


Visual Overview

Abstract
Curcumin, a major polyphenol present in turmeric, is predominantly converted to curcumin-O-glucuronide (COG) in enterocytes and hepatocytes via glucuronidation. COG is a principal metabolite of curcumin in plasma and feces. It appears that the efflux transport of the glucuronide conjugates of many compounds is mediated largely by multidrug resistance–associated protein (MRP) 3, the gene product of the ATP-binding cassette, subfamily C, member 3. However, it is currently unknown whether this was the case with COG. In this study, Mrp3 knockout (KO) and wild-type (WT) mice were used to evaluate the pharmacokinetics profiles of COG, the liver-to-plasma ratio of COG, and the COG-to-curcumin ratio in plasma, respectively. The ATP-dependent uptake of COG into recombinant human MRP3 inside-out membrane vesicles was measured for further identification, with estradiol-17β-d-glucuronide used in parallel as the positive control. Results showed that plasma COG concentrations were extremely low in KO mice compared with WT mice, that the liver-to-plasma ratios of COG were 8-fold greater in KO mice than in WT mice, and that the ATP-dependent uptake of COG at 1 or 10 μM was 5.0- and 3.1-fold greater in the presence of ATP than in the presence of AMP, respectively. No significant differences in the Abcc2 and Abcg2 mRNA expression levels were seen between Mrp3 KO and WT mice. We conclude that Mrp3 is identified to be the main efflux transporter responsible for the transport of COG from hepatocytes into the blood.
SIGNIFICANCE STATEMENT This study was designed to determine whether multidrug resistance–associated protein (Mrp) 3 could be responsible for the efflux transport of curcumin-O-glucuronide (COG), a major metabolite of curcumin present in plasma and feces, from hepatocytes into the blood using Mrp3 knockout mice. In this study, COG was identified as a typical Mrp3 substrate. Results suggest that herb-drug interactions would occur in patients concomitantly taking curcumin and either an MRP3 substrate/inhibitor or a drug that is predominantly glucuronidated by UDP-glucuronosyltransferases.
Introduction
Curcumin (CUR), the main bioactive ingredient of the spice turmeric (Curcuma longa L.), possesses a broad spectrum of biological and pharmacological activities, including but not limited to antioxidant, anticancer, anti-inflammatory, antidiabetic, hepatoprotective, and lipid-lowering effects (Ozawa et al., 2017; Farzaei et al., 2018; Huang et al., 2018; Kunati et al., 2018). However, CUR exhibits extremely poor systemic bioavailability (< 1%) in rodents and humans because of its alkaline instability, poor aqueous solubility, low oral absorption, and rapid metabolism (Anand et al., 2007). Despite its limited use in clinical settings (Garcea et al., 2005; Irving et al., 2013), CUR is still extensively used as an herbal medicine (e.g., a cancer chemopreventive agent) and as a dietary supplement for food coloring in Southeast Asian areas (Mahale et al., 2018b).
Evidence has demonstrated that oral CUR is principally converted to curcumin-O-glucuronide (COG) (see Fig. 1) by certain UDP-glucuronosyltransferases (UGTs) (Hoehle et al., 2007; Lu et al., 2017). In addition, CUR is reduced by certain reductases to dihydrocurcumin, tetrahydrocurcumin, and hexahydrocurcumin (Mahale et al., 2018a). Of these, COG is a primary circulating metabolite in rodents and humans (Ireson et al., 2002; Mahale et al., 2018b). Studies have further demonstrated that CUR metabolites (predominantly COG) activate CYP3A4 activity (Hsieh et al., 2014) and that oral intake of CUR would significantly decrease the systemic bioavailability of CYP3A4 substrate drugs, such as everolimus (an immunosuppressant) (Hsieh et al., 2014). On the other hand, as an inhibitor of UGT1A and of nonselective kinase (Abe et al., 2011), CUR suppresses the glucuronidation of many (if not all) compounds in the body. For example, concomitant use of CUR decreases glucuronidation of acetaminophen (Volak and Court, 2010) and mycophenolic acid (Basu et al., 2007). Clearly, thorough elucidation of the transport and elimination of COG would help us better understand herb-drug interactions involved and their underlying mechanisms.

Chemical structures of curcumin (A) and curcumin-O-glucuronide (B).
COG is cleared predominantly via biliary and fecal excretion, with evidence of the presence of enterohepatic recycling (Sharma et al., 2007; Bangphumi et al., 2016), a process in which the canalicular membrane efflux transporters multidrug resistance–associated protein (MRP) 2 and breast cancer resistance protein (BCRP) of hepatocytes are involved (Roberts et al., 2002). Despite recent public interest and widespread consumption of CUR around the world, little is known about how COG molecules that are formed intracellularly move out for clearance.
MRP3, expressed dominantly on the basolateral membrane side of enterocytes and the sinusoidal membrane of hepatocytes, functions as an efflux transporter that mediates the efflux transport of certain compounds from hepatocytes into the blood for further renal excretion. Because MRP3 prefers transporting the glucuronide conjugates of endogenous or exogenous compounds (Zelcer et al., 2006), it was postulated that COG might be a substrate of MRP3. To test such a hypothesis, we used Mrp3 knockout (KO) and wild-type (WT) mice to evaluate differences in their pharmacokinetics parameters of COG and the liver-to-plasma ratios of CUR and those of COG, respectively, after oral administration of CUR. To further confirm the findings from mice, we measured the ATP-dependent uptake of COG by using recombinant human MRP3 inside-out membrane vesicles in vitro.
Materials and Methods
Chemicals and Reagents.
CUR and estradiol-17β-d-glucuronide (E217βG) were purchased from Sigma-Aldrich (St. Louis, MO). COG was the product of Toronto Research Chemicals (Toronto, Ontario, Canada). Hesperetin [used as the internal standard (IS)] was provided by Aladdin (Shanghai, China). Recombinant human MRP3 inside-out membrane vesicles GM0021-V (GenoMembrane, Yokohama, Japan), AMP, and ATP were supplied by Solvo Biotechnology (Szeged, Hungary). Analytical-grade formic acid, high-performance liquid chromatography–grade acetonitrile, and methanol were from Merck (Darmstadt, Germany). Deionized water was purified using the Millipore Milli-Q system (Millipore, Milford, MA).
Study Animals.
All animal care and study protocols were approved by the Experimental Animal Welfare and Ethics Committee, Nanjing Medical University, and conducted in compliance with the Guidelines for Animal Experimentation, Nanjing Medical University, China. The Mrp3 KO mice were generously supplied by the Netherlands Cancer Institute, Amsterdam, the Netherlands. The WT mice of the FVB strain were purchased from Vital River Laboratories (Beijing, China). All animals were housed in an officially accredited experimental animal facility under air-controlled conditions with a 12-hour light/dark cycle, ambient temperature maintained at 22–24°C, and relative humidity of 40%–60%. All study mice had free access to standard rodent food and tap water, but they were fasted for 12 hours before the initiation of all studies.
Pharmacokinetics Study of COG in Mice.
For mouse studies, the frequently used dosage of CUR ranges from 20 to 400 mg/kg (Szymusiak et al., 2016; Mahale et al., 2018a). Accordingly, a dose of 200 mg/kg was chosen as a moderate dose for mice in this study. Male Mrp3 KO and WT mice (aged 6–8 weeks) were administered with 200 mg/kg of CUR by oral gavage (n = 8 each) based on a similar study (Szymusiak et al., 2016). After oral dosing, a series of blood samples (30 μl each) were withdrawn from the orbital venous plexus at 0.083, 0.25, 0.5, 1, 2, 4, 8, 12, and 24 hours, respectively, and collected into heparinized tubes. Plasma samples were obtained after centrifugation at 4000 rpm for 10 minutes and stored at −80°C until analysis.
Quantification of CUR and COG in Plasma and Liver Samples of Mice.
Again, male Mrp3 KO and WT mice (aged 6–8 weeks; n = 8 each) were administered with a single dose of CUR at 200 mg/kg by oral gavage. Thereafter, blood samples (100 μl each) were withdrawn from the orbital venous plexus at 1 hour after dosing, and liver specimens (approximately 200 mg each) were collected shortly after sacrifice as described elsewhere (Ji et al., 2018). Liver tissues were weighed and homogenized with ice-cold normal saline solution containing 0.1% formic acid (20%, w/v) to keep COG stable. All plasma and liver samples were separated after centrifugation at 4000 rpm for 10 minutes, and the supernatants were stored at −80°C until analysis.
Measurements of the Mouse mRNA Expression Levels of the Efflux Transporters of Interest.
Liver and intestinal specimens (approximately 200 mg each) were collected shortly after sacrifice as described elsewhere (Ji et al., 2018) and were stored at −80°C until analysis. To determine whether deletion of ATP-binding cassette, subfamily C (Abcc), member 3 gene could significantly modulate the expression levels of the genes that encode the efflux transporters of interest, including Abcc1, Abcc2, and Abcg2, we measured the mRNA expression levels in the intestine and liver of the Abcc3 KO versus WT mice using quantitative reverse-transcription polymerase chain reaction (PCR). In brief, the extraction of total RNA and subsequent synthesis of cDNA were described elsewhere (Tai et al., 2016) according to the protocols from the manufacturers. The primer sequences for the genes tested and the internal reference gene encoding glyceraldehyde-3-phosphate dehydrogenase (Gapdh) are shown in Supplemental Table 1. PCR amplification and melting analysis were conducted using the ABI 7500 real-time PCR apparatus (Applied Biosystems, Carlsbad, CA), as detailed elsewhere (Tai et al., 2016). The mRNA expression level was presented by a cycle threshold (Ct) value (defined as the number of PCR cycles that have crossed an arbitrarily chosen signal threshold in the log phase of the amplification curve), and relative expression of a target gene was expressed as a fold change, which was calculated using the following equation: fold change = 2−∆∆Ct, where ∆Ct = Ct(target) − Ct(gapdh).
The Active Uptake of COG into the Inverted (Inside-Out) Human MRP3 Membrane Vesicles.
The vesicular uptake studies were performed using the rapid filtration technique, as described previously (Ji et al., 2018). The “reaction buffer” (Solvo kit) consisted of 50 mM 3-(N-morpholino) propanesulfonic acid-Tris, 70 mM KCl, and 7.5 mM MgCl2, 10 mM ATP (or AMP), COG (1 or 10 μM), E217βG (4 μM, as the positive control), and recombinant human MRP3 membrane vesicles (inside-out). Human MRP3 vesicle suspensions were loaded onto the 96-well flat bottom of the tissue culture plates. After preincubation at 37°C for 5 minute, the reactions were initiated by the addition of the compound tested in the presence of ATP or AMP at 37°C for 5 minutes and stopped by the addition of 200 μl of ice-cold “washing mix” (Solvo kit) containing 400 mM 3-(N-morpholino)propanesulfonic acid-Tris and 700 mM KCl. Samples were then transferred to the 96-well glass-fiber filter plate and filtered using a Multiscreen HTS vacuum manifold (Millipore, Billerica, MA). The filters were washed with an ice-cold washing mix five times. The MRP3 vesicles on the filters were extracted by 80% acetonitrile at room temperature, and the supernatants were collected after centrifugation at 2000 rpm for 2 minutes. The ATP-dependent uptake of the testing compound into recombinant human MRP3 membrane vesicles was measured by the liquid chromatography with tandem mass spectrometry (LC-MS/MS) method (see below for details). The uptake assays were performed in triplicate, and all data are presented as means ± S.D.
The amount of CUR or COG transported by the inverted membrane vesicles of MRP3 was calculated using the following formula:

where “A” refers to the transported amount of a testing compound in the presence of ATP, and “B” refers to the transported amount of the same testing compound in the presence of AMP, both of which were normalized to the incubation time (minutes) and the amount of total vesicular protein (milligrams).
In this study, the measured uptake ratio of E217βG was greater than 2-fold, which was required as the positive control, indicating that the incubation system used worked well and that data generated were reliable and reproducible. Similarly, for the compound to be tested, if its uptake ratio in the presence of ATP was 2-fold greater than that in the presence of AMP, that compound would be considered a substrate of human MRP3.
Sample Preparation.
For the preparation of plasma and liver samples, an aliquot of 10-μl sample was spiked with 100 μl of acetonitrile (containing 0.1% formic acid and 30 ng/ml hesperetin as the IS). For the preparation of MRP3 vesicles, an aliquot of 30-μl sample was spiked with 60 μl of acetonitrile (containing 30 ng/ml hesperetin as the IS). All samples were vortex mixed for 3 minutes and subsequently centrifuged at 4000 rpm at 4°C for 10 minutes. The resulting supernatants were centrifuged at 14,000 rpm for 20 minutes at 4°C. Finally, an aliquot of 5 μl was injected into the LC-MS/MS system for quantification.
LC-MS/MS Conditions for the Quantification of CUR, COG, and E217βG.
The amount of CUR, COG, and E217βG was quantified by an LC-MS/MS system, which was equipped with a Shimadzu LC–30A Series system (Shimadzu, Kyoto, Japan) and a Sciex API 4500 tandem mass spectrometer (Applied Biosystems, Foster City, CA). Chromatographic separation was achieved at 40°C on a Poroshell 120 SB-C18 column (100 × 2.1 mm, 2.7 μm). The eluant was 0.1% formic acid in water (solvent A) and 0.1% formic acid in acetonitrile (solvent B). The gradient elution was set as follows: solvent B was increased from 20% to 90% at 0.01–2.30 minutes, maintained at 90% at 2.31–4.50 minutes, and then declined to 20% at 4.51 minutes and was maintained at 20% until the end of elution at 5.50 minutes at a flow rate of 0.3 ml/min. The temperature of the autosampler was maintained at 4°C, and the injection volume was 5 μl. The MS was operated using an electron spray ionization source with multiple reaction monitoring mode in negative ion mode. The MS parameters were as follows: ion spray voltage at −4500 V; ion source temperature 400°C; and curtain gas, collision gas, and ion source gas-1 and -2 set at 15, 8, 50, and 55 Psi, respectively. The compound-selective MS parameters were optimized and are summarized in Supplemental Table 2.
Statistical Analysis.
Data are presented as means ± S.D. Pharmacokinetics parameters of COG were analyzed for each mouse by Phoenix WinNonlin version 6.1 (Pharsight Co., Ltd., Mountain View, CA) using a noncompartmental analysis model. Statistical analyses were performed using unpaired Student’s t test for the group comparison of a single variable. A two-tailed P value of less than 0.05 is considered statistically significant.
Results
Impaired Efflux Transport of COG in Mrp3 KO Mice.
Because MRP3 preferentially transports its substrates (e.g., glucuronide conjugates) from enterocytes and hepatocytes into the blood, it was predicted that plasma COG concentrations would be significantly lower in Mrp3 KO mice than in WT mice after oral dosing of CUR. Illustrated in Fig. 2 are the mean plasma COG concentration-time curves in Mrp3 KO versus WT mice after receiving an oral dose of 200 mg/kg CUR, with the pharmacokinetics parameters summarized in Table 1. As expected, the maximum plasma COG concentrations (Cmax) were 32-fold lower in KO mice than in WT mice (P < 0.001), and area under the plasma drug concentration-time curve (AUC) to last time point measured values of COG were approximately 25-fold lower in KO mice than in WT mice, but elimination half-life (t1/2) was significantly prolonged in KO mice relative to WT mice. However, CUR pharmacokinetics was not analyzed because of low levels and high variation of unconjugated CUR in mouse plasma samples. These data suggest that systemic exposure of COG in plasma is significantly decreased in Mrp3 KO mice in comparison with WT mice, demonstrating that the efflux transport of COG formed in hepatocytes is almost abrogated because of knockout of the Abcc3 gene in mice.
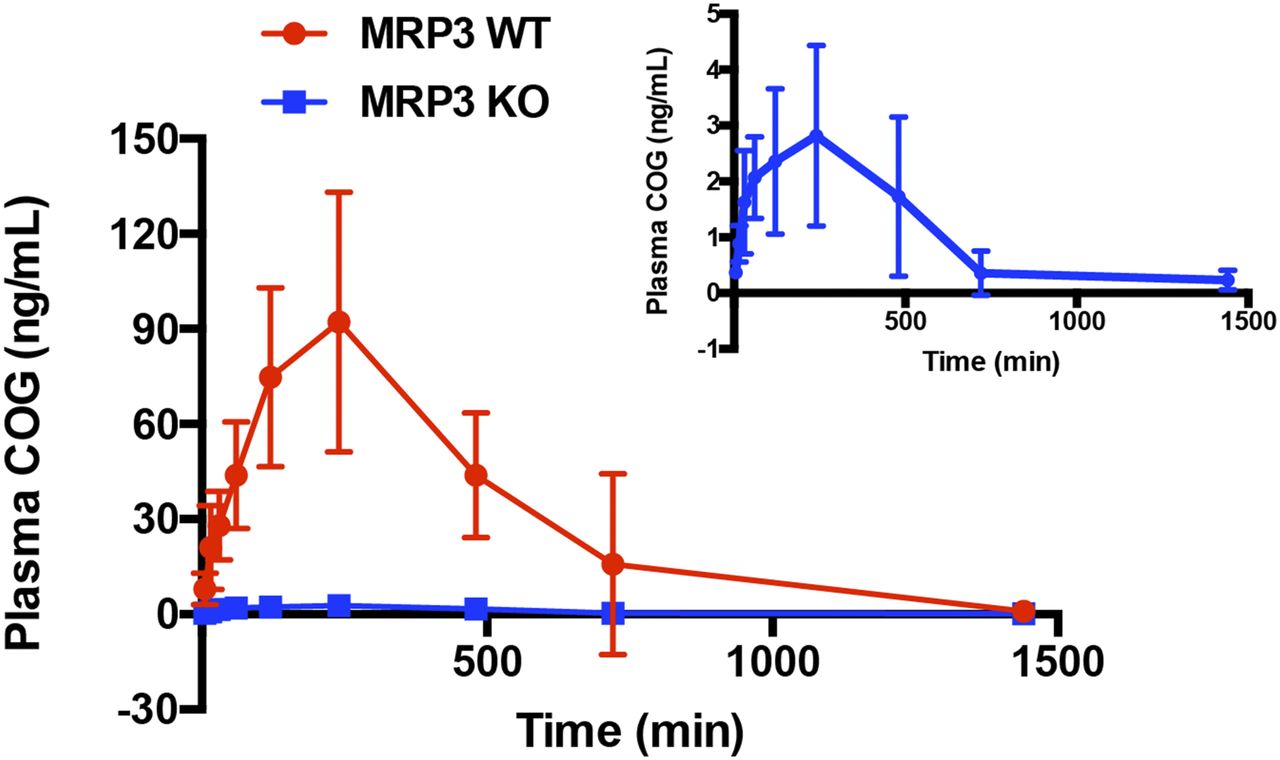
Mean plasma COG concentration-time curves in Mrp3 KO and WT mice after oral administration of 200 mg/kg of curcumin. Data are presented as means ± S.D.; n = 8 each.
The pharmacokinetics parameters of COG in Mrp3 WT and KO mice after oral administration of 200 mg/kg of CUR
Data are presented as means ± S.D. (n = 8 each). Unpaired Student’s t test.
Mrp3-Mediated Efflux Transport of COG Identified In Vivo and In Vitro.
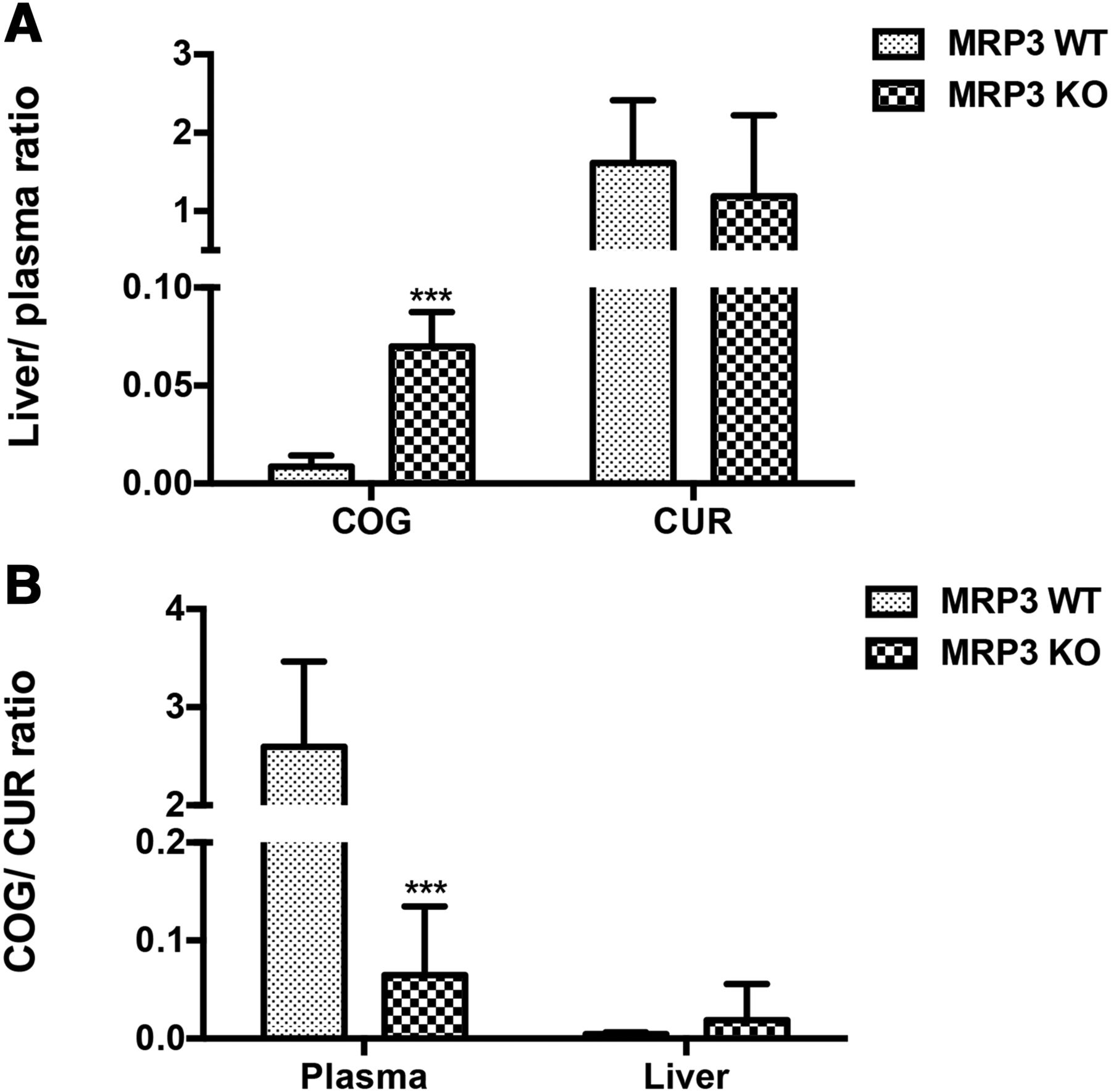
Considering that the hepatocyte Mrp3 transports its substrates from hepatocytes into the blood, the liver-to-plasma ratio of mice is generally recognized as the gold standard for determining whether a compound could be the substrate of Mrp3 as described elsewhere (Ji et al., 2018). As shown in Fig. 3A, the liver-to-plasma ratios of COG and those of CUR were determined in the presence or absence of Mrp3 in mice. Results indicated that the liver-to-plasma ratios of COG were 8-fold greater in Mrp3 KO mice than in WT mice, suggesting that COG is a substrate of Mrp3. As shown in Fig. 3A, there were no marked differences in the liver-to-plasma ratios of CUR between Mrp3 KO and WT mice, indicating that CUR is not a substrate of Mrp3. On the other hand, the COG-to-CUR ratios in plasma of KO mice were significantly lower compared with those of WT mice (Fig. 3B), which was the opposite of their changes in the liver, further suggesting that COG is a substrate of Mrp3.

The liver-to-plasma ratios of COG and CUR (A) and the COG-to-CUR ratio in plasma and liver (B) between Mrp3 KO and WT mice. Data are presented as means ± S.D.; n = 8 each. ***P < 0.001 vs. WT mice. Unpaired Student’s t test.
To confirm that COG is also an MRP3 substrate in humans, inverted membrane vesicles that overexpress human MRP3 were used to evaluate the ATP-dependent uptake of COG. As a well-characterized MRP3 substrate, E217βG was used in parallel as the positive control to further validate the incubation system used and data derived in this study. As shown in Fig. 4A, the ATP-dependent uptake of E217βG was 4.6-fold greater in the presence of ATP than in the presence of AMP, indicating that the vesicular transport assay used was reliable. After incubation with 1 or 10 μM COG in the MRP3-overexpressing vesicles, the uptake activity of MRP3 was 5.0- and 3.1-fold higher in the presence of ATP than in the presence of AMP, respectively. As shown in Fig. 4B, the uptake ratio of COG was greater than 2-fold when added at a concentration of 1 or 10 μM, as was E217βG. These data confirmed that COG is a substrate of human MRP3 in vitro.

The ATP-dependent uptake of COG into the inverted recombinant human MRP3 membrane vesicles. (A) Uptake activity of COG and E217βG in the presence of ATP or AMP. (B) Uptake ratio of COG and E217βG in the presence of ATP vs. AMP. Transport was measured after 5 minutes of uptake at 37°C in the presence of 1 or 10 μM COG. E217βG (4 μM) was used as a positive substrate for MRP3. Data are presented as means ± S.D.; n = 3 each. *P < 0.05; ** P < 0.01; and ***P < 0.001 vs. AMP group. Unpaired Student’s t test.
Similar Levels of the Abcc2 and Abcg2 mRNA Expression between Mrp3 KO and WT Mice.
To further investigate the possible effects of the Abcc3 gene deletion on other relevant transporters in mice, the mRNA expression levels of the four genes of interest (Abcc1, Abcc2, Abcc3, and Abcg2) were measured with normalization against the mRNA expression level of the reference gene Gapdh and subsequently were compared between the Abcc3 KO and WT mice. As shown in Fig. 5, except for the lack of Abcc3 mRNA expression in the Abcc3 KO mice as anticipated, there were no marked differences in the mRNA expression levels of the other genes tested between the Abcc3 KO and WT mice, suggesting that Abcc3 gene disruption seems not to modulate the expression of the genes evaluated.

Differences in the mRNA expression levels of Abcc1, Abcc2, Abcc3, and Abcg2 in the liver (A) and intestine (B) of Abcc3/Mrp3 KO and WT mice. Data are presented as means ± S.D.; n = 6 each. ***P < 0.001 vs. Mrp3 WT group. Unpaired Student’s t test.
Discussion
The most significant finding of this study, which used an Mrp3 KO mouse model and recombinant human MRP3 inverted membrane vesicles, is that MRP3 is responsible exclusively for the efflux transport of COG, revealing a critical role of MRP3 in the efflux transport of COG in the body. These novel findings would be used to better explain herb-drug interactions in patients when they are concomitantly taking CUR and any other drugs known to be MRP3 inhibitors and/or substrates or drugs whose main metabolic pathway is glucuronidation. In terms of the widespread use of CUR in Southeast Asians, such herb-drug interactions seem to be very important.
COG, generated in enterocytes and hepatocytes by UGTs (Lu et al., 2017), is transported to the blood, in turn, by the basolateral membrane transporter MRP3 of enterocytes and the sinusoidal plasma membrane transporter MRP3 of hepatocytes, consequently leading to a dramatic increase in its systemic exposure. As summarized in Table 1, compared with WT mice, Mrp3 KO mice exhibited significantly declined systemic exposure of COG (reflective of decreased AUC and Cmax) and an extended elimination process (increased t1/2), consistent with expected results. As shown in Fig. 2, plasma COG concentrations were extremely low (close to the lower limit of quantification) in Mrp3 KO mice, indicating that the efflux transport of COG formed inside cells is mediated exclusively by Mrp3. In theory, for Mrp3 KO mice, the liver-to-plasma ratio of COG would be elevated significantly, and the plasma COG-to-CUR ratio would be decreased. In this study, there was complete consistency between the observed and expected results.
Evidence has demonstrated that Mrp2 is involved in the biliary excretion of COG (Lee et al., 2012). It was speculated that blunted sinusoidal transport of COG to the blood in hepatocytes of Mrp3 KO mice could be compensated with markedly increased biliary elimination, as a result of enhanced expression of other efflux transporters (such as Mrp2 and Bcrp) that are expressed on the basolateral membrane of hepatocytes. It is not the case. In this study, the mRNA expression levels of several relevant efflux transporters were measured with the liver and intestinal samples for comparison between the two groups (Mrp3 KO vs. WT mice). As shown in Fig. 5, there were no significant differences in the mRNA expression levels of the genes Abcc2 (encoding Mrp2) and Abcg2 (encoding Bcrp) between the two groups, consistent with earlier observations that hepatic Mrp2 expression levels are not different between Mrp3 KO and WT mice (Zelcer et al., 2006), and that the biliary excretion of diclofenac acyl glucuronide (a known Mrp2 substrate) was similar between the Mrp3 KO and WT mice (Seitz et al., 1998). Similar to the case of diclofenac acyl glucuronide, Mrp2-mediated efflux transport of COG (also an Mrp2 substrate) (Lee et al., 2012) from hepatocytes to the bile was not enhanced in Mrp3 KO mice relative to WT mice. The above evidence strongly suggests that Abcc3 gene deletion seems not to affect the expression level and transport activity of Mrp2.
Although CUR is an inhibitor and substrate of BCRP (Berginc et al., 2012; Karibe et al., 2018), no evidence is currently available showing that COG is a substrate of BCRP. In our preliminary study, there were no marked differences in the Abcg2 mRNA expression levels of hepatocytes and enterocytes between the Mrp3 KO and WT mice (see Fig. 5), implying the lack of anticipated increases in the Bcrp-mediated biliary excretion of COG in Mrp3 KO mice.
In the gut mucosa and epithelial cells, ingested CUR is converted to COG by the intestinal UGTs (i.e., UGT1A7, UGT1A8, and UGT1A10) (Lu et al., 2017), which is transported, in turn, to the blood via Mrp3 expressed on the basolateral membrane of enterocytes or back to the gut lumen via the efflux transporters (such as Mrp2 and Bcrp) expressed on the apical side of enterocytes, where the enterohepatic circulation of COG would occur (Bangphumi et al., 2016). Furthermore, COG formed in hepatocytes is transported to the blood via Mrp3 expressed on the sinusoidal side of hepatocytes and to the bile via Mrp2 and Bcrp on the apical side of hepatocytes. Drug metabolism and transport are simultaneous in the body, and their entire processes are overlapping and dynamic over time. The interplay between one transporter and another and between drug metabolism and drug transport in the body is thus more complicated than we thought previously.
In view of extremely low systemic bioavailability of CUR, the major site for metabolic interactions of CUR with other coadministered drugs is most likely in the intestine. As an inhibitor of MRP3 and P-glycoprotein, CUR can increase intracellular accumulation of cancer drug phosphor-sulindac in cancer cell lines A549 and SW480 through inhibition of the affected efflux transporters if the two drugs are used concomitantly (Cheng et al., 2013). Diclofenac acyl glucuronide is also a substrate of MRP3 (Scialis et al., 2019). In addition, our recent work indicated that Mrp3 can transport clopidogrel acyl glucuronide from hepatocytes into the blood in mice (Ji et al., 2018). With concurrent use of CUR and any other known MRP3 substrates or inhibitors, such as phosphor-sulindac, diclofenac (acyl glucuronide), or clopidogrel (acyl glucuronide), clinically important herb-drug interactions would occur. Of all the glucuronide conjugates, many are the MRP3 substrates, such as COG, which is identified in this work. Not limited to these cases, oral intake of CUR would significantly decrease the systemic bioavailability of CYP3A4 substrate drugs through activation of CYP3A4 by COG (Hsieh et al., 2014). Therefore, metabolic interactions of CUR with other drugs seem to be more complex and clinically important than we thought.
In summary, this study reveals that COG is transported predominantly by the efflux transporter Mrp3 in mice and is confirmed by the inverted recombinant human MRP3 membrane vesicle technique. These results suggest that herb-drug interactions might occur in patients concomitantly taking CUR and either an MRP3 substrate/inhibitor or a drug that undergoes the glucuronidation pathway or whose metabolism is catalyzed by CYP3A4 predominantly. These new findings would help enrich and update our knowledge of drug transporters and drug transport for better understanding of the interplay between drug transport and drug metabolism in the body. In view of the widespread use of CUR globally, such herb-drug interactions are of clinical importance.
Acknowledgments
The authors thank Dr. Piet Borst, Dr. Koen van de Wetering, Sin-Ming Sit (office manager for research), Frank Hoorn (office manager for technology transfer), and Carla Rijnders, the Netherlands Cancer Institute, Amsterdam, the Netherlands, for generously providing Abcc3/Mrp3 KO mice and expert assistance, and Mr. Michael B. Xie, Martin Luther King Jr. Magnet High School, Nashville, TN, USA, for his critical proof reading of the manuscript.
Authorship Contributions
Participated in research design: Jia, Xie.
Conducted experiments: Jia, Zhu, Zhou, Ji, Tai.
Performed data analysis: Jia, Xie.
Wrote or contributed to the writing of the manuscript: Jia, Xie.
Footnotes
- Received August 27, 2019.
- Accepted December 18, 2019.
This work was supported in part by the National Natural Science Foundation of China [Grant 81473286 to H.-G.X.] and Nanjing First Hospital [Grant 31010300010339 to H.-G.X.]. H.-G.X. is the recipient of the Distinguished Medical Experts of the Province of Jiangsu, China.
Abbreviations
- ABCC
- ATP-binding cassette, subfamily C
- AUC
- area under the plasma drug concentration-time curve
- BCRP
- breast cancer resistance protein
- Cmax
- the maximum plasma drug concentration (observed)
- COG
- curcumin-O-glucuronide
- CUR
- curcumin
- E217βG
- estradiol-17β-D-glucuronide
- Gapdh
- glyceraldehyde-3-phosphate dehydrogenase
- IS
- internal standard
- KO
- knockout
- LC-MS/MS
- liquid chromatography with tandem mass spectrometry
- MRP
- multidrug resistance–associated protein
- PCR
- polymerase chain reaction
- t1/2
- half-life
- UGT
- UDP-glucuronosyltransferase
- WT
- wild-type
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}