


Abstract
Drug-induced cardiotoxicity may be modulated by endogenous arachidonic acid (AA)–derived metabolites known as epoxyeicosatrienoic acids (EETs) synthesized by cytochrome P450 2J2 (CYP2J2). The biologic effects of EETs, including their protective effects on inflammation and vasodilation, are diverse because, in part, of their ability to act on a variety of cell types. In addition, CYP2J2 metabolizes both exogenous and endogenous substrates and is involved in phase 1 metabolism of a variety of structurally diverse compounds, including some antihistamines, anticancer agents, and immunosuppressants. This review addresses current understanding of the role of CYP2J2 in the metabolism of xenobiotics and endogenous AA, focusing on the effects on the cardiovascular system. In particular, we have promoted here the hypothesis that CYP2J2 influences drug-induced cardiotoxicity through potentially conflicting effects on the production of protective EETs and the metabolism of drugs.
Introduction
Drug-induced cardiotoxicity affects all components and functions of the cardiovascular system. It is characterized by changes in electrocardiographic (ECG) waveform morphology, hemodynamics, pathologic damage to the myocardium and vasculature, and changes in blood function (Laverty et al., 2011). Cardiotoxicity is a major cause of attrition in preclinical and clinical drug development and may be attributed to numerous mechanisms (Pointon et al., 2013). Drug-exerting effects on the cardiovascular system have been shown to change heart contractility, cardiac rhythm, blood pressure, and ischemia (Feenstra et al., 1999). Antihistamines such as astemizole and terfenadine cause abnormalities in ECG wave intervals, such as an increase in QT interval, leading to torsades de pointes (TdP) (Zhou et al., 1999; Lu et al., 2012). The anticancer drug doxorubicin (DOX), however, causes oxidative stress that results in structural damage to the heart, leading to heart failure (Chatterjee et al., 2010). Whereas astemizole exerts it effect primarily on cardiomyocytes, DOX also affects ancillary cells such as fibroblasts, endothelial cells, and vascular smooth muscle cells, disrupting the structure and function of vascular smooth muscle cells and inducing a profibrotic phenotype in fibroblasts (Chatterjee et al., 2010).
This review focuses on cytochrome P450 (P450) 2J2 (CYP2J2), which has been shown to modulate drug-induced cardiotoxicity (Zhang et al., 2009b). The biologic role of CYP2J2 appears to relate primarily to its metabolism of arachidonic acid (AA) to cardioprotective epoxyeicosatrienoic acids (EETs). We present evidence in this review to support the proposal that a complex interplay between EET synthesis and drug metabolism by CYP2J2 exists. It is likely that EET synthesis often predominates and largely protects the cardiovascular system but that drugs might inhibit EET synthesis in a competitive or noncompetitive manner or that drug metabolism by CYP2J2, in the heart or elsewhere, might liberate cardiotoxic drug metabolites.
EETs possess pleiotropic biologic activities, including stimulation of angiogenesis, vasodilation, inhibition of vascular smooth muscle cell migration, protection against hypoxia-reperfusion injury, increased endothelial nitric oxide synthase expression and activity, and protection against DOX-induced cardiotoxicity (Larsen et al., 2007; Spector and Norris, 2007; Yang et al., 2009; Zhang et al., 2009b; Campbell and Fleming, 2010). Given these wide-ranging effects on the cardiovascular system, it is not surprising that EETs and CYP2J2 might modulate the pathogenesis of cardiovascular disease; however, understanding of the protective role of EETs during cardiotoxicity is relatively unexplored, suggesting that further studies on a range of cardiotoxic agents are worthwhile. CYP2J2 is also a drug-metabolizing enzyme and has been implicated in the biotransformation of a variety of drugs in the liver and other tissues (Michaud et al., 2010; Xu et al., 2013). We also highlight how the balance between drug metabolism and protective EET formation may influence cardiotoxicity (Fig. 1).
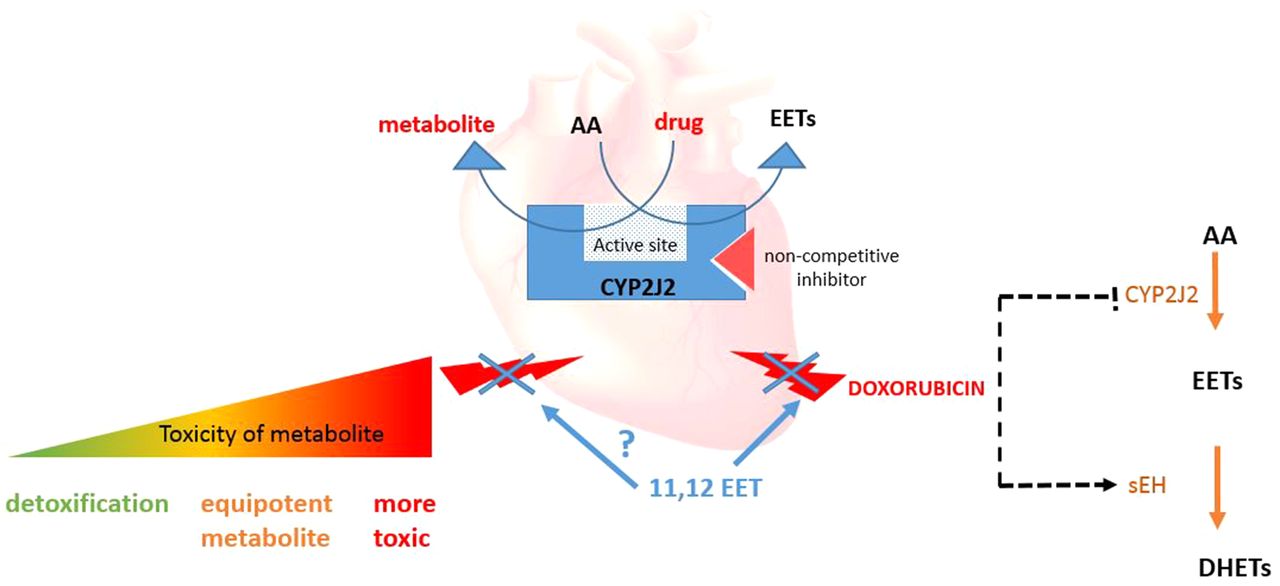
Overview of hypothesized CYP2J2 functions in the heart. CYP2J2 has a role in drug metabolism in the heart, which may lead to either detoxification of drugs or cardiotoxicity. This could be counteracted by its epoxide activity by which it produces several protective molecules, including EETs. DOX inhibits production of EETs through inhibiting epoxygenase enzymes and increasing sEH mRNA production in rats; however, the addition of exogenous EETs protects against DOX toxicity in H9c2 cells (Zhang et al., 2009b). It may be proposed that the CYP2J2/EET pathway has a role in protecting against other drug-related toxicities within the heart. Furthermore, although there is little evidence regarding drug-AA interactions, the competition between drug substrates and AA may alter the balance of protective EETs and cardiotoxic compounds. Drugs such as dronedarone have been shown to inhibit CYP2J2 in a noncompetitive way and so inhibit EET formation in an in vitro system (From Karkhanis et al., 2016, 2017).
Role of CYP2J2 in Cardiovascular Biology
The mRNA expression of CYP2J2 in humans is confined mainly to the cardiovascular system and liver, with predominant expression in the right ventricle of the heart (Michaud et al., 2010); however, mRNA has also been demonstrated in the kidney (Enayetallah et al., 2004); mRNA and protein in the lung (Zeldin et al., 1996), brain (Dutheil et al., 2009), gastrointestinal tract (Zeldin et al., 1997a), pancreas (Zeldin et al., 1997b), and some human carcinoma tissues at lower levels (Jiang et al., 2005). Discrepancies between CYP2J2 mRNA and protein expression have been found in the liver (Gaedigk et al., 2006); the consequence for the heart remains unknown. Additionally, multiple immunoreactive bands on Western blotting from extracts of adult human liver and heart have been reported. It has been hypothesized that these are uncharacterized isoforms of CYP2J2 (Wu et al., 1996; Gaedigk et al., 2006). These isoforms may possess similar or alternate activities to the main isoform of CYP2J2 and so warrant further study. Despite its elevated expression in the cardiovascular system compared with other tissues, the role of CYP2J2 in the metabolism of drugs in the heart is, to some extent, still unknown.
Results from a P450 mRNA screen showed that CYP2J2 is the predominant isoenzyme expressed in cardiomyocytes and human heart tissue (Fig. 2) (Evangelista et al., 2013). Furthermore, in line with these high levels of mRNA expression, CYP2J2 protein levels in human heart microsomes were approximately 50-fold greater than other P450 enzymes (Bylund et al., 2001; Evangelista et al., 2013). Evangelista and colleagues also established mRNA levels of the P450 enzyme CYP4F12 in the human heart. P450 enzymes belonging to the CYP4A and 4F subfamily produce 20-hydroxyeicosatetrenoic acid (20-HETE) from AA (Miyata et al., 2005; Harmon et al., 2006; Tang et al., 2010) and show increased expression in cardiovascular disease. 20-HETE has antagonistic effects toward EETs, exacerbating disease processes (Jenkins et al., 2009). It is largely unknown, however, how these other enzymes contribute to cardiovascular function and whether they have a role in cardiotoxicity.

Relative mRNA expression of P450s in human cardiomyocytes and human heart tissue and cardiomyocytes. Total RNA was extracted, and reverse-transcription polymerase chain reaction (RT-PCR) was carried out. The house keeper GusB was used a housekeeper gene, and 2ΔCT calculation was used to quantitate CYP2J2 mRNA expression. Ten P450 enzymes were investigated, of which CYP2J2 had the highest expression (Reproduced, with permission, from Evangelista et al., 2013).
In addition to cardiomyocytes, CYP2J2 expression has been observed in other cardiovascular cells. CYP2J2 is expressed in endothelial cells in a variety of vascular beds, including the coronary artery, aorta (Delozier et al., 2007), and varicose veins (Bertrand-Thiebault et al., 2004). The presence of CYP2J2 in a variety of tissues and specific expression in the heart led to the hypothesis that CYP2J2 could contribute to endogenous tissue function. Although it is not well explored, the expression of CYP2J2 in endothelial cells, fibroblasts, and smooth muscle cells (Brutsaert, 2003; Deb and Ubil, 2014) could contribute to cardiotoxicity through drug metabolism, as well as protective effects through the formation of EETs.
In endothelial cells, mRNA for another epoxygenase, CYP2C9, is highly expressed compared with CYP2J2 (Delozier et al., 2007). Moreover, CYP2C9 mRNA levels were higher both in human aorta and coronary artery than were CYP2J2 and CYP2C8. Protein analysis further reflected this, as CYP2C9 expression was dominant compared with CYP2J2, and expression of CYP2C8 was not observed (Delozier et al., 2007). Although CYP2C9 can produce EETs, it has also been reported to generate damaging reactive oxygen species (ROS), which are regarded as proinflammatory mediators, increasing nuclear factor-κB (NF-κB) activity (Fleming, 2001) and presumably leading to increased expression of proinflammatory cytokines, growth factors, and adhesion molecules, inducing an inflammatory phenotype and opposing the effects of CYP2J2 (Taniyama and Griendling, 2003).
CYP2J2 catalyzes the epoxidation of the double bonds of endogenous cellular AA to generate EETs (Fig. 3) (Zeldin et al., 1997a). Owing to the four double bonds of AA and the stereochemistry and regiochemistry associated with these bonds, eight EET isomers are possible: 5,6-EET (R/S), 8,9-EET(R/S), -11,12-EET(R/S), and 14,15-EET(R/S) (Xu et al., 2013). In vivo EETs are rapidly metabolized by soluble epoxide hydrolase (sEH) to dihydroxyeicosatrienoic acids (DHETs), which are less biologically active (Zhang et al., 2014). In addition to DHET formation, re-esterfication of EETs and incorporation into the phospholipid membrane for storage occur, allowing release and distribution (Bernstrom et al., 1992). Other relatively minor metabolism pathways of EETs have been described that involve cyclooxygenase, lipoxygenase, and P450 ω-oxidase activities (Spector et al., 2004).

The formation of EETs by CYP2J2. AA is hydrolyzed from the phospholipid bilayer and is the precursor for eicosanoids including prostaglandins, leukotrienes, and HETEs. Epoxidation by CYP2J2 leads to production of four different EETs, which can be deactivated by sEH to form DHETs.
Soluble epoxide hydrolase may have significant biologic activity in a variety of tissues, including the liver, kidney, heart, spleen, endothelium, and mammary gland (Newman et al., 2005). Its deactivation of biologically protective EETs has led to the development of pharmacologic inhibitors of sEH (Liu et al., 2009; Hwang et al., 2013) and resulted in the first sEH inhibitor, AR9281, to begin phase 2a trials for the treatment of type 2 diabetes (Anandan et al., 2011), as well as recent introduction of sEH inhibitor, GSK225629, which is currently in phase 1 trials for the treatment of hypertension (Lazaar et al., 2016) and chronic obstructive pulmonary disease (Yang et al., 2017). Furthermore, several studies have shown that sEH plays an influential role in the development of myocardial hypertrophy (Xu et al., 2006) and atherosclerosis (Zhang et al., 2009a).
Investigation of the CYP2J2 epoxidation pathway in various tissues has provided evidence to support the idea that CYP2J2, and thus EETs, have a biologically protective role, and this is emphasized by the growing potential of sEH inhibitors. Investigation of these small-molecule inhibitors suggests a link between sEH inhibition and improved cardiovascular health and, given the high expression of CYP2J2 in the human heart, suggest a possible protective role for CYP2J2 in drug-induced cardiotoxicity.
Genetic Variation in Human CYP2J2.
Epidemiologic studies conducted to examine variants in many P450 genes have found more than 2000 single nucleotide polymorphisms (SNPs) (Preissner et al., 2013). A number of these have been associated with disease. Within the Chinese population, a proximal promoter polymorphism (−50G>T), rs890293 (CYP2J2*7), has been shown to alter CYP2J2 expression (Table 3). This mutation decreases binding of Sp1 transcription factor to the promoter region of CYP2J2. As Sp1 is responsible for regulating transcriptional basal activity, blocking it results in an ∼50% reduction in promoter activity and decreased expression of the CYP2J2 gene (Spiecker et al., 2004). This polymorphism may be involved in the pathogenesis of type 2 diabetes (Wang et al., 2010), Alzheimer disease (Yan et al., 2015), and chronic kidney disease and was negatively associated with cardiovascular diseases, including myocardial infarction (Jie et al., 2010), coronary artery disease (Zhu et al., 2013), and hypertension (Wu et al., 2007) within this population. On the other hand, studies conducted in the Swedish and German populations looking at cardiovascular risk found no susceptibility to hypertension, coronary artery disease, or stroke in carriers of the rs890293 polymorphism (Hoffmann et al., 2007; Fava et al., 2010), indicating that additional association studies may be required to elucidate the risk of this CYP2J2 polymorphism for cardiovascular disease. None of the other SNPs in CYP2J2 has been associated with disease (Table 3). Furthermore, no reported associations have been found between any CYP2J2 polymorphisms and cardiotoxicity, despite some of the variants having profound effects on enzyme expression or activity in vitro. Without further study, it is unknown whether CYP2J2 polymorphisms may be important in cardiotoxicity.
CYP2J2/EETs in the Maintenance of Cardiovascular Health: a Potential Role in Cardiotoxicity?
Cardiotoxicity can be viewed as a continuum of physiologic states that shares characteristics with cardiovascular disease. Consequently, understanding the role of CYP2J2 and EETs in cardiovascular disease can provide insight into their role in cardiotoxicity. It is widely recognized that many P450 enzymes are upregulated in failing hearts (Zordoky and El-Kadi, 2008a). The upregulation of CYP2J2 and EETs is protective in the heart. In the following sections, we discuss how CYP2J2 may play a role in the modulation of vascular inflammation, vascular tone, ischemia reperfusion injury, and cardiac hypertrophy.
Vascular Inflammation.
Infiltration of inflammatory cells, particularly monocytes/macrophages, is an early event in and causal in the development of cardiovascular pathologies. Potent anti-inflammatory effects of CYP2J2 and EETs have been demonstrated both in vivo and in vitro. In vitro, synthetic EETs can reduce expression of many proinflammatory involved in the activation and adhesion of endothelial cells to leukocytes and leukocyte transmigration across the endothelium (Xu et al., 2011). For example, 11,12-EET suppresses the expression of adhesion molecules E-selectin and vascular cell adhesion molecule-1 in tumor-necrosis factor α (TNF-α)−induced human endothelial cells (Node et al., 1999). In addition to blocking the actions of TNF-α, EETs (11, 12- and 8, 9-EET) have also been shown to inhibit basal TNF-α production in THP-1 cells, a model monocytic cell line (Bystrom et al., 2011). EETs inhibit lipopolysaccharide (LPS)-induced macrophage polarization and reduce the expression of many proinflammatory cytokines while at the same time upregulating anti-inflammatory cytokine interleukin-10 in human embryonic kidney 293 cells overexpressing recombinant CYP2J2; these effects are likely mediated through downregulating NF-kβ and activation of peroxidase proliferation–activated receptors (Dai et al., 2015).
Studies in mouse models have further validated the effects of transgenic CYP2J2 and EETs in inflammation. Continuous infusion of 11,12-EET inhibited TNF-α–induced endothelial vascular cell adhesion molecule-1 expression and mononuclear cell rolling and adhesion in mouse coronary arteries. In addition, in a CYP2J2 transgenic mouse model, expression of CYP2J2 in mice significantly reduced LPS-induced production of proinflammatory mediators interleukin-6 (IL-6), monocyte chemoattractant protein-1, E-selectin, and IL-1β, as well as NF-κB activation and invasion of inflammatory cells in lung tissues (Potente et al., 2003). EETs inhibited phosphorylation of the NF-κB complex, preventing its translocation to the nucleus and hence transcriptional effects (Node et al., 1999). Transgenic CYP2J2 was also found to reduce angiotensin II–induced cardiac fibrosis and inflammation in mice, possibly though the inhibition of the NF-kβ pathway (Yang et al., 2015).
Inflammatory processes have been implicated in the development of atherosclerosis. CYP2J2 also protects against the production of atherosclerotic plaques in a transgenic mouse model of atherosclerosis (Liu et al., 2016). Apo-E–deficient mice and CYP2J2 transgenic mice were given a high-fat diet for 25 weeks. Histologic analysis of aorta samples identified fewer plaques on the luminal surface of the aorta in mice injected with CYP2J2 vector compared with wild-type. Analysis of lipids showed lower circulating triglyceride and cholesterol levels in transgenic mice. In addition, 11,-12 EET was shown to inhibit TNF-α –induced apoptosis in human umbilical vein cells through the activation of protein kinase B and FOXO1, which is downregulated in atherosclerotic aorta (Liu et al., 2016); however, it is challenging to ascertain whether these results are a direct outcome of CYP2J2-derived EETs or other protective molecules, for example, metabolites of EPA and DHA, and, moreover, to ascertain which of the EETs are responsible for the protective action.
Likewise, one of the hallmarks of some drug-induced cardiotoxicities is the increase in production of inflammatory mediators. For example, epirubicin, known to alter cardiac morphology, increases IL-6 and its soluble receptor sIL-6R, which have been shown to contribute to the pathophysiology of cardiomyopathy (Mercuro et al., 2007). It is possible that the anti-inflammatory effects of EETs could attenuate some of the myocardial damage mediated by elevated levels of these inflammatory mediators.
Vascular Tone.
Maintenance of vascular tone is critical for cardiovascular function, particularly contributing to blood pressure regulation. EETs, in particular 11,12-EET, are also known as endothelial-derived hyperpolarizing factors and cause relaxation of rat coronary arteries, as well as renal and cerebral arteries of rats and rabbits (Campbell et al., 1996; Fisslthaler et al., 1999; Imig et al., 2001; Larsen et al., 2006; Dimitropoulou et al., 2007). EETs reduce vascular tone by attenuating calcium entry via voltage-sensitive channels, leading to hyperpolarization of vascular smooth muscle cells; however, EETs may also increase intracellular calcium concentration in endothelial cells by activation of KCa channels.
Although an increase in EETs has been shown to reduce vascular tone and protect against cardiovascular disease in animal models, it is still unknown how this protective pathway may influence cardiotoxicity.
Ischemia Reperfusion Injury.
Ischemia-reperfusion injury can lead to accumulation of protective EETs after the release of fatty acids by membrane-bound phospholipases (Seubert et al., 2004). Increasing EETs in an sEH null mouse model showed limited mitochondrial damage after ischemia compared with wild-type (Akhnokh et al., 2016). EETs can enforce their cardioprotective effects through the activation of mitoK (ATP) and opening of the mitochondrial membrane permeability pore (Barau et al., 2015). Activation of this protective pathway by EETs maintains mitochondrial structure and function in CYP2J2 transgenic mice (Seubert et al., 2004). It is well known that mitochondrial ischemia reperfusion injury activates apoptosis. EETs inhibit proapoptotic pathways through the prosurvival enzyme phosphoinositide 3 kinase (PI3K) in rat cardiomyocytes (Isomoto et al., 2006).
Transgenic mice with endothelial cell–specific CYP2J2 expression showed that endothelial-derived EETs did not protect against cardiac ischemia; moreover, transgenic mice with enhanced endothelial expression of sEH showed no changes in left ventricular developed pressure (LVDP) and infarct size; however, transgenic mice with myocardial specific expression of CYP2J2 had increased recovery of LVDP and decreased infarct size after ischemia-reperfusion compared with wild-type. Furthermore, transgenic mice with expression of myocardial sEH showed no changes in LVDP and infarct size (Edin et al., 2011). These results add another layer of complexity to the protective capabilities of EETs and how EETs produced from different areas and cell types of the cardiovascular system may have varying effects.
Cardiac Hypertrophy.
Cardiac hypertrophy is a prominent risk factor for heart failure and a strong predictor of adverse cardiovascular events (Zordoky and El-Kadi, 2008a). It is normally characterized by an increase in cardiomyocyte size, increased synthesis of β natriuretic peptide (BNP), atrial natriuretic peptide (ANP), myosin, and actin accompanied by fibrosis and remodeling (Alsaad et al., 2013). Animal models have shown that during isoproterenol-induced cardiac hypertrophy, there is a decrease in protective EETs. Modulation of this process, that is, increasing the EET half-life by using sEH inhibitors, protected against the detrimental effects of cardiac hypertrophy, although the exact mechanism for this protection is yet to be determined. (Zordoky and El-Kadi, 2008a).
Animal Homologs of CYP2J2 to Investigate Preclinical Drug-Induced Cardiotoxicity
Before they can be used in humans, novel small molecules must be tested in both rodent and nonrodent in vivo models to assess for potential safety liabilities in major organs, including the heart. Many mammals possess homologs of the human CYP2J2 protein with varying sequence similarities, including the commonly used species for regulatory safety, pharmacology, and toxicology studies (in rodents, dogs, and monkeys) (Tables 1 and 2).
Amino acid sequence homology between human CYP2J2 and mammalian CYP2Js
Humans, monkeys, and dogs all have single isoforms for CYP2J2, whereas rodents (e.g., rats) have multiple isoforms, CYP2J3 and 2J4 the most similar to those in humans. Values are the percentage of sequence homology.
CYP2J gene expressions in human and other mammalian tissues
Expression is quantified, whereby (+) is low expression, (++) is moderate expression and (+++) is high expression. Within the heart, human CYP2J2 has the highest level of RNA and protein expression. Rat CYP2J3 has high levels of protein and mouse CYP2J11 has high RNA expression in the heart, indicating that there are often discrepancies between RNA and protein expression in different homologs.
Compared with the single CYP2J2 gene in humans, mice have a cluster of CYP2J isoform genes. This subfamily is highly homologous: 62%–84% homology at the amino acid level compared with humans. Mice CYP2J isoforms are distributed in the liver, kidneys, intestine, brain, and lung and abundantly in the heart (Graves et al., 2013). All enzymes produced from the CYP2J cluster have similar substrate preference, but the products have a unique profile (Nelson et al., 2004). Compared with recombinant CYP2J2 microsomes, all isoforms have been shown to metabolize AA, albeit at a lower rate (Graves et al., 2013).
Mouse models are used to investigate cytochrome P450-dependent metabolism. Knockout and transgenic mice are used to study the metabolism pathways pertaining to a specific enzyme leading to toxicity. For example, knockout and humanized mouse models for CYP2E1 have been used to characterize acetaminophen hepatotoxicity (Gonzalez et al., 2015). Currently, transgenic mouse models for CYP2J have been created to understand the biologic significance of EETs in disease; however, studies specifically addressing the role of CYP2J in the induction of toxicity have not yet been described. In vitro, mouse-derived HL-1 cell lines have limited cardiac morphologic, biochemical, and electrophysiologic properties compared with human adult cardiomyocytes; however, their expression of P450 enzymes has not been clearly investigated and so may offer a potential in vitro cardiac model for studying CYP2J.
The main rat homologs, CYP2J3 and CYP2J4, have 72% and 76% sequence similar to human CYP2J2, and have a similar tissue distribution to human. Furthermore, CYP2J3 is reported to be found primarily within atrial and ventricular myocytes (Wu et al., 1997; Zhang et al., 1997), whereas increased expression of CYP2J3 in the heart after ischemic postconditioning significantly increased EET generation (Wang et al., 2012), suggesting that CYP2J3 may have epoxygenase activity analogous to CYP2J2. Therefore, rat CYP2J3 may be the closest homologous enzyme to CYP2J2 in terms of distribution and epoxygenase activity and may be applicable to investigations of cardiotoxicity. Also, rat cardiac cell lines, such as rat myoblast H9c2 cells, have been used to investigate cardiac biology and toxicology; however, they lack key functional features of cardiomyocytes, exhibit a mainly skeletal muscle phenotype, and do not respond to electrical stimulation (Kimes and Brandt, 1976); however, they have recently been used to investigate the cardioprotective effects of drugs after oxidative damage (Zhou et al., 2016), thus suggesting the potential use of these cells in investigating the mechanisms of cardioprotection from drug-induced cardiotoxicity.
In both dogs and monkeys, a single CYP2J isoform (CYP2J2) has been identified (Nelson, 2009). Monkey CYP2J2 has the greatest sequence similarity (95%) to CYP2J2 in humans (Uno et al., 2007). Immunoquantification of cynomolgus CYP2J2 identified higher levels of protein in the monkey liver compared with the human liver (Uehara et al., 2015), as reflected in activity where there was a higher hepatic clearance of astemizole (Nishimuta et al., 2011), which has been shown to be a drug substrate for CYP2J2 (Uehara et al., 2015). To our knowledge, however, expression of CYP2J2 at the mRNA or protein level has not yet been quantified in the monkey or dog heart.
Cardiotoxicity encompass a variety of features, including changes in pathology, ECG, and hemodynamics. Therefore, some animal models may be recommended for investigating functional changes, whereas other models may be more suited to examining pathologic changes. For example, rats, which reflect the protective capabilities of CYP2J, would be acceptable for studying EETs; however, their use in predicting cardiotoxicity associated with ion channel inhibition is limited. Consequently, when selecting a suitable preclinical model system, both the potential ability to modulate CYP2J2 and the expected cardiac effects being risk assessed should be taken into consideration in selecting the most appropriate approach and species. Furthermore, although the dog has a single CYP2J2 isoform with high sequence similarity, more studies are required to fully determine whether the dog is the best model to study CYP2J2 from both cardiovascular biology and CYP2J2 perspectives (Table 3).
CYP2J2 allelic variations in humans known to have changes in activity in vitro
Currently only a variation in the CYP2J2*7 allele has been shown to have associations with disease.
Role of CYP2J2 in Xenobiotic Metabolism
In the human liver, CYP2J2 protein comprises 1% to 2% of total P450 content, similar to that in the small intestine (1.4%) (Paine et al., 2006) compared with CYP3A4, which constitutes ∼30% of total P450 content (Michaels and Wang, 2014); however, as CYP3A4 expression is low in cardiac tissue, it is unlikely to contribute to drug metabolism within the heart (Chaudhary et al., 2009). Although not the most highly expressed P450 in the liver and intestine, CYP2J2 mediates drug biotransformation reactions with numerous exogenous substances. CYP2J2 is the primary enzyme involved in several metabolic reactions, including amiodarone 4-hydroxylation, astemizole O-demethylation, and ebastine hydroxylation (Matsumoto and Yamazoe, 2001; Matsumoto et al., 2002; Liu et al., 2006). For ebastine, CYP2J2 plays a superior role in first-pass intestinal metabolism to its pharmacologically active metabolite and less toxic carebastine. All three of these compounds are known cardiotoxins. Both astemizole and ebastine block the Kv11.1 (hERG) potassium channel, causing TdP and QT prolongation arrhythmias; however both amiodarone and astemizole metabolites are as toxic as the primary compound when metabolized (Matsumoto et al., 2002). Thus, the individual product profile of a compound resulting from CYP2J2 metabolism may lead to less toxic or equipotent metabolites and altered toxicity of these drugs within the heart.
A study in human liver microsomes identified eight novel substrates for CYP2J2 after screening 139 compounds, including marketed therapeutic agents. These chemical entities had wide structural diversity and ranged from small molecules like albendazole to larger complex structures such as cyclosporine (Lee et al., 2010). This diversity in drug substrates highlights how CYP2J2 may be vital in mediating drug responses and gives a glimpse into the similarity between CYP2J2 and other P450 enzymes of similar function. CYP2J2 and CYP3A4 share several substrates, including antihistamines (terfenadine, astemizole, and ebastine), anticancer drugs (DOC and tamoxifen), and immunosuppressants (cyclosporine); a list of known CYP2J2 substrates is shown in Table 4. In silico approaches suggest structural similarity between CYP2J2 and CYP3A4, and a comparison of active sites showed homology; however, further examination shows slight differences in structural geometry. CYP2J2 has a more cylindrical shape and is narrower than CYP3A4 as the β-4 part of the protein is smaller and has a loop that inserts into the active site restricting metabolism (Lee et al., 2010).
Range of substrates for CYP2J2 and the metabolic pathways by which they are formed
Substrates include both endogenous compounds and exogenous drugs and the main metabolic pathways are hydroxylation and epoxygenation.
Biotransformation studies looking at the metabolism profile of CYP2J2 and CYP3A4 showed that CYP2J2 produced numerous metabolites, many of which were also observed with CYP3A4 (Fig. 4); however, indications of differences were found in the regioselectivity in metabolites from albendazole, amiodarone thioridazine, mesoridazine, danazol, and astemizole after incubation with the two enzymes. Evidence showed that some metabolites were produced exclusively by CYP2J2. (Lee and Murray, 2010). Further investigation by Kaspera et al. (2014) showed the significant contribution of CYP2J2 to ritonavir metabolism in the liver, with a unique metabolism profile compared with CYP3A4/5. CYP2J2 produced specific metabolites from oxidation of the thiazole rings on different sides of the molecule. This study found that CYP2J2 had a greater affinity for ritonavir (Km 0.016 µM) compared with CYP3A4 (Km 0.068 µM) and CYP3A5 (Km 0.047 µM) in liver microsomes (Kaspera et al., 2014). In addition, CYP2J2 and CYP2C19 were the major enzymes responsible for the metabolism of albendazole and fenbendazole in human liver microsomes. Both these drugs can be transformed to their sulfoxide and hydroxyl metabolites (Wu et al., 2013). CYP3A4 and flavin-containing monooxygenase are thought to be major enzymes in producing sulfoxide metabolites (Virkel et al., 2004); however, Wu et al., (2013) demonstrated CYP2J2 to be the primary enzyme mediating albendazole hydroxylation. CYP2C19 and CYP2E1 also contributed to this hydroxylation but to a lesser extent. (Wu et al., 2013). The consequences of the formation of these specific metabolites by CYP2J2 has yet to be fully determined.

Chromatographic data showing the differences in products formed by recombinant CYP3A4 and CYP2J2. Albendazole metabolism by both CYP3A4 and CYP2J2 produced the same metabolite; however, an additional metabolite was observed for CYP2J2, indicating hydroxylation. CYP2J2 and CYP3A4 produced similar peaks, indicating hydroxylation. Chromatographic data of astemizole metabolism showed that CYP3A4 produced the hydroxylated product, whereas metabolism by CYP2J2 resulted in both the o-dealkylated metabolite and some N-dealkylated product. When danazol was incubated with both enzymes, similar metabolites were produced. (Reproduced, with permission, from Lee et al., 2010).
Several CYP2J2 substrates are known to have pharmacologic effects in the heart and may be metabolized in this tissue (Evangelista et al., 2013). Applying this logic, CYP2J2 may be able to regulate the local concentrations of these compounds and therefore modulate cardiotoxicity. Studies in heart microsomes incubated with verapamil led to the formation of nine P450 metabolites. As verapamil is an L-type calcium channel blocker, which is commonly prescribed for heart conditions such as angina and arrhythmias, CYP2J2 may be able to regulate functional activity of the drug (Michaud et al., 2010). Furthermore, in isolated rat heart hydroxylation of the H1 receptor antagonist, ebastine to hydroxyebastine and carebastine was detected and compared with human liver microsomes and showed a similar metabolism profile; however, as there was no comparison with metabolism in the human heart, it is difficult to ascertain comparative activity between CYP2J2 and CYP2J3. It is also unclear whether other P450 enzymes could be responsible, in part, for ebastine metabolism (Kang et al., 2011). Overall, the metabolic activity of CYP2J2 in the liver and its ability to metabolize a wide array of drugs, coupled with its high expression in the heart, warrant further studies to clarify the significance of cardiac CYP2J2 in drug metabolism in physiologic relevant systems.
Modulation of CYP2J2 Activity by Drugs
Evangelista et al. (2013) investigated the role of different drugs in the induction and inhibition of CYP2J2 in adult primary human cardiomyocytes. A major limitation of this research is that the cells used in this study can divide and are morphologically and functionally different compared with freshly isolated cells. CYP2J2 activity was measured via terfenadine hydroxylation at two different inhibitor concentrations. The most potent inhibitor of CYP2J2 tested was danazol, which reduced activity by ∼95%; other less potent inhibitors included ketoconazole and astemizole (Evangelista et al., 2013). In addition to the inhibitors of CYP2J2 recognized in this review, other drugs are reviewed elsewhere (Lafite et al., 2007; VanAlstine and Hough, 2011).
A more recent study highlighted the reversible mixed-mode inhibition of recombinant CYP2J2 by dronedarone (Ki = 0.034 µM), amiodarone (Ki = 4.8 µM), and their active metabolites, N-desbutyldronedarone (NDBD) (Ki = 0.55 µM) and N-desethylamiodarone (NDEA) (Ki = 7.4 µM) and further irreversible inhibition by dronedarone and NDBD (Karkhanis et al., 2016). Both these drugs are multi-ion channel blockers designed to reduce cardiac arrhythmias but paradoxically are potentially also cardiotoxic, resulting in bradycardia, hypotension, congestive heart failure, and ventricular tachycardia (Dixon et al., 2013). Dronedarone has been demonstrated to cause a reduction in a recurrent atrial fibrillation in patients compared with amiodarone (Piccini et al., 2009) but can lead to QT prolongation and subsequently TdP in some instances (Heijman et al., 2013). As both amiodarone and dronedarone are substrates for CYP2J2, the interaction between these drugs and CYP2J2 may modulate cardiac side effects and drug-drug interactions, leading to further toxicity. Dronedarone, amiodarone, and their respective metabolites have been shown to inhibit CYP2J2-mediated AA metabolism and production of EETs, NDBD being the most potent (Karkhanis et al., 2017). Because of the protective role of CYP2J2 in cardiac function, it is feasible that the inhibition of CYP2J2 may block epoxygenase activity and reduce protective EETs, although our understanding of the interplay between drug metabolism and EET formation is rudimentary. In addition, it is possible that modulation of CYP2J2 activity may lead to altered levels of toxic parent compound or toxic metabolite and changes in the toxicity profile. These concepts are worthy of further study and analysis.
Potential Role of EETs in Mitigating Drug-Induced Cardiotoxicity
The mechanisms by which compounds cause cardiotoxicity resulting in changes in ECG waveforms/intervals, hemodynamics, or cardiac pathology are diverse and compound-specific. Each of these outcomes can be further defined both in terms of the physiologic and pathologic state and the molecular mechanism and or pathways by which perturbations arise. Currently, the level of molecular understanding varies considerably, depending on the physiologic or pathologic perturbation. For example, the ion channels behind the cardiac action potential, and thus changes in the ECG, are well defined. In contrast, multiple mechanisms are proposed for changes in cardiac pathology. For example, the anticancer tyrosine kinase inhibitor sunitinib causes oxidative stress (Aparicio-Gallego et al., 2011) and cardiac hypertrophy (Maayah et al., 2014); whether these perturbations are linked and whether upstream unidentified mechanisms exist remain to be determined; however, with this diversity of molecular mechanisms responsible for cardiotoxicity, the role of EETs in cardioprotection is just as diverse (Table 5). In an animal model of isoproterenol-induced cardiac hypertrophy, the use of sEH inhibitors increased the levels of EETs and decreased the induction of ANP and BNP and EPHX2 mRNA (Althurwi et al., 2013). Furthermore, increased circulating EETs reduced oxidative stress and increased expression of antioxidant enzymes (Wang et al., 2014). Therefore, some evidence suggests that protection against sunitinib (and other drug) toxicity may be achieved by an upregulation of these epoxygenases with resultant promotion of EET formation.
Proposed mechanisms of protection by EETs against selected cardiotoxic drugs
The listed compounds are known substrates for CYP2J2.
Other cardiotoxic drugs, such as the antiarrhythmic drug amiodarone, alter apoptotic pathways, increasing caspase 3 activity and leading to an increase in apoptosis and cell death in h9c2 cells (Isomoto et al., 2006). The addition of exogenous EETs in vitro boosts activity of the prosurvival enzyme PI3K and inhibits proapoptotic pathways in primary cardiomyocytes derived from a rat heart hypoxia/reperfusion model (Dhanasekaran et al., 2008).
Cardiotoxic drugs also impact hemodynamics within the cardiovascular system. An example of detrimental effects of drugs on blood pressure is the CYP2J2 substrate 5-fluorouracil, which has been shown to be vasoconstrictive through the inhibition of nitric oxide synthase, leading to coronary spasms, and via protein kinase C, leading to vasoconstriction (Alter et al., 2006). Inhibition of sEH increased EETs and dilated human coronary arterioles through BKca channels, and 11,12-EET also caused relaxation of rat coronary arteries and renal and cerebral arteries of rats and rabbits. (Campbell et al., 1996; Fisslthaler et al., 1999, Imig et al., 2001; Larsen et al., 2006; Dimitropoulou et al., 2007). Taken together, EETs act on several pathways involved in cardiotoxicity, and there is circumstantial yet plausible evidence to suggest a protective impact of CYP2J2 via EET synthesis in the heart during cardiotoxicity.
Role of CYP2J2 in DOX-Induced Cardiotoxicity
The strongest case for a role of CYP2J2 in protection against cardiotoxicity has been presented for DOX. DOX is an anthracycline used for the treatment of solid tumors and hematologic carcinomas, which has been shown to modulate CYP2J2 production of EETs. Despite its anticancer action, the clinical value of this drug is reduced owing to acute and chronic, cumulative, and irreversible dose-dependent cardiotoxicity (Belham et al., 2007). Cardiovascular effects include acute cardiomyopathy, chronic heart failure, ventricular dysfunction, and arrhythmias (Yeh et al., 2004). Currently, it is unclear whether the mechanism of cardiotoxicity is the same when occurring acutely (within days) or chronically (years) after treatment (Takemura and Fujiwara, 2007).
Preclinically, both acute and chronic DOX administration have been associated with changes in EET formation (Zordoky et al., 2010; Alsaad et al., 2012). In rats, acute DOX treatment altered the mRNA expression of P450 and sEH enzymes in kidney and liver, leading to decreases in 5,6-, 8,9-, 11,12-, and 14,15-EET (Zordoky et al., 2010). Ichikawa et al. (2014) found that acute DOX-induced cardiotoxicity was associated with generation of ROS, cellular iron accumulation, and disruption of the mitochondria, which in turn initiated apoptotic pathways in isolated neonatal rat cardiomyocytes (Ichikawa et al., 2014). Previously, EETs have been shown to attenuate production of ROS and mitochondrial dysfunction in carcinoma cells treated with arsenic trioxide (Liu et al., 2011), and an interesting and potentially important study suggested that CYP2J2 mRNA expression increased in adult human ventricular myocytes in culture in response to both external oxidants and addition of DOX; the latter, among other effects, stimulates intracellular oxidant production. Moreover, cell survival decreased with oxidant exposure when CYP2J2 was inhibited either using danazol or by siRNA for CYP2J2. Not surprisingly, this ROS toxicity was mitigated with the addition of exogenous EETs. Although the report is intriguing and plausible, there are limitations to be highlighted. The use of millimolar concentrations of pyruvate as the only antioxidant trialed limits mechanistic interpretation, as does the use of danazol as a CYP2J2 inhibitor; however, in support of CYP2J2-derived EETs as protective agents, knockdown of CYP2J2 mRNA also negatively affected cell survival after DOX, yet levels of CYP2J2 protein were not addressed. The relatively nonspecific measurement of intracellular oxidant formation resulting from DOX treatment of cells also limits interpretation of the mechanism of molecular signaling. Nevertheless, this mechanism may have implications for other cardiotoxic compounds acting, in part, via oxidant formation in cells of the cardiovascular system. If the mechanism is proven to be mediated by increased oxidants in cells, then protection from these agents could also be afforded by changes in CYP2J2 gene expression and the resultant increased EET formation.
Alsaad et al. (2012) studied the effects of chronic DOX cardiotoxicity in the heart on the mRNA expression of proteins involved in the formation of AA metabolites, as well as levels of these metabolites via liquid chromatography/mass spectroscopy in rats. Animals were treated with multiple intraperitoneal injections over 14 days, followed by a 14-day recovery period, emulating the clinical administration (Alsaad et al., 2012). Chronic DOX treatment in vivo caused no change in mRNA expression of the rat analog of CYP2J2, CYP2J3; however, it increased mRNA expression of other P450 enzymes, including CYP4A3, CYP4F1, and CYP4F5, known to produce the alternative AA metabolite 20-HETE, resulting in increased levels of 20-HETE compared with an untreated control group (Alsaad et al., 2012). These investigators also observed a rise in the mRNA expression of the EPHX2 (sEH) gene in the heart in DOX-treated rats, accompanied by decreased EET levels and an increase in the levels of inactive DHETs. Furthermore, treatment of H9c2 cells, a rat cardiac cell line, with a sEH inhibitor (t-AUCB), in combination with DOX, reduced both ANP and BNP release (markers of cardiac hypertrophy), suggesting a cardioprotective effect of sEH inhibition. Additionally, DOX upregulates levels of many P450 enzymes in this cell model, including 2J3 and 2E1 (Zordoky and El-Kadi, 2008b). Given the biologic protective activity of EETs, particularly 11,12-EET and 14,15-EET against cardiovascular disease (Sudhahar et al., 2010), it is hypothesized that EETs may provide protection against cardiotoxicity induced by a whole spectrum of xenobiotics in vivo.
Discrepancies in mRNA expression between acute and chronic DOX administration have been documented (Zordoky et al., 2010; Alsaad et al., 2012), highlighting the caution required when interpreting studies comparing acute versus chronic DOX administration. Furthermore, the treatment period used by Alsaad and colleagues may not be sufficient to induce many of the characteristic features of chronic DOX cardiotoxicity. A detailed assessment by Cove-Smith et al. (2014) of both cardiac morphology and function in rats for 8 weeks after 1.25 mg/kg DOX demonstrated changes in cardiac function, in particular, cardiac output, stroke volume, and ejection fraction from day 15. Gross morphologic changes and biomarkers associated with cardiomyocyte degeneration occurred much later (Cove-Smith et al., 2014). Consequently, the changes observed after 2 weeks of DOX treatment are unlikely to be a true indication of the protective role of EETs during DOX cardiotoxicity, and therefore the study length would need to be extended to determine this role definitively; however, it is plausible to suggest that epoxygenase enzymes are involved in the cellular development of DOX-induced cardiotoxicity.
Using transgenic mice with cardiomyocyte-specific overexpression of human CYP2J2, Zhang et al. (2009b) identified the possible beneficial effects of EETs in the protection against DOX-induced cardiotoxicity. Elevations in serum LDH and CK and activation of cardiac caspase 3 and catalase caused by acute treatment with DOX (0, 5 and 15 mg DOX/kg daily i.p. for 3 days followed by 24-hour recovery) were mitigated in mice overexpressing CYP2J2, suggesting protection against DOX-induced myocardial damage. Some of these markers are not specific to the heart, however, and this may affect the interpretation of these findings. After chronic treatment with DOX (0, 1.5, 3 mg/kg biweekly for 5 weeks followed by a 2-week recovery), the CYP2J2 transgenic mice showed a lower heart weight-to-body ratio, reduced cardiac ankyrin repeat protein (CARP), and expression ratio of βMHC:αMHC, and no change in LVDP compared with wild-type mice. This finding indicates that cardiac structure and contractile function that may have been mediated by protective EETs (Zhang et al., 2009b) were preserved. Although cardiac ankyrin repeat protein is a specific biomarker of cardiac hypertrophy, it may not be a good biomarker of early cardiac remodeling. In fact, many of the serologic, pathologic, and functional biomarkers of early cardiac damage during cardiotoxicity were not reported. For example, cardiac troponin, which is known to be the gold standard in investigating cardiac damage preclinically and clinically, was not measured (Cove-Smith et al., 2014).
Studies have found that CYP2J2 is capable of metabolizing DOX (Zhang et al., 2009b); however, as this was a CYP2J2 transgenic model and not a null model, other P450 enzymes may also be responsible for the increased metabolism of DOX. Furthermore, increased expression and activity of reductase in the transgenic mouse hearts may also lead to an increase in activity of many other P450 enzymes. It is known that the major metabolite of DOX, doxorubicinol (DOXol), contributes to toxicity in the myocardium; however, other evidence suggests other metabolites of DOX, including DOX deoxyaglycone and DOXol hydroxyaglycone, may also contribute to the cardiotoxicity (Licata et al., 2000). In addition, P450 reductase can metabolize DOX to 7-deoxydoxyrubicin aglycone, which has been shown to inhibit AA metabolism, reducing the production of EETs and altering the regiosomers of EETs produced (Arnold et al., 2017). Therefore, increased metabolism through CYP2J2 may have a conflicting effect on the heart, elevating levels of toxic metabolites for certain drugs but increasing protection through EETs. Other P450 enzymes belonging to the murine CYP2J family, including CYP2J8, 2J11, 2J12, and 2J13, also have epoxidase activity toward AA, which may confound results, especially as expression levels of these enzymes were not measured in the CYP2J2 transgenic mice (Graves et al., 2013). In rats, only CYP2J3 has been demonstrated to possess expoxygenase activity, and so protective effects by EETs may be less apparent. In addition, the use of transgenic models where there is a genetic alteration, leading to differences in protein expression and hence activity, may not directly model pharmacologic activation of CYP2J2 in the wild-type mouse (Knight and Shokat, 2007).
Taken together, the limited number of studies on DOX toxicity suggests that CYP2J2 expression and EET production have profound cardioprotective effects, modulating DOX cardiotoxicity through their influence on molecular pathways involved in apoptosis, ROS generation, hypertrophy, and cardiac remodeling, leading to an overall conservation of structure and function. It may be hypothesized that the cardiac potential of other drugs might be affected by CYP2J2 metabolism and/or EET formation. The effects of EETs on DOX raises the possibility that the CYP2J2/EET pathway may also modulate the cardiotoxicity of other drugs, working through different mechanisms (Table 5).
Conclusions
CYP2J2 is an AA and drug-metabolizing enzyme highly expressed in the human heart; however, the role of CYP2J2 in drug metabolism in the heart has not been well defined. Nevertheless, there is increasing awareness that many current drugs are substrates for CYP2J2. Substantial data suggest that CYP2J2, along with CYP3A4, plays a significant part in the metabolism of drugs known to cause cardiotoxicity. Therefore, changes in the expression or activity of this enzyme may alter drug concentrations in the body, leading to either an ineffective drug response or increased levels of metabolites, thereby leading to potential cardiotoxicity. Known P450 inducers do not modulate CYP2J2 levels, but some compounds have been recognized to induce the expression and activity of CYP2J2 in adult human cardiomyocytes. In addition, several SNPs in the human CYP2J2 gene are associated with altered in vitro activity of the CYP2J2 enzyme, which may lead to changes in EET formation and drug metabolism, potentially altering cardiotoxicity in some individuals. In addition to its role in drug metabolism, CYP2J2-derived EETs have also been shown to have a protective effect, although this has only really been reported for DOX-induced cardiotoxicity. Given that our understanding of the role of CYP2J2 and EET formation is based largely on the data from this single drug and the importance of cardiotoxicity to drug development and application in humans, understanding more fully the role and possible protection by CYP2J2 in the heart is worthy of further study.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Solanki, Pointon, Jones, Herbert.
Footnotes
- Received October 13, 2017.
- Accepted April 19, 2018.
A.P. and B.J. are employees and shareholders of AstraZeneca. M.S. is in receipt of a Biotechnology and Biological Sciences Research Council (BBSRC)–funded PhD CASE award in collaboration with AstraZeneca [Grant BB/M503368/1].
Abbreviations
- AA
- arachidonic acid
- ANP
- atrial natriuretic peptide
- BNP
- β-natriuretic peptide
- CYP2J2
- cytochrome P450 2J2
- DHET
- dihydroxyeicosatrienoic acid
- DOX
- doxorubicin
- ECG
- electrocardiographic
- EET
- epoxyeicosatrienoic acid
- 20-HEET
- 20-hydroxyeicosatetraenoic acid
- LPS
- lipopolysaccharide
- LVDP
- left ventricular developed pressure
- NDBD
- N-desbutyldronedarone
- NF-κB
- nuclear factor-κB
- P450
- cytochrome P450
- PI3K
- phosphoinositide 3 kinase
- ROS
- reactive oxygen species
- sEH
- soluble epoxide hydrolase
- SNP
- single nucleotide polymorphisms
- TdP
- torsades de pointes
- TNF
- tumor necrosis factor
- Copyright © 2018 The Author(s).
This is an open access article distributed under the CC BY Attribution 4.0 International license.
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- Introduction
- Role of CYP2J2 in Cardiovascular Biology
- CYP2J2/EETs in the Maintenance of Cardiovascular Health: a Potential Role in Cardiotoxicity?
- Animal Homologs of CYP2J2 to Investigate Preclinical Drug-Induced Cardiotoxicity
- Role of CYP2J2 in Xenobiotic Metabolism
- Modulation of CYP2J2 Activity by Drugs
- Potential Role of EETs in Mitigating Drug-Induced Cardiotoxicity
- Role of CYP2J2 in DOX-Induced Cardiotoxicity
- Conclusions
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters