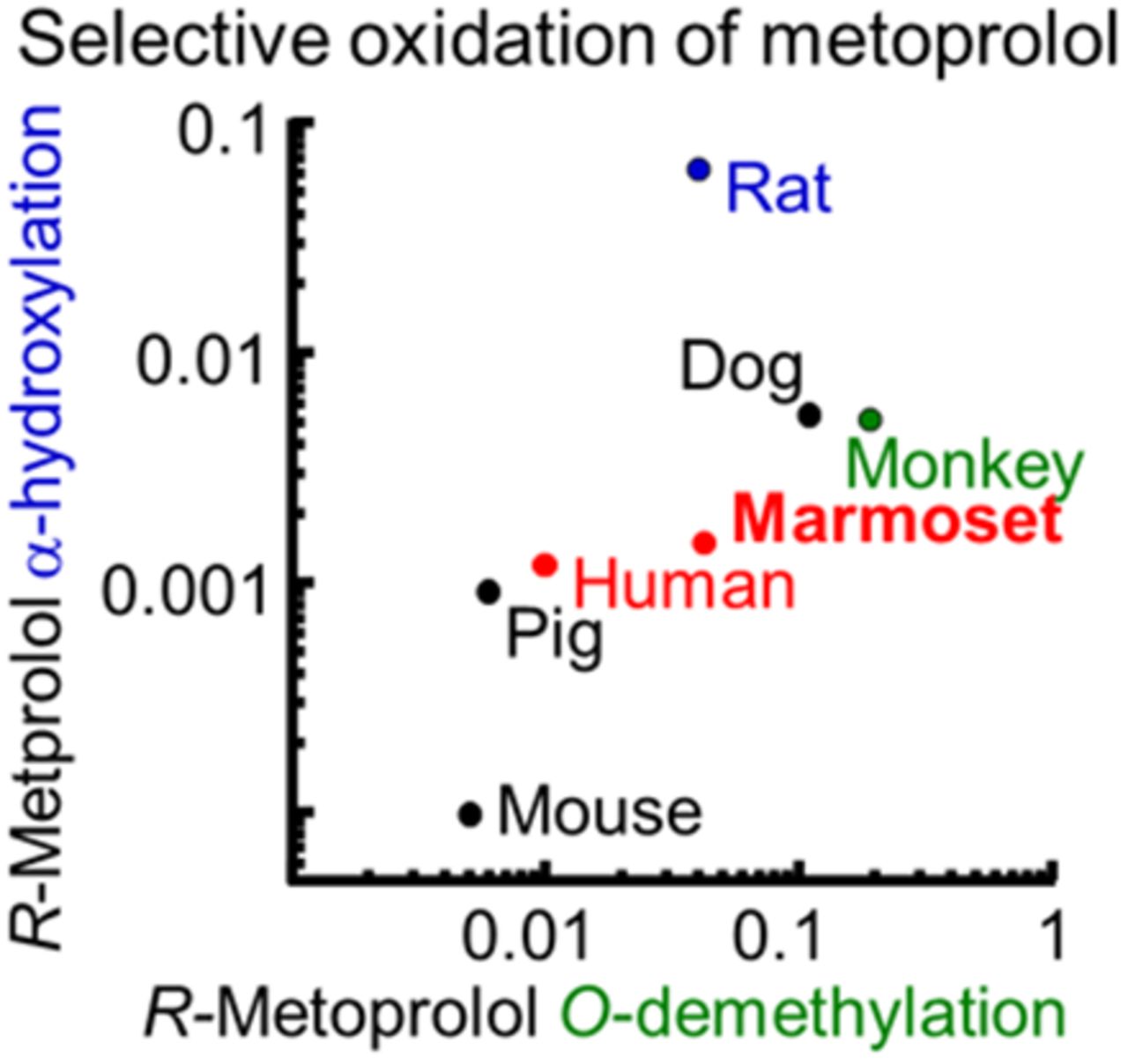
A β-blocker, metoprolol, is one of the in vivo probes for human cytochrome P450 (P450) 2D6. Investigation of nonhuman primate P450 enzymes helps to improve the accuracy of the extrapolation of pharmacokinetic data from animals into humans. Common marmosets (Callithrix jacchus) are a potential primate model for preclinical research, but the detailed roles of marmoset P450 enzymes in metoprolol oxidation remain unknown. In this study, regio- and stereo-selectivity of metoprolol oxidations by a variety of P450 enzymes in marmoset and human livers were investigated in vitro. Although liver microsomes from cynomolgus monkeys and rats preferentially mediated S-metoprolol O-demethylation and R-metoprolol α-hydroxylation, respectively, those from humans, marmosets, minipigs, and dogs preferentially mediated R-metoprolol O-demethylation, in contrast to the slow rates of R- and S-metoprolol oxidation in mouse liver microsomes. R- and S-metoprolol O-demethylation activities in marmoset livers were strongly inhibited by quinidine and ketoconazole, and were significantly correlated with bufuralol 1′-hydroxylation and midazolam 1′-hydroxylation activities and also with P450 2D and 3A4 contents, which is different from the case in human livers that did not have any correlations with P450 3A–mediated midazolam 1′-hydroxylation. Recombinant human P450 2D6 enzyme and marmoset P450 2D6/3A4 enzymes effectively catalyzed R-metoprolol O-demethylation, comparable to the activities of human and marmoset liver microsomes, respectively. These results indicated that the major roles of P450 2D enzymes for the regio- and stereo-selectivity of metoprolol oxidation were similar between human and marmoset livers, but the minor roles of P450 3A enzymes were unique to marmosets.
Footnotes
- Received February 21, 2017.
- Accepted May 10, 2017.
This work resulted from “Construction of System for Spread of Primate Model Animals” under the Strategic Research Program for Brain Sciences of the Japan Agency for Medical Research and Development. S.U. was also supported partly by a Japan Society for the Promotion of Science Grant-in-Aid for Young Scientists B [15K18934].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

- Copyright © 2017 by The American Society for Pharmacology and Experimental Therapeutics
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}