


Abstract
Human arylacetamide deacetylase (AADAC) plays a role in the detoxification or activation of drugs and is sometimes involved in the incidence of toxicity by catalyzing hydrolysis reactions. AADAC prefers compounds with relatively small acyl groups, such as acetyl groups. Eslicarbazepine acetate, an antiepileptic drug, is a prodrug rapidly hydrolyzed to eslicarbazepine. We sought to clarify whether AADAC might be responsible for the hydrolysis of eslicarbazepine acetate. Eslicarbazepine acetate was efficiently hydrolyzed by human intestinal and liver microsomes and recombinant human AADAC. The hydrolase activities in human intestinal and liver microsomes were inhibited by epigallocatechin gallate, a specific inhibitor of AADAC, by 82% and 88% of the control, respectively. The hydrolase activities in liver microsomes from 25 human livers were significantly correlated (r = 0.87, P < 0.001) with AADAC protein levels, suggesting that the enzyme AADAC is responsible for the hydrolysis of eslicarbazepine acetate. The effects of genetic polymorphisms of AADAC on eslicarbazepine acetate hydrolysis were examined by using the constructed recombinant AADAC variants with T74A, V172I, R248S, V281I, N366K, or X400Q. AADAC variants with R248S or X400Q showed lower activity than wild type (5% or 21%, respectively), whereas those with V172I showed higher activity than wild type (174%). Similar tendencies were observed in the other four substrates of AADAC; that is, p-nitrophenyl acetate, ketoconazole, phenacetin, and rifampicin. Collectively, we found that eslicarbazepine acetate is specifically and efficiently hydrolyzed by human AADAC, and several AADAC polymorphic alleles would be a factor affecting the enzyme activity and drug response.
SIGNIFICANCE STATEMENT This is the first study to clarify that arylacetamide deacetylase (AADAC) is responsible for the activation of eslicarbazepine acetate, an antiepileptic prodrug, to eslicarbazepine, an active form, in the human liver and intestines. In addition, we found that several AADAC polymorphic alleles would be a factor affecting the enzyme activity and drug response.
Introduction
Drug-metabolizing enzymes play roles in the detoxification or activation of drugs and sometimes cause drug-induced toxicity by forming reactive metabolites. Among drug-metabolizing enzymes, cytochrome P450 (P450) is responsible for the metabolism of over 50% of clinical drugs with the highest contribution. Recently, a better understanding of roles of enzymes other than P450, non-P450 enzymes, is required, because their contribution for the metabolism of clinical drugs has gradually increased (Cerny, 2016). Hydrolases catalyze the hydrolysis of ester, amide, and thioester bonds and are largely involved in the activation of prodrugs, detoxification, and sometimes toxicity (Fukami and Yokoi, 2012; Cerny, 2016). Among hydrolases, carboxylesterase (CES) is the best-characterized hydrolase family, and two isoforms, CES1 and CES2, mainly catalyze the hydrolysis reaction of drugs (Fukami and Yokoi, 2012). CES1 is predominantly expressed in the liver, whereas CES2 is expressed in the small intestine as well as liver. In addition to CES, our laboratory has demonstrated that arylacetamide deacetylase (AADAC) participates in the hydrolysis of clinical drugs including aromatic amides/esters such as flutamide (Watanabe et al., 2009), phenacetin (Watanabe et al., 2010), and prasugrel (Kurokawa et al., 2016), and aliphatic amides/esters such as rifamycins (rifampicin, rifabutin, and rifapentine) (Nakajima et al., 2011), and ketoconazole (Fukami et al., 2016). AADAC is mainly expressed in the human liver and intestines (Watanabe et al., 2009). We have reported that human hydrolase(s) involved in the hydrolysis of the target compounds can be roughly predicted based on the size of the acyl or alcohol/amine group in the compounds (Fukami et al., 2015). CES1 prefers compounds with relatively large acyl groups, whereas CES2 and AADAC prefer compounds with relatively small acyl groups. Especially, AADAC prefers compounds with acetyl group.
Carbamazepine, a widely used antiepileptic drug that was developed in the 1960s, sometimes causes adverse reactions, including central nervous system disorders. It has been suggested that an intermediate metabolite, carbamazepine 10,11-epoxide (Fig. 1), could be the cause of the adverse reactions of carbamazepine (Fricke-Galindo et al., 2018). To reduce carbamazepine-induced adverse reactions, oxcarbazepine was developed (Koch and Polman, 2009). Oxcarbazepine is a prodrug that is converted to eslicarbazepine (S-licarbazepine) and R-licarbazepine via a reduction reaction in the human liver, but only the former metabolite has pharmacological actions (Malátková et al., 2014; Potschka et al., 2014). Later, eslicarbazepine acetate was developed with the aim of efficient and specific formation of eslicarbazepine (Nunes et al., 2013) (Fig. 1). Eslicarbazepine acetate, of which at least 90% of the dose is absorbed from the intestines regardless of food intake, is quickly hydrolyzed to eslicarbazepine by first-pass metabolism (Maia et al., 2005; Almeida et al., 2008). According to the known substrate preference of drug-metabolizing hydrolases (Fukami et al., 2015), it is conceivable that eslicarbazepine acetate might be hydrolyzed by AADAC because it has an acetic acid ester.

Metabolic pathways of carbamazepine, oxcarbazepine, and eslicarbazepine acetate in humans.
In this study, we sought to identify the enzyme(s) responsible for the hydrolysis of eslicarbazepine acetate. For the AADAC gene, many single nucleotide polymorphisms (SNPs) are registered on dbSNP (https://www.ncbi.nlm.nih.gov/snp/). However, the significance of most SNPs, except g.14008T>C (X400Q), which has been found to substantially decrease enzyme activity (Shimizu et al., 2012), remains to be determined. If AADAC is the enzyme responsible for eslicarbazepine acetate hydrolysis, it is possible that SNPs in the AADAC gene affect the drug response. We examined the effects of six kinds of SNPs causing amino acid substitutions on eslicarbazepine acetate hydrolysis.
Materials and Methods
Chemicals and Reagents.
Eslicarbazepine acetate and eslicarbazepine were purchased from Tokyo Chemical Industry (Tokyo, Japan) and Tocris Bioscience (Bristol, UK), respectively. Epigallocatechin gallate (EGCg), diisopropyl fluorophosphate (DFP), flutamide, p-nitrophenol, 4-nitro-3-(trifluoromethyl)phenylamine, phenacetin, phenylmethylsulfonyl fluoride (PMSF), rifampicin, and vinblastine were purchased from FUJIFILM Wako Pure (Osaka, Japan). Bis (p-nitrophenyl) phosphate (BNPP), p-nitrophenyl acetate (PNPA), and p-phenetidine were purchased from Sigma-Aldrich (St. Louis, MO). Ketoconazole, N-deacetyl ketoconazole, and telmisartan were purchased from LKT Laboratories (St. Paul, MN). 25-Deacetylrifampicin was purchased from Toronto Research Chemicals (Toronto, Canada). Human intestinal and liver microsomes (pooled HIMs, n = 7 and pooled HLMs, n = 50) and human liver cytosol (pooled HLC, n = 50) were purchased from Corning (Corning, NY). Human intestinal cytosol (pooled HIC, n = 6) was purchased from XenoTech (Lenexa, KS). A Bac-to-Bac Baculovirus Expression System, Sf-900 II SFM insect cell medium, and Spodoptera frugiperda Sf21 cells were purchased from Thermo Fisher Scientific (Waltham, MA). Taq DNA polymerase was purchased from Greiner (Tokyo, Japan). The pTargeT Mammalian Expression Vector was purchased from Promega (Madison, WI). Recombinant human AADAC, CES1, and CES2 expressed in baculovirus-infected insect cells were prepared previously (Fukami et al., 2010; Watanabe et al., 2010). All other reagents used in this study were of analytical or the highest quality grade commercially available.
Preparation of Microsomal Fractions from a Panel of 25 Individual Human Livers.
Human liver samples from 25 individual donors were obtained from the National Disease Research Interchange (Philadelphia, PA) through the Human and Animal Bridging Research Organization (Chiba, Japan). HLMs were prepared according to a previously reported method (Kobayashi et al., 2012). The use of liver samples was approved by the Ethics Committee of Kanazawa University (Kanazawa, Japan). The donor information is shown in Table 1.
Characteristics of 25 donors of liver samples used in this study
Eslicarbazepine Acetate Hydrolase Activity.
Eslicarbazepine acetate hydrolase activity was determined as follows: a typical incubation mixture (final volume, 0.2 ml) contained 100 mM Tris-HCl buffer (pH 7.4) and enzyme sources (pooled HIMs, HIC, HLMs, HLC, and homogenates of Sf21 cells expressing human AADAC, CES1, or CES2: 0.1 mg/ml). After a 2-minute preincubation at 37°C, eslicarbazepine acetate dissolved in DMSO was added to the incubation mixture so that the final concentration of DMSO was 1.0%. The concentration of eslicarbazepine acetate was set to 1 mM. After a 5-minute incubation at 37°C, the reaction was terminated by the addition of 100 µl of ice-cold methanol. In preliminary experiments, we confirmed that the linearities of eslicarbazepine formation in terms of the protein concentration and incubation time were up to 0.2 mg/ml and 10 minutes, respectively. After the removal of the protein by centrifugation at 20,380g for 5 minutes, a 60 μl aliquot of the supernatant was subjected to high-performance liquid chromatography (HPLC). The HPLC equipment consisted of an L-2100 pump (Hitachi, Tokyo, Japan), an L-2200 autosampler (Hitachi), an L-2400 UV detector (Hitachi), and a D-2500 HPLC Chromato-Integrator (Hitachi) equipped with Wakopak eco-ODS (4.6 × 150 mm internal diameter, 5 μm; Wako, Tokyo, Japan). The eluent was monitored at 235 nm. The mobile phase was 35% methanol, and the flow rate was 1.0 ml/min. The column temperature was 35°C. The quantification of the metabolites was performed by comparing the HPLC peak heights with those of an authentic standard.
Kinetic Analysis.
Kinetic analysis was performed using pooled HIMs, HLMs, and recombinant AADAC with substrate concentrations ranging from 0.1 to 5 mM. The kinetic parameters were estimated from a curve fitted using a computer program designed for nonlinear regression analysis (KaleidaGraph; Synergy Software, Reading, PA).
Inhibition Analysis of Eslicarbazepine Acetate Hydrolase Activity.
To examine the responsibility of AADAC for eslicarbazepine acetate hydrolysis in humans, inhibition analyses were performed using pooled HIMs, HLMs, and recombinant AADAC. DFP and BNPP are serine hydrolase inhibitors (Heymann and Krisch, 1967; Yamaori et al., 2006). PMSF is an inhibitor for both CES enzymes, telmisartan is a specific inhibitor for CES2, and vinblastine is an inhibitor for CES2 and AADAC (Shimizu et al., 2014b). EGCg is a specific inhibitor for AADAC (K. Yasuda et al., manuscript in preparation). CaCl2 is an activator of paraoxonase. The concentrations of the inhibitors were set according to those previous reports as follows: 100 μM for DFP, BNPP, and PMSF; 5 μM for telmisartan; 50 μM for vinblastine and EGCg; and 1 mM for CaCl2. DFP, PMSF, telmisartan, and vinblastine were dissolved in DMSO such that the final concentration of DMSO in the incubation mixture was 2%. The other inhibitors were dissolved in distilled water. The experimental procedure and conditions were the same as above.
Immunoblot Analysis.
SDS-polyacrylamide gel electrophoresis and immunoblot analysis for human AADAC were performed according to our previous report (Watanabe et al., 2009). Human liver microsomes (30 μg) were separated on 10% polyacrylamide gels and electrotransferred onto polyvinylidene difluoride membranes (Immobilon-P; Millipore Corporation, Billerica, MA). The membrane was probed with monoclonal mouse anti-human AADAC antibody (Abnova, Taipei City, Taiwan) and the corresponding fluorescent dye–conjugated secondary antibody. The band densities were quantified with an Odyssey Infrared Imaging system (LI-COR Biosciences, Lincoln, NE). This analysis was performed in the linear range of band intensity with respect to the amount of protein.
Construction of Expression Plasmids for AADAC Variants.
The expression plasmids for four kinds of AADAC variants causing amino acid substitutions, c.310A>G (T74A), c.604G>A (V172I), c.834A>T (R248S), or c.1188C>G (N366K), were constructed according to our previous reports (Watanabe et al., 2009; Shimizu et al., 2012). The plasmids for wild type and V281I or Q400X variants were previously constructed (Shimizu et al., 2012). These six SNPs were focused based on the over 0.1% minor-allele frequencies in the dbSNP data base (Table 2). Each mutation was introduced into the pTargeT mammalian expression vector of AADAC wild type through site-directed mutagenesis with a Quick Change II XL site-directed mutagenesis kit with the primer sets (Table 3). Nucleotide sequences were confirmed with DNA sequence analysis. The AADAC cDNA in the pTargeT vector was then transferred into the pFastBac1 vector using appropriate restriction enzymes.
SNPs investigated in this study
All frequencies were referred to dbSNP.
Primers used to introduce mutations in the AADAC gene and for AADAC genotyping
The underlined letters indicate mutations.
Expression of Human AADAC Variants in Sf21 Cells.
The pFastBac1 vectors containing each AADAC cDNA were transformed into DH10Bac competent cells, followed by transposition of the inserts into bacmid DNA. A Bac-to-Bac Baculovirus Expression System was used to express human AADAC, as described in our previous report (Watanabe et al., 2010). The AADAC protein levels of the constructed expression systems were determined by immunoblotting.
Hydrolase Activities for PNPA, Ketoconazole, Phenacetin, and Rifampicin by AADAC Variants.
PNPA, ketoconazole, phenacetin, and rifampicin hydrolase activities were determined according to our previous reports (Watanabe et al., 2010; Nakajima et al., 2011; Shimizu et al., 2012; Fukami et al., 2016) with slight modifications. The concentrations of homogenates of Sf21 cells expressing AADAC wild type or variants were set as follows: PNPA hydrolase activity, 0.05 mg/ml; ketoconazole hydrolase activity, 1.0 mg/ml; phenacetin hydrolase activity, 0.4 mg/ml; and rifampicin hydrolase activity, 0.4 mg/ml. The concentrations of PNPA, ketoconazole, phenacetin, and rifampicin were set at 100, 20 µM, 3 mM, and 200 µM, respectively.
Genomic DNA.
Genomic DNA samples were extracted from human blood samples by using a Puregene DNA isolation kit (Gentra Systems, Inc., Minneapolis, MN). The blood samples were from 176 Japanese, 205 Korean, 209 Caucasian, and 218 African American healthy subjects who provided written informed consent. The use of genomic DNA samples was approved by the Ethics Committee of Kanazawa University, Soonchunhyang University Hospital (Chonan, Korea), and the Human Studies Committee of Washington University School of Medicine (St. Louis, MO).
Genotyping of AADAC Alleles.
The frequencies of AADAC variant alleles having g.10583G>A (V172I) or g.13452A>T (R248S) were determined using genomic DNA samples. In addition to these two SNPs, the frequency of a whole gene deletion type, which has been suggested to be associated with Tourette syndrome in Europeans (Bertelsen et al., 2016), was determined. For genotyping of the alleles having g.10583G>A (V172I) or g.13452A>T (R248S), polymerase chain reaction (PCR)–restriction fragment length polymorphism analyses were performed. The PCR mixture contained genomic DNA (100 ng), 1× PCR buffer [67 mM Tris-HCl, pH 8.8, 16.6 mM (NH4)2SO4, 0.45% Triton X-100, 0.02% gelatin], 1.5 mM MgCl2, 0.2 mM dNTPs, 0.4 μM each primer (Table 3), and 0.5 U of Taq polymerase in a final volume of 25 µl. After an initial denaturation at 94°C for 3 minutes, the amplification was performed with denaturation at 94°C for 25 second, annealing at 54°C for 25 second, and extension at 72°C for 50 second for 30 cycles, followed by a final extension at 72°C for 5 minutes. The PCR products were digested with EcoRI and HinfI for g.10583G>A and g.13452A>T, respectively. The restriction products underwent electrophoresis in a 3% agarose gel. In the genotyping of g.10583G>A, the wild type yielded a 125–base pair fragment, and the variant yielded 101– and 24–base pair fragments. In the genotyping of g.13452A>T, the wild type yielded 145– and 72–base pair fragments, and the variant yielded a 217–base pair fragment. For genotyping of the AADAC whole gene deletion type, allele-specific PCR was performed. The locations of the primer pairs used for genotyping of the gene deletion type are shown in Supplemental Fig. 1. The PCR conditions were the same as above. The 584– and 679–base pair fragments should be detected for the wild type and deletion type, respectively.
Statistical Analysis.
Statistical significance between two groups was determined by an unpaired, two-tailed Student’s t test, and statistical significance between multiple groups was determined by ANOVA followed by Tukey’s test using Instat 2 software (GraphPad Software, San Diego, CA). Correlation analysis was performed using the Spearman rank method. A value of P < 0.05 was considered statistically significant.
Results
Eslicarbazepine Acetate Hydrolase Activities by Various Enzyme Sources.
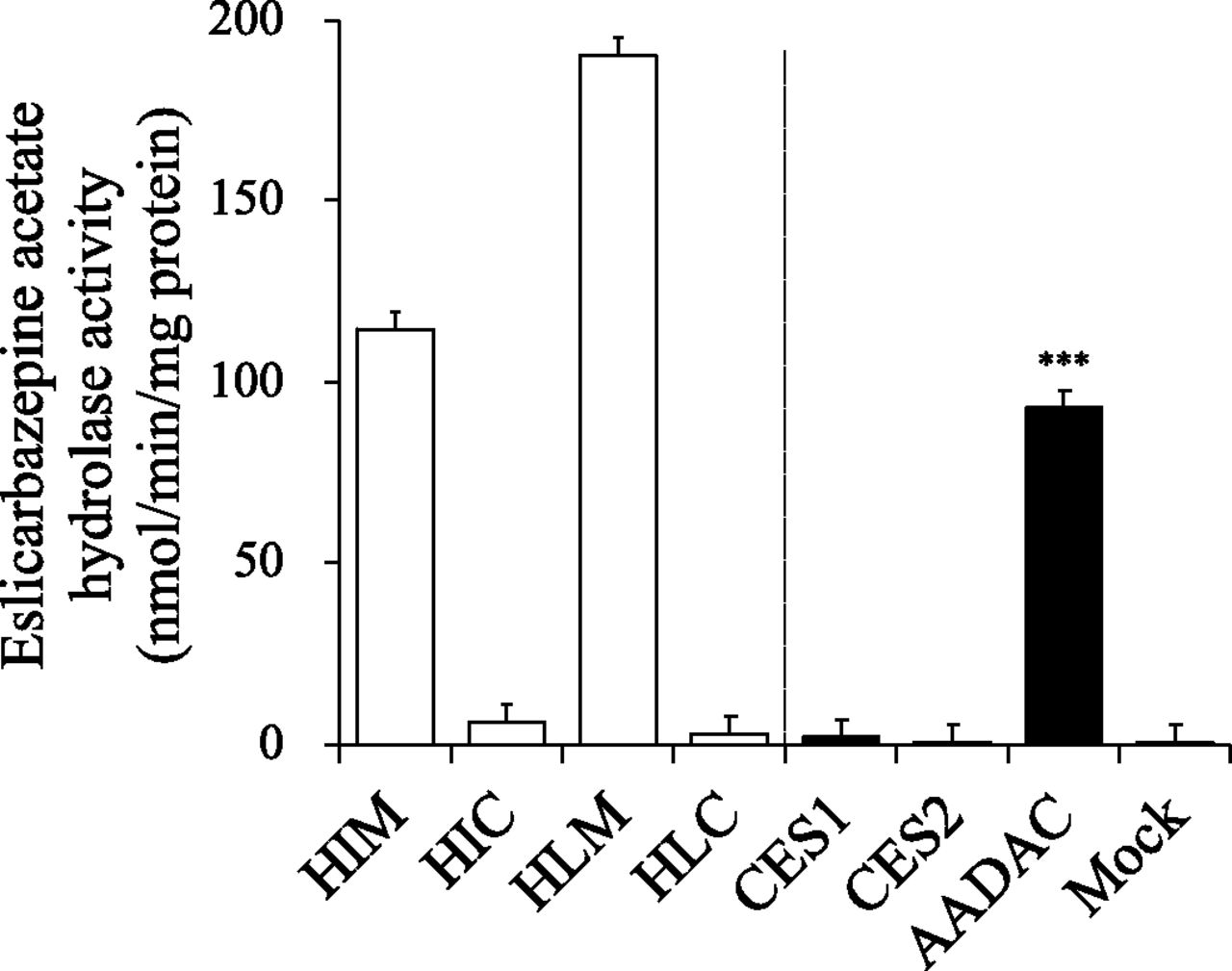
It is known that eslicarbazepine acetate is prominently hydrolyzed to eslicarbazepine by first-pass metabolism (Almeida et al., 2008). In this study, eslicarbazepine acetate hydrolase activities in pooled HIMs, HIC, HLMs, and HLC were evaluated at a substrate concentration of 1 mM, assuming the concentration in the intestinal lumen, although it was not expected to reach in the liver at this concentration (Fig. 2). The eslicarbazepine acetate hydrolase activities in HIMs and HLMs were 114.2 ± 4.2 nmol/min per milligram protein and 189.6 ± 13.4 nmol/min per milligram protein, respectively, whereas HIC and HLC showed marginal activity, suggesting that eslicarbazepine acetate is efficiently hydrolyzed in both the intestines and liver, and the responsible enzyme(s) is localized in the endoplasmic reticulum. Then, to examine the involvement of major drug-metabolizing hydrolases, CES1, CES2, or AADAC in eslicarbazepine acetate hydrolysis, the activities were measured by using their recombinant enzymes. As shown in Fig. 2, AADAC showed prominent eslicarbazepine acetate hydrolase activity (92.7 ± 4.2 nmol/min per milligram protein), whereas CES1 (1.7 ± 0.2 nmol/min per milligram protein) and CES2 (0.4 ± 0.3 nmol/min per milligram protein) showed marginal activities, which were close to the activity of the control (0.3 ± 0.2 nmol/min per milligram protein). Since AADAC is localized on the endoplasmic reticulum membrane in the intestine and liver, whereas CES1 (liver) and CES2 (intestine and liver) are localized in the cytosol as well as the endoplasmic reticulum (Xu et al., 2002; Frick et al., 2004; Tabata et al., 2004), it was suggested that the eslicarbazepine acetate hydrolase activities in HIMs and HLMs may be explained by AADAC.

Eslicarbazepine acetate hydrolase activities in HIMs, HIC, HLMs, HLC, and recombinant hydrolases. The substrate concentration was 1 mM. Each column represents the mean ± S.D. of triplicate determinations. ***P < 0.001, compared with the activity of the control using ANOVA followed by Tukey’s test.
Kinetic Analysis of Eslicarbazepine Acetate Hydrolase Activities in HIMs, HLMs, and Recombinant AADAC.
Kinetic analysis for eslicarbazepine acetate hydrolysis was performed using pooled HIMs, HLMs, and recombinant AADAC (Fig. 3). The kinetics in all cases were fitted to Michaelis-Menten kinetics, and the Eadie-Hofstee plots showed straight lines. The Km, Vmax, and intrinsic clearance values in HIMs were 1.16 ± 0.20 mM, 202.3 ± 7.2 nmol/min per milligram protein, and 0.17 ± 0.02 ml/min per milligram protein, respectively, and those in HLMs were 0.72 ± 0.08 mM, 283.7 ± 4.1 nmol/min per milligram protein, and 0.40 ± 0.05 ml/min per milligram protein, respectively (Table 4). Those by recombinant AADAC were 0.79 ± 0.11 mM, 157.5 ± 3.9 nmol/min per milligram protein, and 0.20 ± 0.02 ml/min per milligram protein, respectively (Table 4). The Km value by HIMs was slightly higher than that by recombinant AADAC, but the Km value by HLMs was close to that by recombinant AADAC. Thus, it was suggested that an enzyme catalyzing eslicarbazepine acetate hydrolysis in HIMs and HLMs would be AADAC.

Kinetic analysis of eslicarbazepine acetate hydrolase activities in HIMs (A), HLMs (B), and recombinant AADAC (C). The kinetic parameters were estimated from the fitted curve using the KaleidaGraph computer program designed for nonlinear regression analysis. Each data point represents the mean ± S.D. of triplicate determinations.
Kinetic parameters of eslicarbazepine acetate hydrolase activities
Inhibition Analysis of Eslicarbazepine Acetate Hydrolase Activities in HIMs, HLMs, and Recombinant AADAC.
To examine whether AADAC is a principal enzyme for eslicarbazepine acetate hydrolysis in human intestines and liver, inhibition analysis using representative inhibitors for eslicarbazepine acetate hydrolase activities in HIMs, HLMs, and recombinant AADAC was performed (Fig. 4). The eslicarbazepine acetate hydrolase activities in HIMs and HLMs were inhibited by DFP and BNPP, serine hydrolase inhibitors, but not by PMSF, a CES inhibitor, and telmisartan, a specific CES2 inhibitor. The activities were potently inhibited by vinblastine, an inhibitor of AADAC and CES2, and EGCg, a specific inhibitor of AADAC. The potent inhibition of AADAC by vinblastine and EGCg was confirmed by using recombinant AADAC. CaCl2, a paraoxonase activator, did not alter the activities. These results strengthened the finding that AADAC is responsible for eslicarbazepine acetate hydrolysis in the human intestine and liver.
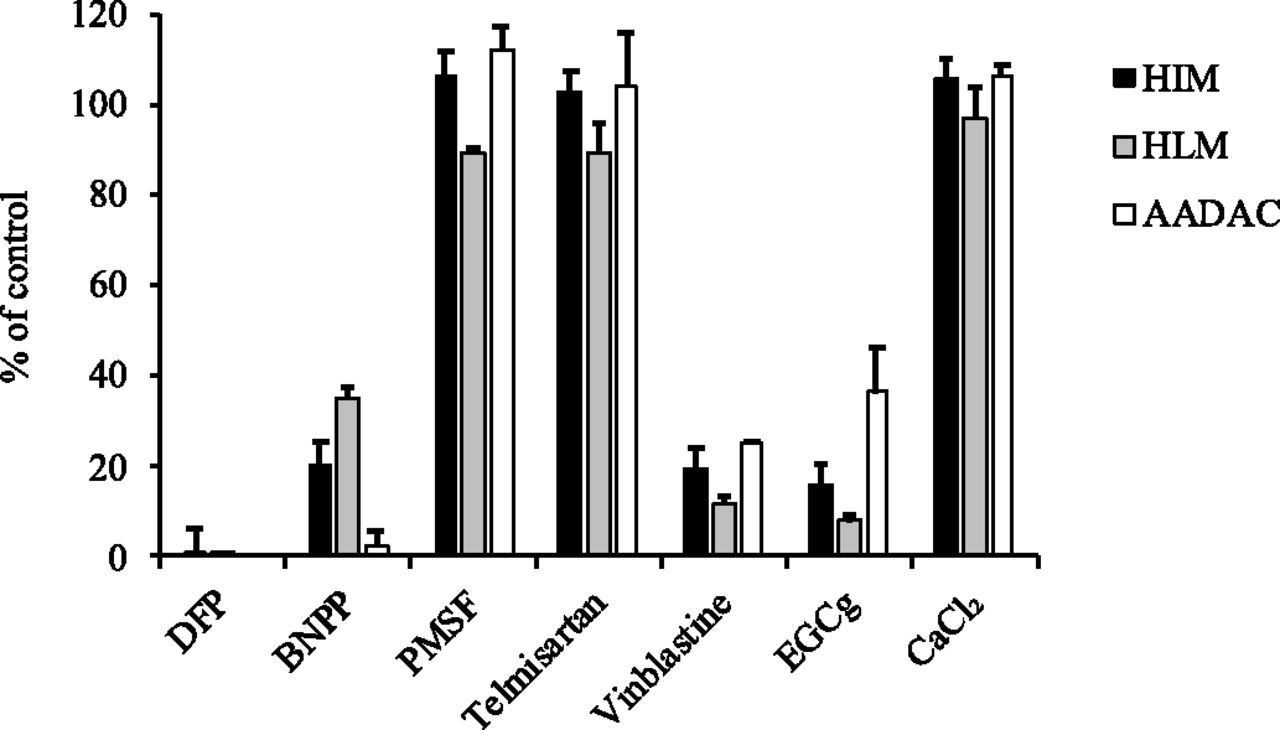
Effects of chemical inhibitors on eslicarbazepine acetate hydrolase activities in HIMs, HLMs, and recombinant AADAC. The substrate concentration was 1 mM. The concentrations of the inhibitors were as follows: DFP, BNPP, and PMSF, 100 µM; telmisartan, 5 µM; vinblastine and EGCg, 50 µM; and CaCl2, 1 mM. The control activities in HIMs, HLMs, and recombinant AADAC were 112.7 ± 7.6, 149.5 ± 2.5, and 92.9 ± 4.0 nmol/min per milligram protein, respectively. Each column represents the mean ± S.D. of triplicate determinations.
Correlation Analysis Between Eslicarbazepine Acetate Hydrolase Activities and AADAC Protein Levels in a Panel of 25 Human Liver Microsomes.
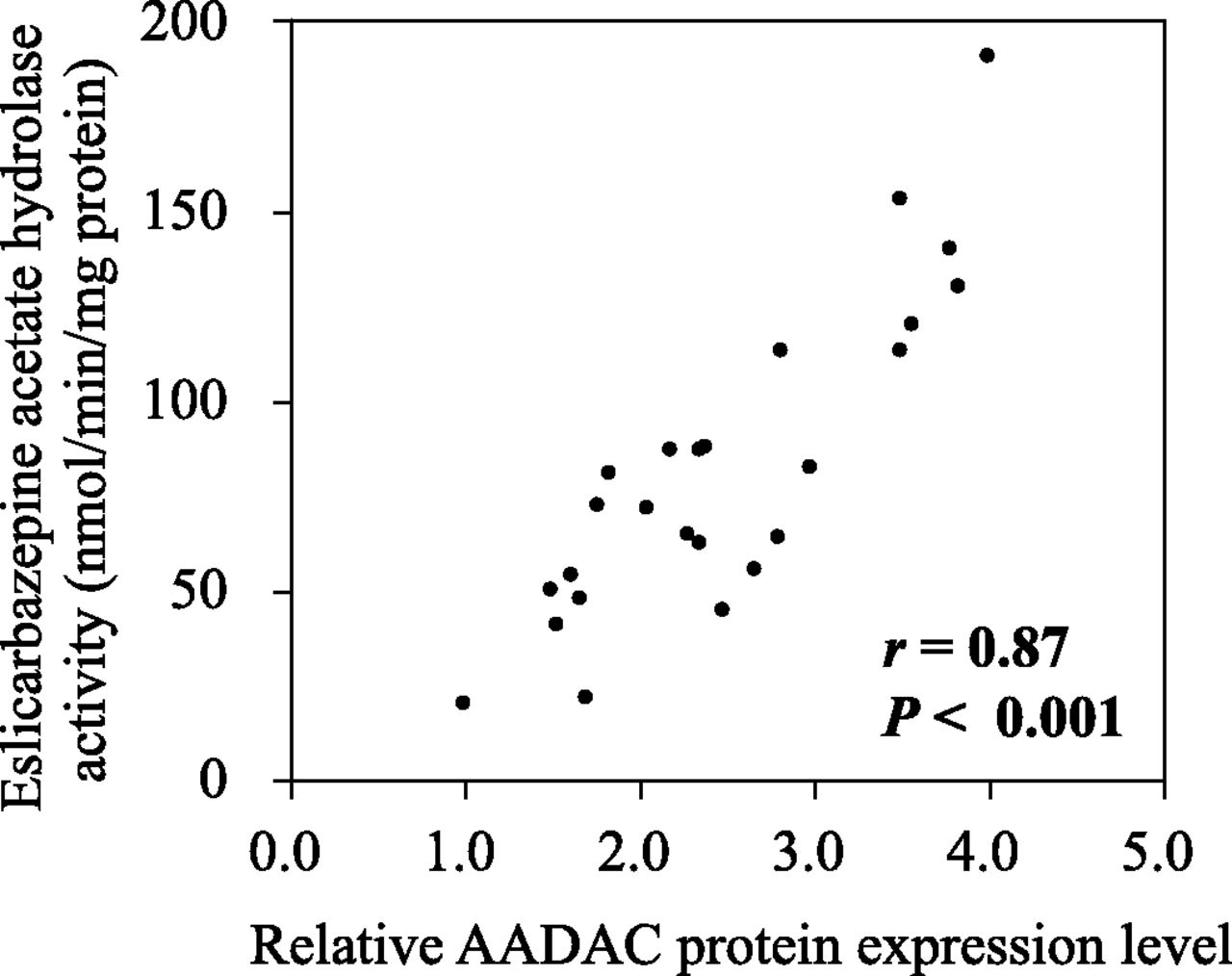
To further examine the responsibility of AADAC for eslicarbazepine acetate hydrolysis, correlation analysis between eslicarbazepine acetate hydrolase activities and AADAC protein levels was performed using a panel of liver microsomes from 25 individuals (Fig. 5). The eslicarbazepine acetate hydrolase activities ranged from 20.1 to 190.4 nmol/min per milligram protein, resulting in 9.5-fold interindividual variability. The AADAC protein levels ranged from 1.0 to 4.0, resulting in 4-fold interindividual variability. As shown in Fig. 5, the eslicarbazepine acetate hydrolase activities were significantly correlated with AADAC protein levels (r = 0.87, P < 0.001), supporting that AADAC is a principal enzyme for eslicarbazepine acetate hydrolysis.

Correlation analysis between AADAC protein levels and eslicarbazepine acetate hydrolase activities in a panel of 25 human liver microsomes. The AADAC protein levels determined by immunoblot analysis are represented as relative levels to the sample with the lowest expression level. The eslicarbazepine acetate hydrolase activity at a substrate concentration of 1 mM was determined by HPLC.
Effects of Genetic Polymorphisms of AADAC on Hydrolase Activities for Eslicarbazepine Acetate and Other AADAC Substrates.
To examine the effects of genetic polymorphisms of AADAC on eslicarbazepine acetate hydrolase activities, recombinant AADAC variants with T74A, V172I, R248S, and N366K were constructed. In addition, recombinant AADAC variants with V281I and X400Q, which had been constructed in our previous study (Shimizu et al., 2012), were also used (Fig. 6A). By immunoblotting, the expression levels of recombinant AADAC variants with T74A, V172I, R248S, V281I, N366K, and X400Q relative to that of wild type were calculated to be 6.71, 1.55, 2.25, 3.27, 2.87, and 5.54, respectively (data not shown). These values were used to normalize the hydrolase activities by recombinant AADAC variants. The eslicarbazepine acetate hydrolase activity by AADAC wild type was 101.1 ± 3.4 nmol/min per milligram protein. As shown in Fig. 6A, the eslicarbazepine acetate hydrolase activity by T74A, V281I or N366K variant was slightly but significantly higher than that by wild type (% of normalized activity by wild type: 106.4% ± 1.8%, 125.1% ± 6.4% or 121.7% ± 5.0%, respectively). The activity of the V172I variant was significantly higher than that of the wild type (173.9% ± 8.9%), whereas the activities of the R248S or X400Q variant were significantly lower than that of the wild type (5.4% ± 0.7% or 21.1% ± 1.2%, respectively).

Effects of AADAC variants on the hydrolase activities of eslicarbazepine acetate and other AADAC substrates. Eslicarbazepine acetate (A), PNPA (B), ketoconazole (C), phenacetin (D), and rifampicin hydrolase (E) activities by recombinant human AADAC wild type and variants were measured by HPLC or liquid chromatography–tandem mass spectrometry. *P < 0.05; **P < 0.01; ***P < 0.001, compared with the activity of wild type using an unpaired, two-tailed Student’s t test. Each column represents the mean ± S.D. of triplicate determinations. The eslicarbazepine acetate, PNPA, ketoconazole, phenacetin, and rifampicin hydrolase activities by wild type were 101.1 ± 3.4 nmol/min per milligram protein, 726.6 ± 21.8 nmol/min per milligram protein, 33.0 ± 2.3 pmol/min per milligram protein, 4.6 ± 0.0 nmol/min per milligram protein, and 63.9 ± 2.2 pmol/min per milligram protein, respectively.
To investigate whether similar trends were observed in the hydrolysis of other AADAC substrates, PNPA, ketoconazole, phenacetin, and rifampicin hydrolase activities were measured. As shown in Fig. 6, B–E, the T74A, V281I, and N366K variants had activities similar to those in wild types. The V172I variant commonly had higher activities, whereas the R248S and X400Q variants had lower activities than wild type. The trend was consistent with that observed in eslicarbazepine acetate hydrolysis. Thus, we found that the V172I variant has significantly increased activity, whereas R248S and X400Q variants have significantly decreased activities regardless of the substrates.
Allele Frequencies of V172I or R248S Variant as Well as a Whole Gene Deletion Type in 176 Japanese, 205 Koreans, 209 Caucasians, and 218 African Americans.
Previously, we reported that the allele frequencies of the X400Q variant in Caucasians and African Americans were 1.3% and 2.0%, respectively, whereas the variant was not found in Japanese and Koreans (Shimizu et al., 2012). In this study, the allele frequencies of g.10583G>A (V172I), g.13452A>T (R248S), and a whole gene deletion type found in Europeans (Bertelsen et al., 2016) were determined in the above four populations (Table 5). For the SNP g.10583G>A (V172I), two heterozygotes were found in 218 African Americans, resulting in a frequency of 0.5%, but no carriers were detected in the other populations. For the SNP g.13452A>T (R248S), no carriers were found in this study. For the whole gene deletion type, three heterozygotes were found in 209 Caucasians, resulting in a frequency of 0.7%, but no carriers were found in the other populations. Since the allele frequencies of these variants appear to be quite low, further investigation with an increased number of samples would be required to determine more accurate allele frequencies and their ethnic differences.
Allele frequencies of AADAC variants altering enzymatic activities in four populations
Discussion
Eslicarbazepine acetate is a prodrug that has been developed to be efficiently converted to an active form and to lower toxicity by chemical modifications of carbamazepine and oxcarbazepine (Nunes et al., 2013; Galiana et al., 2017). Because eslicarbazepine acetate is an acetylated form of eslicarbazepine, it was surmised to be hydrolyzed by AADAC according to its substrate preference. In this study, we sought to examine whether AADAC is an enzyme responsible for the hydrolysis of eslicarbazepine acetate in humans.
Clinical studies have shown that eslicarbazepine, but not eslicarbazepine acetate, was detected in the blood after oral administration of eslicarbazepine acetate (Almeida et al., 2008), implying that eslicarbazepine acetate is rapidly and completely hydrolyzed in the body. Therefore, we measured eslicarbazepine acetate hydrolase activities in HIMs, HIC, HLMs, and HLC and found that eslicarbazepine acetate was efficiently hydrolyzed by HIMs and HLMs (Fig. 2). Then, it was demonstrated that eslicarbazepine acetate was hydrolyzed by recombinant AADAC but not CES1 or CES2 (Fig. 2). Unlike CES1 and CES2, which are localized both in the endoplasmic reticulum and cytosol (Tabata et al., 2004; Satoh and Hosokawa, 2006), AADAC is specifically localized in the endoplasmic reticulum (Frick et al., 2004). Previously, we demonstrated that the AADAC protein level per total microsomal protein content is almost equal between HIMs and HLMs (Watanabe et al., 2009). This is consistent with the finding that the eslicarbazepine acetate hydrolase activities in HIMs and HLMs were close to each other (Fig. 2). The similarities of Km values (Fig. 3) and the inhibition pattern (Fig. 4) between HIMs, HLMs, and recombinant AADAC, as well as similar inhibition patterns with other AADAC substrates, flutamide, indiplon, and ketoconazole (Watanabe et al., 2009; Shimizu et al., 2014a; Fukami et al., 2016), supported the predominant role of AADAC on eslicarbazepine acetate hydrolysis in the human intestines and liver. The correlation between eslicarbazepine acetate hydrolase activity and AADAC protein expression in human liver samples (Fig. 5) also strengthened the responsibility of AADAC for eslicarbazepine acetate hydrolysis. Collectively, we clearly demonstrated that eslicarbazepine acetate is specifically hydrolyzed by AADAC.
For the compounds used in the inhibition study, it has been reported that the plasma concentration of vinblastine is 200 nM immediately after intravenous administration of the clinical dose (Nelson, 1982). Because the IC50 values of vinblastine for AADAC activities are on the order of μM (Shimizu et al., 2014b), drug-drug interactions between eslicarbazepine acetate and vinblastine are unlikely to be critical in clinical practice. EGCg is a major polyphenolic constituent in green tea, and its concentration in green tea is approximately 25 μM (Dashwood et al., 2002). The plasma concentration of EGCg after taking 20 mg green tea solids/kg body weight, which corresponds to 195 mg EGCg in a person weighing 70 kg, was approximately 0.2 µM (Lee et al., 2002), but the small intestine may be exposed to EGCg at concentrations sufficiently inhibiting AADAC depending on dietary habitat. Since it is known that the bioavailability of EGCg is low (Gan et al., 2018; Dai et al., 2020), in vivo study would be required to clarify whether EGCg intake affects the pharmacokinetics of AADAC substrates in humans.
It is known that the human AADAC gene is genetically polymorphic. Previously, we reported that a SNP of g.148008T>C (X400Q) in AADAC results in a considerable decrease in hydrolase activities for PNPA, flutamide, phenacetine, and rifampicin (Shimizu et al., 2012). In the present study, we found that the V172I variant showed higher activities than the wild type, whereas the R248S or X400Q variants showed lower activities than the wild type (Fig. 6). In the amino acid residues of AADAC, Ser189, Asp343, and His373 form the catalytic triad, which is essential for the catalytic activity (Williams et al., 2010). It is worth examining the possibility that Val172 and Arg248 might be three-dimensionally close to the substrate pocket containing the catalytic triad, leading to the alternation of the structure or size of the substrate pocket. Since the three-dimensional structure of AADAC has not yet been elucidated, progress in research is expected. In this study, the frequency of the AADAC gene deletion type in Caucasians was calculated to be 0.7%, but no carriers were found in the other populations. Originally, the deletion type was found in Europeans, and in a large European cohort, a significant association was observed between AADAC gene deletion and Tourette syndrome, which is a common neurodevelopmental disorder characterized by movement tic and vocal tic (P = 4.4 × 10−4; odds ratio = 1.9, Bertelsen et al., 2016). Another study revealed that Chinese patients with Tourette syndrome possessed an AADAC allele with R248S and an allele with c.361+1G>A, which is predicted to produce mRNA transcripts lacking exon 2, heterozygously (Yuan et al., 2018). Therefore, the R248S variant has been suggested to cause dramatically decreased enzyme activity. Our study could directly prove that the R248S variant has drastically decreased activity. Because the global frequency of c.361+1G>A appears to be quite low (0.008%, Yuan et al., 2018), genotyping of its AADAC variant was not performed in this study. Elucidation of the role of AADAC in neurodevelopment is still awaited.
It has been reported that there was 25-fold interindividual difference in serum concentration of eslicarbazepine/dose ratio in 168 Norwegian patients taking eslicarbazepine acetate (Svendsen et al., 2017). In package insert, it was described that there was no significant difference in the area under the curve of eslicarbazepine between ethnics (Caucasians n = 849, Blacks n = 53, Asians n = 65, and Others n = 51). This is consistent with the results of our study, showing low frequencies of variants affecting enzyme activity. Factors other than AADAC genetic polymorphisms might be involved in the interindividual differences. Eslicarbazepine is further metabolized by UDP-glucuronosyltransferases (Perucca et al., 2011). The interindividual differences in serum level of eslicarbazepine may be accounted for by the differences in the catalytic potency of UDP-glucuronosyltransferase.
In conclusion, we found that eslicarbazepine acetate is specifically hydrolyzed by AADAC in human liver and intestines, and the genetic polymorphism of AADAC could be a factor affecting drug response, although the allele frequencies of variants appear to be low in all populations.
Authorship Contributions
Participated in research design: Hirosawa, Fukami, Tashiro, Nakano, Nakajima.
Conducted experiments: Hirosawa, Tashiro, Kisui.
Contributed new reagents or analytical tools: Hirosawa, Tashiro, Kisui.
Performed data analysis: Hirosawa, Fukami, Tashiro.
Wrote or contributed to the writing of manuscript: Hirosawa, Sakai, Nakajima.
Footnotes
- Received October 24, 2020.
- Accepted January 7, 2021.
This work was supported in part by a Grant-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science [19K07082].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- AADAC
- arylacetamide deacetylase
- BNPP
- bis (p-nitrophenyl) phosphate
- CES
- carboxylesterase
- DFP
- diisopropyl fluorophosphate
- EGCg
- epigallocatechin gallate
- HIC
- human intestinal cytosol
- HIM
- human intestinal microsome
- HLC
- human liver cytosol
- HLM
- human liver microsome
- HPLC
- high-performance liquid chromatography
- P450
- cytochrome P450
- PCR
- polymerase chain reaction
- PMSF
- phenylmethylsulfonyl fluoride
- PNPA
- p-nitrophenyl acetate
- SNP
- single nucleotide polymorphism
- Copyright © 2021 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}