



Abstract
Pharmacokinetic prediction after oral ingestion is important for quantitative risk assessment of food-derived compounds. To evaluate the utility of human intestinal absorption prediction, we compared the membrane permeability and metabolic activities of human induced pluripotent stem cell–derived small intestinal epithelial cells (hiPSC-SIECs) with Caco-2 cells or human primary enterocytes (hPECs). We found that membrane permeability in hiPSC-SIECs had better predictivity than that in Caco-2 cells against 21 drugs with known human intestinal availability (r = 0.830 and 0.401, respectively). Membrane permeability in hiPSC-SIECs was only 0.019–0.25-fold as compared with that in Caco-2 cells for 7 in 15 food-derived compounds, primarily those that were reported to undergo glucuronidation metabolism. The metabolic rates of the glucuronide conjugate were similar or higher in hiPSC-SIECs as compared with hPECs but lower in Caco-2 cells. Expression levels of UDP-glucuronosyltransferase (UGT) isoform mRNA in hiPSC-SIECs were similar or higher as compared with hPECs. Therefore, hiPSC-SIECs could be a useful tool for predicting human intestinal absorption to simultaneously evaluate membrane permeability and UGT-mediated metabolism.
SIGNIFICANCE STATEMENT Gastrointestinal absorption is an important step for predicting the internal exposure of food-derived compounds. This research revealed that human induced pluripotent stem cell–derived small intestinal cells (hiPSC-SIECs) had better predictivity of intestinal availability than Caco-2 cells; furthermore, the metabolic rates of UDP-glucuronosyltransferase (UGT) substrates of hiPSC-SIECs were closer to those of human primary enterocytes than those of Caco-2 cells. Therefore, hiPSC-SIECs could be a useful tool for predicting human intestinal absorption to simultaneously evaluate membrane permeability and UGT-mediated metabolism.
Introduction
Foods consist of a wide variety of compounds from the food itself, from the environment, and also those generated during processing or storage, some of which possess functionality in the living body or have potential for harm (Gibis, 2016; Luca et al., 2020; Thompson and Darwish, 2019). Animal experiments are often used for human risk assessment to comprehensively understand the toxic mechanisms and pharmacokinetics of such compounds, but there are ethical issues and poor predictivity issues for different species (Punt et al., 2017). Well controlled clinical trials are very limited for food-derived compounds and are particularly limited to those that have concern for human health risks. To quantitatively assess whether a food-derived compound exerts effects in vivo, it is important to understand pharmacokinetics after oral ingestion, which can address the internal exposure levels and cellular toxic concentration of the compounds. Alternatives to animal experiments and clinical trials that can directly and quantitatively predict human internal exposure after oral ingestion are required as risk assessment tools.
To predict the internal exposure of food-derived compounds, an initial and important step is gastrointestinal absorption after oral ingestion. The gastrointestinal tract works as a functional barrier made of cells, incorporating nutrients, detoxifying and excreting xenobiotics, and possessing immune function (Shimizu, 2012). Human colon adenocarcinoma–derived Caco-2 cells are widely used to predict the intestinal absorption (Larregieu and Benet, 2013; Vancamelbeke and Vermeire, 2017). Caco-2 cells cultured on a cell culture insert for about 3 weeks form a monolayer membrane and show similar morphologic features to human intestinal epithelial cells, such as intercellular connections and microvilli formation, as well as cellular functions such as efflux transporter P-glycoprotein or breast cancer resistance protein expression on the apical side. However, membrane permeability assays using Caco-2 cells have some limitations. First, permeability of compounds via the paracellular route in Caco-2 cells is very low since the tight junction is strong compared with human small intestinal cells (Takenaka et al., 2016). Second, metabolism of compounds differs from human primary enterocytes (hPECs) because the expression levels and balances of metabolic enzymes such as cytochrome P450s and UDP-glucuronosyltransferases (UGTs) differ from those in human small intestinal epithelial cells (Zhang et al., 2011). UGT is one of the most important enzymes for intestinal absorption of food-derived compounds and extensively metabolizes some of these compounds in the intestine and liver before they enter systemic circulation (Wu et al., 2011). Therefore, there is a long way to go to replace animal experiments with alternative in vitro methods to quantitatively predict the health risks of the compounds. Previously, predictions using parallel artificial membrane permeability assays and metabolic stability assays with human intestinal microsomes were combined with in vitro–in vivo extrapolation techniques (Nishimuta et al., 2011). However, this technique required two separate experiments and might not predict active and paracellular transports. More recently, new cells and methods, such as human small intestinal epithelial cells differentiated from adult intestinal stem cells and Ussing chambers with freshly isolated human jejunum using residual intestinal tissue after human surgery, have been reported to improve predictivity of human intestinal absorption (Michiba et al., 2021; Takenaka et al., 2016); however, these human-derived biomaterials have availability issues.
In recent years, human induced pluripotent stem cell–derived small intestinal epithelial cells (hiPSC-SIECs) have been developed and are commercially available from several vendors. hiPSC-SIECs have been reported to express various functional drug-metabolizing enzymes, such as CYP3A4, and transporters comparable to those in human intestinal epithelial cells (Kabeya et al., 2020; Negoro et al., 2018). Moreover, differentiated hiPSC-SIECs that are easy to use can be obtained from cell vendors, and the intellectual property rights are included in these products for researchers. In addition, hiPSC-SIECs can form a monolayer on cell culture inserts, which cannot be formed by cryopreserved human primary small intestinal cells due to the limited period of viability after thawing. Therefore, the application of hiPSC-SIECs in membrane permeability assays is expected to improve the predictivity of oral availability in vitro by evaluating intracellular metabolism and transporter functions simultaneously. Although several studies have characterized hiPSC-SIECs (Kabeya et al., 2020; Negoro et al., 2018; Yoshida et al., 2021), to our knowledge, reports on their application for membrane permeability assays in combination with metabolic characterization of UGTs for hiPSC-SIEC metabolism of food-derived compounds are limited. In this study, we compared the membrane permeability of hiPSC-SIECs and intestinal metabolic activity, especially by UGTs, with Caco-2 cells and hPECs to evaluate their utility for estimating the oral absorption of food-derived compounds for quantitative human risk assessment.
Materials and Methods
Cells and Reagents.
hiPSC-SIECs were purchased from Fujifilm Corporation (F-hiSIEC, Tokyo, Japan), which included seeding and maintenance medium. Preplated Caco-2 cells were purchased from ReadyCell (CacoReady Plate, Barcelona, Spain), and hPECs were purchased from In Vitro ADMET Laboratories (Columbia, MD). Dulbecco’s modified Eagle’s medium, penicillin-streptomycin solution, nonessential amino acids solution, and FBS were purchased from Thermo Fischer Scientific (Waltham, MA). Hanks’ balanced salt solution (HBSS) and DMSO were purchased from Fujifilm Wako Pure Chemical Industries (Osaka, Japan).
The drugs, food-derived compounds, and metabolites listed below were dissolved in DMSO and stored at −20°C until use. Carbamazepine, verapamil, warfarin, antipyrine, cephalexin, metoprolol, propranolol, hydrochlorothiazide, cimetidine, atenolol, pravastatin, methotrexate, curcumin, epicatechin, epigallocatechin, epigallocatechin gallate, bisphenol A, 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline (MeIQx), and 7-hydroxycoumarin were purchased from Fujifilm Wako Pure Chemical Industries. Piroxicam, acebutolol, sulpiride, acrylamide, fenitrothion, and β-estradiol were purchased from Sigma-Aldrich (St. Louis, MO). Ribavirin, metformin, enalapril, ranitidine, bisphenol S, picloram, and raloxifene were purchased from Tokyo Chemical Industry (Tokyo, Japan). Terbutaline was purchased from LKT Laboratories (St. Paul, MN). Daidzein and genistein were purchased from LC Laboratories (Woburn, MA). Quercetin was purchased from Nacalai Tesque (Kyoto, Japan). 7-Hydroxycoumarin glucuronide, 7-hydroxycoumarin sulfate, daidzein 4′-glucuronide, and daidzein 7-glucuronide were purchased from SantaCruz Biotechnology (Dallas, TX). Raloxifene 4′-glucuronide and raloxifene 6-glucuronide, genistein 4′-glucuronide, and bisphenol A sulfate were purchased from Toronto Research Chemicals (Ontario, Canada). Genistein 7-glucuronide was purchased from Extrasynthese (Lyon France). Estradiol 3-glucuronide and estradiol 17-glucuronide were purchased from Cayman Chemical (Ann Arbor, MI).
TB Green Premix Ex Taq (Tli RNaseH Plus) was purchased from Takara Bio (Shiga, Japan), and specific primer pairs were synthetized in FASMAC (Kanagawa, Japan). Matrigel was purchased from Corning (Corning, NY). Cryopreserved enterocyte recovery medium and hepatocyte/enterocyte incubation medium were purchased from In Vitro ADMET Laboratories.
Cell Culture.
hiPSC-SIECs were thawed and seeded at a density of 1.0 × 105 cells per well on Matrigel-coated 24-well cell culture inserts (Merck Millipore, Burlington, MA), and then maintained for 9–13 days until use according to the manufacturer’s instruction. Caco-2 cells were shipped on day 18 after seeding on 24-well cell culture inserts and maintained with Dulbecco’s modified Eagle’s medium supplemented with 1% nonessential amino acids solution, 10% FBS, and 50 U/ml penicillin/streptomycin for an additional 3–7 days according to the supplier’s instruction manual. hPECs were thawed according to the manufacturer’s instructions and immediately used after plating. Transepithelial electronic resistance (TEER) values were measured with Millicell ERS-2 (Merck Millipore) just before the membrane permeability assays.
Membrane Permeability Assay.
hiPSC-SIECs or Caco-2 cells were washed three times with apical transport buffer [HBSS containing 10 mM 2-(N-morpholino)ethanesulfonic acid (Fujifilm Wako) and 4.5 g/L glucose, pH 6.5] and in basal transport buffer [HBSS containing 10 mM HEPES (Thermo Fischer Scientific) and 4.5 g/L glucose, pH 7.4] chambers and incubated at 37°C for at least 30 minutes. Membrane transport assays were performed at 37°C for 30, 60, and 90 minutes after replenishing the apical chambers with transport buffer containing each substrate at a final concentration of 10 µM. The solution was collected from the basal chambers at 30-minute intervals and diluted with an equal volume of acetonitrile, then stored at −20°C. Unreacted substrates and/or metabolites were measured using ultraperformance liquid chromatography–tandem mass spectrometry. Intestinal availability (Fa or FaFg) data for humans were acquired from publicly available data, and detailed information is shown in Supplemental Table 1.
Metabolic Rate Measurement.
hiPSC-SIECs or Caco-2 cells were cultured in 24-well cell culture inserts as described above, washed three times with transport buffer, and incubated for 30 minutes in a 5% CO2 incubator. hPECs were thawed in cryopreserved enterocyte recovery medium and seeded with serum-free enterocyte incubation medium at a density of 1.5 × 105 cells per well on a flat-bottomed 96-well plate according to the manufacturer’s instructions and incubated for 30 minutes in a 5% CO2 incubator. These cells were exposed to UGT substrates at a concentration of 10 µM for 2 hours. Finally, apical and basal buffers were obtained and diluted with equal volumes of acetonitrile and centrifuged at 10,000g at 4°C for 5 minutes, and supernatants were stored at −20°C until ultraperformance liquid chromatography–tandem mass spectrometry quantification.
mRNA Expression Levels.
Real-time reverse transcription polymerase chain reaction (PCR) was conducted according to previously published methods, with minor modifications (Ohno and Nakajin, 2009). Total RNA for hiPSC-SIECs, Caco-2 cells, and hPECs was extracted with RNeasy Plus Mini kit and RNase free DNase (Qiagen, Hilden, Germany). Reverse transcription was carried out using high-capacity RNA-to-cDNA kits (Thermo Fischer Scientific) according to the manufacturer’s recommendations, using 5 ng of total RNA as the template. Quantification of mRNA expression levels for human UGT isoforms was performed using the ViiA 7 system (Applied Biosystems, Foster City, CA) in a total reaction volume of 20 µl per well, which consisted of 0.5 µM of each of the specific primer pairs (Table 1) and 2 µl of cDNA solution. PCR amplification consisted of an initial 30-second denaturation step at 95°C, followed by 40 cycles of denaturation at 95°C for 5 seconds, and annealing and extension at 60°C for 30 seconds. Specificity of the PCR amplification was confirmed via analysis of the melting curve.
Primer sequences used for real-time polymerase chain reaction analysis
Data Analysis.
Apparent permeability coefficient (Papp) values were calculated as follows:
 where dQ/dt, A, and C0 represent the amount of permeated compound per unit of time, the surface area of the transport membrane, and the initial compound concentration in the donor chamber, respectively. The relationship between apical-to-basal Papp and Fa or FaFg was calculated in accordance with previous reports (Takenaka et al., 2016 and Amidon et al., 1988):
where dQ/dt, A, and C0 represent the amount of permeated compound per unit of time, the surface area of the transport membrane, and the initial compound concentration in the donor chamber, respectively. The relationship between apical-to-basal Papp and Fa or FaFg was calculated in accordance with previous reports (Takenaka et al., 2016 and Amidon et al., 1988):
 where Psf is the scaling factor. Nonlinear regression was used for curve fitting with JMP version 15 (SAS Institute, Cary, NC). Relative gene expression data were analyzed using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
where Psf is the scaling factor. Nonlinear regression was used for curve fitting with JMP version 15 (SAS Institute, Cary, NC). Relative gene expression data were analyzed using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Results
Relationships Between the Papp and Fa or FaFg Using 21 Known Drugs.
The TEER values of monolayers of hiPSC-SIECs and Caco-2 cells were 882 ± 258 (n = 42) and 1348 ± 377 (n = 63) Ω·cm2, respectively. The relationships between Papp and Fa or FaFg for the 21 drugs investigated in hiPSC-SIECs and Caco-2 cells are shown (Fig. 1, A and B). hiPSC-SIECs had a better correlation between Papp and Fa or FaFg (r = 0.830) than Caco-2 cells (r = 0.401). The estimated Psf values of hiPSC-SIECs and Caco-2 cells were 3.38 ± 0.447 and 2.98 ± 0.815 × 106 s/cm, respectively.

Relationship between Papp values and intestinal availability (Fa or FaFg) for 21 drugs. hiPSC-SIECs (A) or Caco-2 cells (B) were incubated with transport buffer containing each of the 21 different drugs for 90 minutes at 37°C. The regression curves between the Papp and Fa or FaFg of these 21 drugs were fitted to the following formula in accordance with previous reports (Takenaka et al., 2016; Amidon et al., 1988): Fa or FaFg = 1 − e−Psf×Papp. Compounds used were as follows: 1, piroxicam; 2, carbamazepine; 3, verapamil; 4, warfarin; 5, antipyrine; 6, cephalexin; 7, metoprolol; 8, propranolol; 9, acebutolol; 10, ribavirin; 11, metformin; 12, hydrochlorothiazide; 13, terbutaline; 14, cimetidine; 15, enalapril; 16, ranitidine; 17, atenolol; 18, sulpiride; 19, pravastatin; 20, methotrexate; and 21, raloxifene. Data are presented as means for hiPSC-SIECs (n = 2) and means ± S.D. for Caco-2 cells (n = 3).
Comparisons of Papp Across Food-Derived Compounds.
Membrane permeabilities were compared between 15 food-derived compounds in hiPSC-SIECs and Caco-2 cells (Table 2). The Papp values of daidzein, genistein, quercetin, curcumin, bisphenol A, bisphenol S, and MeIQx in hiPSC-SIECs were lower than those in Caco-2 cells. These compounds were reported to undergo glucuronidation, and the Papp values of hiPSC-SIECs ranged from 0.019 (curcumin) to 0.25-fold (bisphenol S) compared with those of Caco-2 cells. The Papp values of caffeic acid and gallic acid in hiPSC-SIECs were higher than those in Caco-2 cells. The Papp values of epicatechin, epigallocatechin, epigallocatechin gallate, acrylamide, fenitrothion, and picloram were within 0.71–1.3-fold between hiPSC-SIECs and Caco-2 cells.
Papp of food-derived compounds in hiPSC-SIECs and Caco-2 cells
Comparisons of the Amounts of Unchanged and Glucuronidated Compounds Between hiPSC-SIECs and Caco-2 Cells.
The amounts of unchanged compounds and glucuronide or sulfated metabolites were compared with raloxifene, daidzein, genistein, and bisphenol A after performing the membrane permeability assays using hiPSC-SIECs and Caco-2 cells (Table 3). The total glucuronide produced in hiPSC-SIECs was 6.4- (raloxifene) to 40- (daidzein) fold higher than that in Caco-2 cells. The total amount of unchanged compounds in basal compartments and metabolites in apical and basal compartments were 0.9- (daidzein) to 2.7- (raloxifene) folds between hiPSC-SIECs and Caco-2 cells.
The amounts of unchanged compounds and metabolites after the membrane permeability assays in hiPSC-SIECs and Caco-2 cells.
Unchanged (basal)2.638.2 ± 5.2 Unchanged (basal)49504 ± 31 Unchanged (basal)167450 ± 14 Unchanged (basal)111325 ± 48
Comparisons of Glucuronidation Rates Between hiPSC-SIECs, Caco-2 Cells, and hPECs Using Typical UGT Substrates and Food-Derived Compounds.
We compared the metabolic rates of three typical UGT substrates (7-hydroxycoumarin, raloxifene, and estradiol) and three food-derived compounds (daidzein, genistein, and bisphenol A) in hiPSC-SIECs, Caco-2 cells, and hPECs (Table 4). The metabolic rates of UGT substrates were similar or slightly higher in hiPSC-SIECs (0.14–18-fold), and lower in Caco-2 cells (0.0083–0.90-fold), as compared with those in hPECs, except 7-hydroxycoumarin in Caco-2 cells (6.5-fold increase) and genistein 4′-glucuronide, which was neither detected in hiPSC-SIECs nor in Caco-2 cells. The metabolic rates of other known metabolites, such as 7-hydroxycoumarin sulfate and bisphenol A sulfate in hiPSC-SIECs and Caco-2 cells, were within 0.28–3.8-fold compared with those rates in hPECs.
Comparison of metabolic rates of typical UGT substrates and food-derived compounds between hiPSC-SIECs, Caco-2 cells, and hPECs.
Comparisons of mRNA Expression Levels for UGT Isoforms Between hiPSC-SIECs, Caco-2 Cells, and hPECs.
Figure 2 shows the results of the relative mRNA expression levels of representative UGT isoforms by real-time reverse transcription PCR in hiPSC-SIECs and Caco-2 cells compared with hPECs. The expression levels of small intestine–specific UGT isoforms UGT1A8 and UGT1A10 in hiPSC-SIECs were within 10-fold of the levels in hPECs (1.4-fold for UGT1A8 and 0.13-fold for UGT1A10, respectively), whereas those in Caco-2 cells were quite low (0.027-fold for UGT1A8 and 0.0017-fold for UGT1A10, respectively). The expression levels of UGT isoforms in hiPSC-SIECs and Caco-2 cells such as UGT1A6, UGT1A9, and UGT2B7, which are not limited to the small intestinal cells, were similar to those of hPECs. The expression levels of UGT1A1 and UGT2B15 were higher in hiPSC-SIECs than in hPECs (95-fold and 127-fold for UGT1A1 and UGT2B15, respectively).
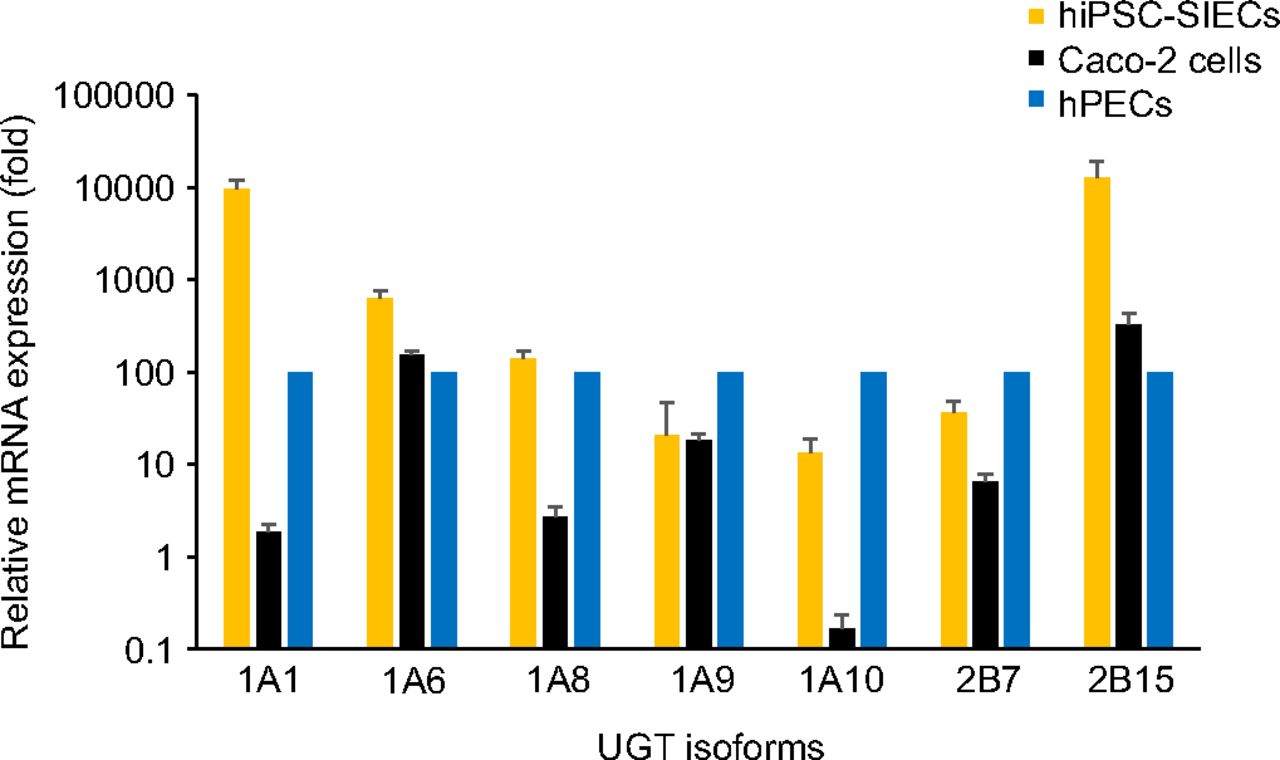
Relative expression levels of genes encoding UGT isoforms of hiPSC-SIECs and Caco-2 cells. Real-time reverse transcription polymerase chain reaction analysis of the expression levels of UGT isoform-related proteins. mRNA levels were normalized to those of glyceraldehyde-3-phosphate dehydrogenase mRNA gene expression levels in the hPECs, the levels of which were arbitrarily defined as 100. The gene expression levels of hPECs were measured in and averaged across two separate lots. Data are presented as means ± S.D. (n = 3) for hiPSC-SIECs and Caco-2 cells.
Discussion
When membrane permeability of hiPSC-SIECs and Caco-2 cells were compared using drugs with known intestinal availability, Papp values for the high-membrane-permeability drugs (Papp > 10 × 10−6 cm/s) were similar between hiPSC-SIECs and Caco-2 cells. On the other hand, Papp differed between hiPSC-SIECs and Caco-2 cells for the low-to-medium permeability drugs (Papp < 10 × 10−6 cm/s). hiPSC-SIECs had a better correlation between Papp and Fa or FaFg than Caco-2 cells, especially with low-to-medium Fa or FaFg compounds. It is unclear why, but high-permeability drugs are mainly passed via transcellular routes, whereas low-to-medium absorption compounds are passed via transcellular and paracellular routes. Since hiPSC-SIECs have lower TEER values than Caco-2 cells, it might be due to the contribution of the paracellular route, as previously discussed (Kabeya et al., 2020). Similarly, the Papp of food-derived compounds, including caffeic acid and gallic acid, which have been reported to be passed through the paracellular route (Konishi et al., 2003; Konishi and Kobayashi, 2004), differ between hiPSC-SIECs and Caco-2 cells, supporting this explanation. Another reason might be the difference in intracellular permeability between hiPSC-SIECs and Caco-2 cells. Although total concentrations for unchanged compounds and metabolites were similar between Caco-2 cells and hiPSC-SIECs (Table 3), we could not ignore the possibility of a difference in membrane permeability between the two types of cells because we could not measure the concentration of the refluxed unchanged substrates to apical compartments. Another explanation might be the involvement of intestinal transporters. Konishi et al. reported that caffeic acid is incorporated via monocarboxylic transporter 1 but gallic acid is not (Konishi et al., 2003; Konishi and Kobayashi, 2004). In our study, the fold increase in the Papp value of caffeic acid between hiPSC-SIECs and Caco-2 cells was higher than the fold increase of caffeic acid (5.7-fold versus 1.7-fold), suggesting the involvement of transporter-mediated uptake as well as paracellular transport. Methotrexate is known as a substrate for both influx transporter [proton-coupled folate transporter (PCFT)] and efflux transporters [multidrug resistance-associated protein 2 (MRP2), breast cancer resistance protein (BCRP)], and genistein is a substrate for efflux transporters [multidrug resistance-associated protein 2 (MRP2)] (Yokooji et al., 2007, Yokooji et al., 2009, and Takaai et al., 2010, Kobayashi et al., 2013). Further studies are needed to understand the detailed mechanisms of these compounds. Based on these results, it was suggested that hiPSC-SIECs could predict the human intestinal availability better than Caco-2 cells.
When predicting the human intestinal absorption, it is important to consider not only the intestinal permeability but also metabolism in intestinal cells. For example, several compounds in food, such as daidzein and bisphenol A, are known to be metabolized by UGTs, and over 99% of these compounds were present as glucuronide metabolites in human serum (Setchell et al., 2001; Thayer et al., 2015). The Papp values of these compounds in hiPSC-SIECs were 4–53-fold lower than those in Caco-2 cells, suggesting the possible involvement of UGT metabolism in hiPSC-SIECs in membrane permeability assays. Additionally, we observed that hiPSC-SIECs produced more glucuronidated metabolites for raloxifene, daidzein, genistein, and bisphenol A than Caco-2 cells during the membrane permeability assays. Thus, we speculated that hiPSC-SIECs might have the potential to predict human intestinal absorption of food-derived compounds via simultaneous evaluation of membrane permeability and intestinal cellular metabolism. The metabolic rates of glucuronide in hiPSC-SIECs were quantified and found to be similar or higher than those in hPECs, which were found to be much greater than in Caco-2 cells, excepting only for 7-hydroxycoumarin and bisphenol A for Caco-2 cells. Finally, to account for the differences of UGT activities, we further compared the mRNA expression levels of UGT isoforms between hiPSC-SIECs, Caco-2 cells, and hPECs. UGT mRNA expression experiments indicated the same order (hiPSC-SIECs > hPECs ≫ Caco-2 cells), excepting only UGT1A6 and UGT2B15 for Caco-2 cells, the expressions of which were similar between Caco-2 cells and hPECs.
7-Hydroxycoumarin is mainly glucuronidated by UGT1A6 and UGT1A9, both of which are tissue-nonspecific UGT isoforms (Moeller et al., 2011). Metabolic rates of 7-hydroxycoumarin are slightly higher in hiPSC-SIECs and Caco-2 cells than those in hPECs. Expression levels of UGT1A6 were also higher in hiPSC-SIECs and Caco-2 cells. This may account for differences in the metabolic rates of 7-hydroxycoumarin glucuronide in these cells. Previous reports have indicated that the UGT1A6 activity in Caco-2 cells is higher than that in hPECs, consistent with our current study (Zhang et al., 2011). The metabolic rates of another metabolite, 7-hydroxycoumarin sulfate, were similar in hiPSC-SIECs and Caco-2 cells as compared with in hPECs. Therefore, we concluded that the metabolic rates of sulfate conjugate did not cause a significant impact in interpreting these results.
Raloxifene is known to have two glucuronidation sites at the 4′-OH position and 6-OH position. The 4′-OH position is conjugated by UGT1A8 and UGT1A10, which are small intestine–specific UGT isoforms, whereas the 6-OH position is conjugated by UGT1A1 and UGT1A8 (Kemp et al., 2002). As shown in Fig. 1, Papp of raloxifene in hiPSC-SIECs were lower than those in Caco-2 cells. The metabolic rates of 4′-glucuronide were 0.96- and 0.014-fold in hiPSC-SIECs and Caco-2 cells, respectively, compared with those in hPECs. We also observed the expression levels of UGT1A8 and UGT1A10 were within 10-fold in hiPSC-SIECs, and far lower in Caco-2 cells compared with hPECs, which may explain the difference in metabolic rates of raloxifene 4′-glucuronide. The metabolic rates of 6-glucuronidated form were 12- and 0.030-fold in hiPSC-SIECs and Caco-2 cells as compared with hPECs. We also observed the expression levels of UGT1A1 were 119- and 0.024-fold in hiPSC-SIECs and Caco-2 cells as compared with hPECs. These results may explain the difference in metabolic rates for raloxifene 4′-glucuronide and 6-glucuronide among these three cell types.
β-Estradiol was determined to be a specific substrate for UGT1A1 by measuring the metabolic rate of β-estradiol 3-glucuronide (Lv et al., 2019b). The metabolic rates of β-estradiol 3-glucuronide were comparable in hiPSC-SIECs (3.8-fold) and lower in Caco-2 cells (0.017-fold) as compared with that in hPECs; these results also support intestine-specific UGT activities in hiPSC-SIECs. Metabolic rates of another metabolite, β-estradiol 17-glucuronide, were similar among the three cell types. This metabolite was reported to possibly be catalyzed by UGT1A10, UGT2B7, and UGT2B17 (Asai et al., 2017).
Daidzein and genistein are also known to have two glucuronidation sites at the 4′-OH position and 7-OH position. There are some reports detailing daidzein metabolism in the intestine and the liver (Hanioka et al., 2018) showing that the 4′-glucuronidated form is mainly generated in the small intestine. The metabolic rates of these 4′-glucuronidated forms were 0.14-fold in hiPSC-SIECs and 0.0083-fold in Caco-2 cells as compared with those in hPECs. However, the metabolic rates of the 7-glucuronide were 7.1-fold in hiPSC-SIECs and 0.050-fold in Caco-2 cells as compared with those in hPECs.
Bisphenol A is known to undergo glucuronidation mainly by UGT1A9 and UGT2B15, which are abundant in the liver (Hanioka et al., 2008). These metabolic rates were 21-fold in hiPSC-SIECs and 0.5-fold in Caco-2 cells compared with hPECs. The relative mRNA expression levels of UGT2B15 were 226-fold in hiPSC-SIECs and 3.2-fold in Caco-2 cells as compared with hPECs, which may explain the difference in metabolic rates of bisphenol A.
Based on these results, it was strongly suggested that the activities and expressions of intestine-specific UGT1A8 and UGT1A10 in hiPSC-SIECs were comparable with hPECs, whereas those in Caco-2 cells were much lower. However, it was also suggested that nonspecific UGT isoforms such as UGT1A1 and UGT2B15 in hiPSC-SIECs were higher than those of hPECs. hiPSC-SIECs have great potential to predict intestinal absorption, but there are some limitations. For example, raloxifene 4′-glucuronide was reported to be present as a major metabolite in human plasma, because the intrinsic clearance of the 4′-glucuronidated form is 5.6-fold higher than the 6-glucuronidated form in intestinal microsomes (Kemp et al., 2002). We observed the metabolic rates of raloxifene 4′-glucuronide in hiPSC-SIECs were comparable to those in hPECs, but the metabolic rates of raloxifene 6-glucuronide were 12-times higher than in hPECs; thus, metabolite ratios between the 4′-glucuronide and 6-glucuronide forms were different (4′-glucuronide: 6-glucuronide ratios = 0.60 in hiPSC-SIECs and 7.9 in hPECs). A similar result was observed for daidzein. Although previous reports found that the metabolic ratio of 4′-glucuronide and 7-glucuronide in human intestinal microsomes was 1.7 (Hanioka et al., 2018), metabolite ratios between the 4′-glucuronide and 7-glucuronide forms were different (4′-glucuronide: 7-glucuronide ratios = 112 in hiPSC-SIECs and 2.2 in hPECs). We therefore considered that hiPSC-SIECs reproduced glucuronic metabolism in small intestinal cells but was not exactly the same in terms of UGT isoforms examined in this study. There are some limitations to our conclusion; because specific inhibitors for UGT isoforms are limited and the substrate specificity of UGT overlaps between each isoform (Lv et al., 2019a), we could not confirm the molecular mechanisms. Further studies might be needed to confirm our speculation. We still believe that hiPSC-SIECs could be an improvement in predicting human intestinal absorption of unchanged compounds, because when considering the overall metabolic clearance rates of the unchanged compounds in this study, the metabolic clearance rates of the unchanged compounds ranged from 0.96 to 18-fold in hiPSC-SIECs and 0.017 to 6.5-fold in Caco-2 cells as compared with in hPECs. Therefore, hiPSC-SIECs might have closer characteristics to hPECs than Caco-2 cells. Further studies might improve upon these metabolic characteristics. Since we found that UGT activities and mRNA expression levels were correlated, expression of UGT isoforms could be valuable to evaluate the cellular properties of hiPSC-SIECs from individual vendors. In addition, application for human plasma concentration prediction should be conducted in combination with other in vitro assays such as hepatocyte metabolic stability and plasma protein binding assays, and in silico prediction assays.
In conclusion, this study is the first report to simultaneously compare hiPSC-SIECs, Caco-2 cells, and hPECs in terms of membrane permeability and intracellular UGT metabolism. We conclude that hiPSC-SIECs might be useful for predicting the human intestinal absorption of food-derived compounds related to UGT conjugation enzymes for quantitative human risk assessment.
Authorship Contributions
Participated in research design: Kitaguchi, Mizota, Ito, Ohno, Kobayashi, Ogawa, Qiu, Iwao, Hanioka, Tanaka, Matsunaga.
Conducted experiments: Kitaguchi, Mizota, Ito.
Performed data analysis: Kitaguchi, Ito.
Wrote or contributed to the writing of the manuscript: Kitaguchi, Mizota, Ohno, Kobayashi, Iwao, Hanioka, Matsunaga.
Footnotes
- Received July 1, 2021.
- Accepted October 18, 2021.
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

ABBREVIATIONS
- Fa or FaFg
- intestinal availability
- HBSS
- Hanks’ balanced salt solution
- hiPSC-SIEC
- human induced pluripotent stem cell–derived small intestinal epithelial cell
- hPEC
- human primary enterocyte
- MeIQx
- 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline
- Papp
- apparent permeability coefficient
- PCR
- polymerase chain reaction
- TEER
- transepithelial electronic resistance
- UGT
- UDP-glucuronosyltransferase
- Copyright © 2021 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}