


Abstract
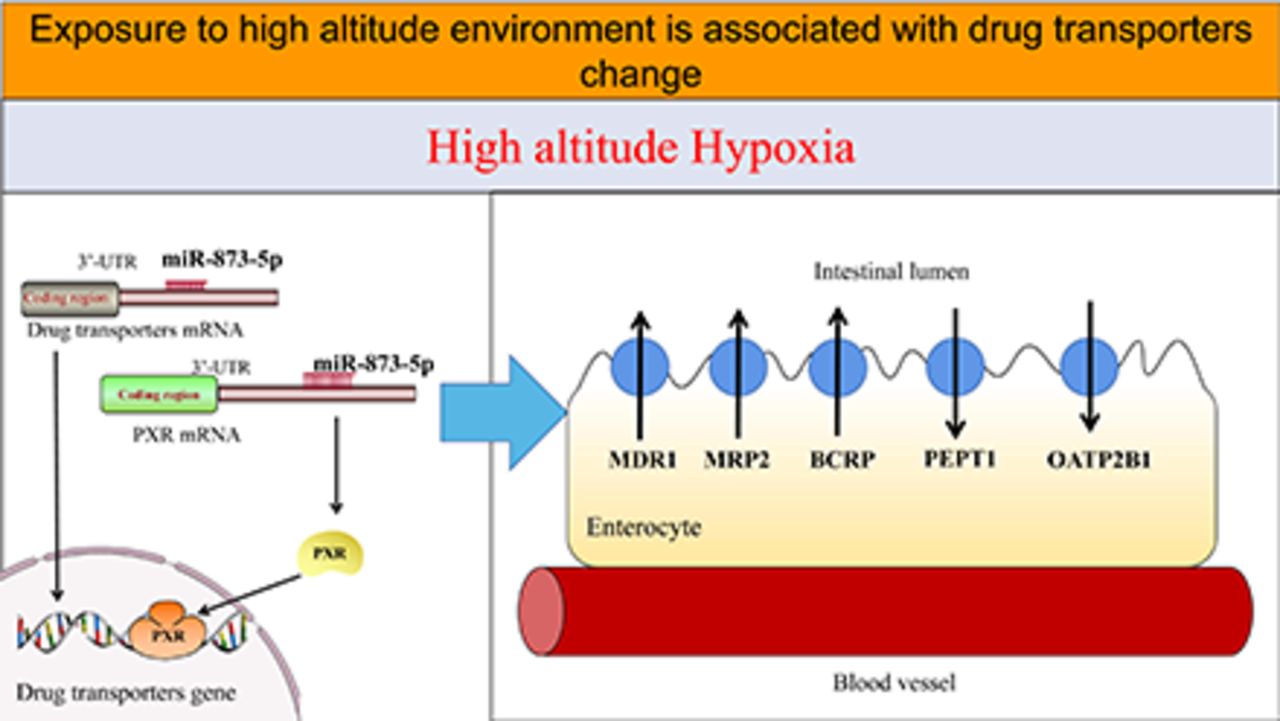
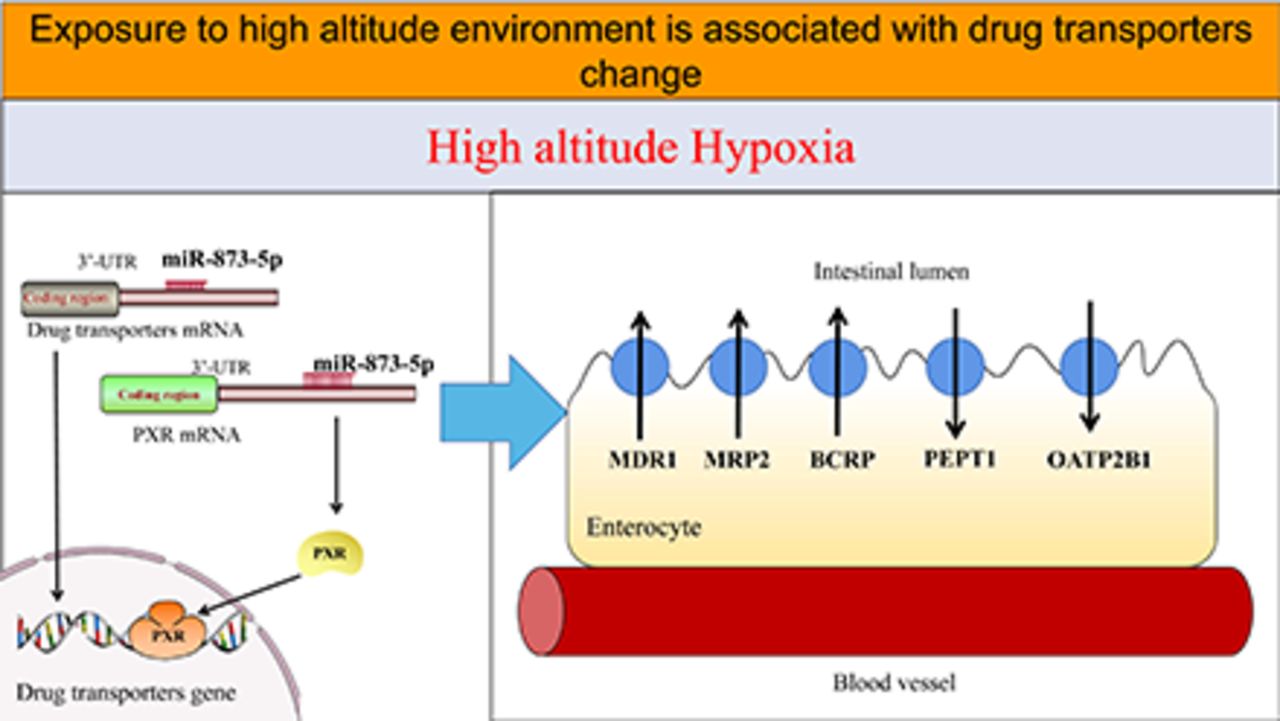
Hypoxia is the main characteristic of a high-altitude environment, affecting drug metabolism. However, so far, the mechanism of microRNA (miRNA) involved in the regulation of drug metabolism and transporters under high-altitude hypoxia is still unclear. This study aims to investigate the functions and expression levels of multidrug resistance protein 1 (MDR1), multidrug resistance-associated protein 2 (MRP2), breast cancer resistance protein (BCRP), peptide transport 1 (PEPT1), and organic anion-transporting polypeptides 2B1 (OATP2B1) in rats and colon cancer (Caco-2) cells after exposure to high-altitude hypoxia. The protein and mRNA expression of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 were determined by Western blot and qPCR. The functions of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 were evaluated by determining the effective intestinal permeability and absorption rate constants of their specific substrates in rats under high-altitude hypoxia, and uptake and transport studies were performed on Caco-2 cells. To screen the miRNA associated with hypoxia, Caco-2 cells were examined by high throughput sequencing. We observed that the miR-873-5p was significantly decreased under hypoxia and might target MDR1 and pregnane X receptor (PXR). To clarify whether miR-873-5p regulates MDR1 and PXR under hypoxia, Caco-2 cells were transfected with mimics or inhibitors of miR-873-5p and negative control (NC). The function and expression of drug transporters were found to be significantly increased in rats and Caco-2 cells under hypoxia. We found that miR-873-5p regulated MDR1 and PXR expression. Herein, it is shown that miRNA may affect the expression of drug transporter and nuclear receptor under hypoxia.
SIGNIFICANCE STATEMENT This study explores if alterations to the microRNAs (miRNAs), induced by high-altitude hypoxia, can be translated to altered drug transporters. Among miRNAs, which show a significant change in a hypoxic environment, miR-873-5p can act on the multidrug resistance protein 1 (MDR1) gene; however, there are multiple miRNAs that can act on the pregnane X receptor (PXR). This study speculates that the miRNA-PXR-drug transporter axis is important in the physiological disposition of drugs.
Introduction
Since ancient times, it has been a long-known fact that human beings can experience dyspnea caused by hypoxia in high-altitude areas. The first clear description of mountain sickness experienced by lowland natives at high altitudes was written by the priest Jose de Acosta in 1590 (Gilbert et al., 1983). The prominent feature of a high-altitude environment is solar radiation, decreased ambient oxygen tension, extreme diurnal range of temperature, arid climate, poor soil quality, etc., but the most striking feature is hypoxia (Eide and Asplund, 2012). With the increase in altitude, the atmospheric pressure and partial pressure of oxygen decreased. The partial pressure inhaled, alveolar oxygen, and alveolar oxygen diffusion into the blood had corresponding changes due to the decrease in the partial pressure of oxygen in the atmosphere. In addition, studies found that the hypoxemia and tissue hypoxia, caused by arterial oxygen pressure and oxygen saturation decrease, become more intense as a result of altitude change (Simon and Simon, 2014).
The circulatory, nervous, and endocrine systems are adversely influenced by high-altitude hypoxia, which alters essential bodily functions, metabolism, and cell structures and contributes to altitude sickness (Netzer et al., 2013). Altitude sickness is influenced by the speed of entering the hypoxic areas of the plateau, the length of stay, and the unique metabolism and status of individuals. Slow entry into high altitude elicits a compensatory response, and rapidly moving to higher altitudes causes tissue and cellular metabolic dysfunction and functional disorders. Additionally, acute hypoxia can directly cause altitude sickness, pulmonary edema, and cerebral edema, whereas chronic hypoxia induces polycythemia and cardiovascular diseases (Mairbäurl, 1994; Imray et al., 2010).
Research on high-altitude hypoxia has focused on high-altitude disease prevention, nutrition, and high-altitude plateau adaptation (Rashid et al., 2013); recently, emphasis has been laid on high-altitude hypoxia and drug metabolism (Zhang et al., 2019). The drug absorption, distribution, metabolism, and excretion (ADME) are changed under hypoxic conditions, which alter the pharmacokinetics (Vij et al., 2012). Cytochrome P450 (CYP450) enzymes and drug transporters play an important role in the physiologic disposition of drugs, and the chemical and nonchemical factors affect the expression of the CYP450 family and transporters family gene at the transcriptional level (Nivoix et al., 2008; Yijie et al., 2019). There are many studies conducted on nonchemical factors, such as biophysical influences, pathologic states, diseases, and constitutional features, but it is also necessary to carry out more typical research on the effect of hypoxia on CYP450 and transporters. (Vesell, 1977; Jones et al., 1989).
Numerous studies have shown that the pharmacokinetics of lidocaine hydrochloride, sulfamethoxazole, lithium, acetazolamide, meperidine, and prednisolone significantly changed at high altitudes (Arancibia et al., 2003; Li et al., 2009; Li et al., 2012a; Zhang et al., 2016). Several previous studies have found increased expression of CYP3A6 and decreased expression of CYP1A1, 1A2, 2B4, 2C5, and 2C16 in rabbits after acute exposure to hypoxia (Kurdi et al., 1999; Fradette et al., 2002; Fradette and Du Souich, 2004; Fradette et al., 2007). In our previous study, we reported that CYP1, CYP2, CYP3, and NAT2 expressions significantly changed in rats at high altitudes. (Li et al., 2012b; Li et al., 2014a; Li et al., 2014b). Drug transporters play a key role in the physiologic disposition of drugs, which is also influenced by chemical and nonchemical factors. All these effects will change the effectiveness of drugs and further impact the clinical treatment. Schöning et al. found that low-oxygen tumor environments induced the activity of multidrug resistance protein 1 (MDR1), multidrug resistance-associated protein 1 (MRP1), and breast cancer resistance protein (BCRP) by activating and increasing the expression of these transporters in several cancer cell lines (Schöning et al., 2017).
MicroRNAs (miRNAs) are small endogenous RNAs that regulate gene expression. Research on the role of miRNA in drug metabolism is expanding because miRNAs are key regulators of gene expression (Lu and Rothenberg, 2018). He et al. reviewed the effects of miRNA on the ADME of drugs and summarized a hypothetical network that comprises 189 regulatory relationships between miRNA and their genes, and involves 120 miRNAs and 261 ADME genes, such as miR-148a-regulated pregnane X receptor (PXR), miR-92a-regulated peptide transport 1 (PEPT1), miR-7, miR-27a, miR-451-regulated MDR1, etc. (He et al., 2015).
However, to date, there are no reports that resolve the problem of changes in the activity and expression of drug transporters due to high-altitude hypoxia. In addition, the role of miRNA in regulating drug transporters under hypoxia is not clear. Consequently, we investigated the function and expression levels of MDR1, MRP2, BCRP, PEPT1, and organic anion-transporting polypeptides 2B1 (OATP2B1) and their mechanism in the hypoxic environment at high altitudes.
Materials and Methods
Chemicals.
Primary antibodies of β-Actin (1:500, mouse monoclonal, Abcam, ab8226), MDR1 (1:1000, rabbit monoclonal, Abcam, ab170904), MRP2 (1:1000, rabbit polyclonal, Sigma-Aldrich, M8316), BCRP (1:500, rabbit monoclonal, Abcam, ab207732), PEPT1 (1:500, rabbit polyclonal, Thermo Fisher, PA5-37010), and OATP2B1 (1:500, rabbit polyclonal, Novus, NBP-1-59811) were used for Western-blot test. RNAiso Plus and PrimeScript RT Reagent were obtained from Takara Bio Inc. (Kyoto, Japan). The primers used for real-time PCR were synthesized by Shanghai Sangon Biotech Co., Ltd. (Shanghai, China). Rhodamine 123 (Lot: L630S74), Vincristine (Lot: LQCOS60), Prazosin (Lot: L780R01), Ubenimex (Lot: B3XEH-DO), and Glibenclamide (Lot: L910Q73) were purchased from J&K Scientific Co., Ltd. (Beijing, China). Colon cancer (Caco-2) cell line was obtained from the Chinese Academy of Sciences Cell Bank (Shanghai, China). Fetal bovine serum was purchased from Biologic Industries Co., Ltd. (Shanghai, China). LC-MS (Liquid Chromatography Mass Spectrometry) grade methanol, acetonitrile, and ultrapure water were purchased from Thermo Fisher Scientific (Shanghai, China). Lipofectamine 3000 Transfection Reagent (Lot: L3000015) was purchased from Invitrogen Biotech Co., Ltd. (CA).
Treatment of Animals.
A total of 250 male Sprague Dawley rats (200 ±20 g; Certificate No. 2018-001) of Specific Pathogen Free grade were provided by Xi’an Jiaotong University Medical College. Ten animals were housed per cage with separate rooms to ensure that each of them could be restricted to a single space to avoid restraint stress. All rats were kept at room temperature at 23 ±2°C with constant humidity of 55% ±5% under a 12-hour light/dark cycle for one week. The experimental procedures were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and the protocol was approved by the Animal Ethics Committee of The Qinghai University.
Two hundred and fifty male rats were randomly divided into MDR1, MRP2, BCRP, PEPT1, and OATP2B1 groups, and every group had 50 rats. MDR1, MRP2, BCRP, PEPT1, and OATP2B1 groups were further randomly divided into five subgroups, respectively, and every group had 10 rats. The plain (P) group (altitude: 390 m, PaO2: 20 kPa) included 50 rats living in the city of Xi’an in Shanxi Province of northwest China. The acute moderate-altitude hypoxia (AMH) group (altitude: 2,800 m, PaO2: 15.1 kPa) and the chronic moderate-altitude hypoxia (CMH) group (altitude: 2,800 m, PaO2: 15.1 kPa) included 100 rats living in the city of Xi’an, but they received a 24-hour acute exposure and a 30-day chronic exposure to a middle-altitude area. The rats in these two groups were transported by bus to Gonghe County, Qinghai Province of northwest China, located at an altitude of 2,800 m. The rats arrived in Gonghe County at 3:00 PM, and the tissues of each rat in the AMH group were collected at 3:00 PM the following day after single-pass intestinal perfusion. Then, the tissues of rats in the CMH group were collected at 3:00 PM on the 31st day after single-pass intestinal perfusion. The acute high-altitude hypoxia (AHH) group (altitude: 4,300 m, PaO2: 12.4 kPa) and the chronic high-altitude hypoxia (CHH) group (altitude: 4,300 m, PaO2: 12.4 kPa) consisted of 100 rats living in the city of Xi’an; however, the rats in the AHH group were acutely exposed to high altitudes for 24 hours, and the rats in the CHH group were chronically exposed to high altitudes for 30 days. The rats of AHH and CHH groups were transported to Maduo County by bus, which is in Qinghai Province of northwest China, to an altitude of 4300 m. Similar to the moderate-altitude experiment, the rats arrived in Maduo County at 9:00 PM, and the tissues of each rat in the AHH group were collected at 9:00 PM the following day and of rats in the CHH group at 9:00 PM on the 31st day after single-pass intestinal perfusion.
Sample collection of the P group was performed at Xi’an Jiaotong University Medical College, whereas the sample collection of the AMH and CMH groups was carried out at Gonghe County Hospital. Moreover, the sample collection of the AHH and CHH groups was performed at Maduo County Hospital. To determine the expression and function of drug transporters, all tissues were stored in liquid nitrogen and were transported to the Research Center for High Altitude Medicine of Qinghai University by bus.
The absorption of the specific substrates of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 was measured in the rats’ intestines by the intestinal perfusion method. The concentration of each substrate (MDR1: Rhodamine 123, MRP2: Vincristine, BCRP: prazosin, PEPT1: Ubenimex, OATP2B1: Glibenclamide) was determined by a high-performance liquid chromatography (HPLC) system (Agilent 1200, CA).
Evaluation of the Functions of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 in Rats.
The functions of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 were evaluated by the single-pass intestinal perfusion method. Before experiments, rats were immediately anesthetized via enterocoelia injection with sodium pentobarbital (50 mg/kg). After this, the rats were placed on a heated operating table to keep the body temperature at 37°C. All rats were given laparotomy, and a 10–15 cm intestinal segment (duodenum, jejunum, and ileum) was isolated. Both ends of the intestinal segment were cannulated with plastic tubing. The intestinal segment was washed with sterile normal saline at 37°C to clear the content. Next, intestinal perfusion was performed with Krebs-Ringer solution that contained phenol red and substrates of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 at 37°C. The intestine was perfused for 30 minutes at a rate of 1 mL/min to stabilize the state of rats and then perfused for 120 minutes at a rate of 0.2 mL/min by a peristaltic pump (BT100-2J, Heibei, China). The Krebs-Ringer solution and the collection bottle were replaced every 15 minutes. The intestine was cut off at both ends of the intubation, and the length and inner diameter of the intestine were measured. The substrates of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 are shown in Table 1. The absorption rate constant (Ka, s−1) and intestinal effective permeability (Peff, cm/s) of the substrates were calculated according to the following formula (Chen et al., 2010; Li et al., 2017):

The substrates of MDR1, MRP2, BCRP, PEPT1, and OATP2B1
Where PRout and PRin are the concentrations of outlet and inlet phenol red. Cout (corrected) is corrected outlet drug concentration; Cout is outlet drug concentration; and Cin is inlet drug concentration (μg/mL). Q is the flow rate in mL/min (0.2 mL/min), l is the length of the intestinal segment, and r is the radius of the intestinal segment.
HPLC Analysis.
The concentrations of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide in the Krebs-Ringer solution were determined by HPLC. The Krebs-Ringer solution sample was processed with acetonitrile to denature the protein, and the mixture of acetonitrile and Krebs-Ringer solution was vortexed for 0.5 minute and then centrifuged for 10 minutes at 16,000 g at 4°C. A 10 μL aliquot of the supernatant was injected into the HPLC system. A C18 column (Agilent, Agilent Analytics, Inc., CA; 4.6 × 250 mm, inner diameter, 5 μm) was prepared in the HPLC system. Chromatographic conditions used were as follows:
Rhodamine 123: mobile phase: methanol-water (70:30, v/v), column temperature: 40°C, flow rate: 1.0 mL/min, detection wavelength: 229 nm.
Vincristine: mobile phase: methanol-monopotassium phosphate solution (70:30, v/v), column temperature: 40°C, flow rate: 1.0 mL/min, detection wavelength: 279 nm.
Prazosin: mobile phase: methanol-monopotassium phosphate solution (52:48, v/v), column temperature: 40°C, flow rate: 1.0 mL/min, detection wavelength: 245 nm.
Ubenimex: mobile phase: methanol-water (63:37, v/v), column temperature: 40°C, flow rate: 1.0 mL/min, detection wavelength: 254 nm.
Glibenclamide: mobile phase: acetonitrile-water (58:42, v/v), column temperature: 30°C, flow rate: 1.0 mL/min, detection wavelength: 230 nm.
Cell Culture.
Caco-2 cells were used in this study. These cells were seeded on 6 cm culture plates and supplemented with 10% fetal bovine serum. Cultures were incubated at 37°C in a humidified 5% CO2 incubator. Caco-2 cells were plated at a density of 2 × 105 cells per well into 6-well plates or transwell plates. The protein and mRNA expressions of drug transporters were determined after the Caco-2 cells were exposed to 2% O2 for 24 hours (2%–24 hours), 10% O2 for 24 hours (10%–24 hours), 5% O2 for 24 hours (5%–24 hours), 5% O2 for 2 hours (5%–2 hours), 5% O2 for 6 hours (5%–6 hours), 5% O2 for 12 hours (5%–12 hours), 5% O2 for 24 hours (5%–24 hours), and 5% O2 for 48 hours (5%–48 hour)s. The 2% oxygen concentration was used for uptake and transport assays because the 2%–24h group had the most significant effect on the expression of MDR1, MRP2, BCRP, PEPT1, and OATP2B1.
Uptake and Transport Study.
The uptake and transport study was performed as described previously (Xiang et al., 2018). The Caco-2 cells were seeded at 1 × 105 cells/cm2 in 6-well plates for the uptake study and transwell polycarbonate inserts for transport study. Caco-2 cells were cultured for 14 days and 21 days for uptake and transport study, after seeding with the medium that was replaced every day.
For the uptake study, the culture medium was removed, and the cells were preincubated at 37°C for 20 minutes in 1 mL of Hank’s balanced salt solution (HBSS) after exposure to hypoxia. The medium was replaced, and the cells were incubated with 2 mL of HBSS solution containing rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide, for 30, 60, 90, and 120 minutes. The Caco-2 cells surface was quickly rinsed 3 times with ice-cold HBSS solution after incubation with substrates, and the Caco-2 cells were collected with a cell scraper, suspended in 1 mL of extraction solution and homogenized in an ultrasonicator. The concentration of substrates and protein was analyzed.
For the transport study, the reliability of the cell monolayer was assessed by measuring the transepithelial electrical resistance with a Millicell-ERS-electrode (Millipore, Billerica, MA). The Caco-2 cell monolayers were considered tight enough for the transport experiments when the transepithelial electrical resistance value was above 400 Ω·cm2. The Caco-2 cells were preincubated at 37°C for 20 minutes in 1 mL of HBSS after exposure to hypoxia. For influx direction, 0.5 mL substrates solution was added to the apical (AP) side, whereas 1.5 mL HBSS was added to the basolateral (BL) side. For efflux direction, 1.5 mL substrates solution was added to the BL side, and 0.5 mL HBSS was added to the AP side. Then, 100 μL sample was collected from receiver chambers every 30 minutes and immediately replaced with the same volume of fresh transport buffer until 120 minutes. The concentration of the substrates was determined by UPLC-MS (Ultra Performance Liquid Chromatography Mass Spectrometry) (Thermo Scientific Ultimate 3000 and Q-Exactive Focus MS, Sunnyvale, CA).

Where dC/dt (μmol/s) is the rate at which the compound appears in the receiver chamber, A (cm2) is the surface area of the transwell polycarbonate membrane (1.12 cm2), and C0 (μmol/L) is the initial concentration of drugs.
Sample Processing.
The HBSS solution sample was processed with acetonitrile to denature the protein, and the mixture of acetonitrile, internal standard (IS), and HBSS solution was vortexed for 0.5 minute and then centrifuged for 10 minutes at 11,400 g at 4°C. An aliquot of the supernatant was analyzed using LC-MS (Liquid Chromatography Mass Spectrometry) (Thermo Scientific Ultimate 3000 and Q-Exactive Focus MS, Sunnyvale, CA).
Chromatographic Conditions.
An HPLC separation module (Thermo Scientific UltiMate 3000, Sunnyvale, CA) with a cooled autosampler and column oven was used in this assay. Thermo Hypersil GOLD C18 50mm × 2.1mm, 1.9 μm column (Thermo Fisher, Sunnyvale, CA) was used for chromatographic separation. The gradient elution was performed as follows:
Ubenimex: flow rate: 0.3 mL/min, column temperature: 30°C.
Prazosin: flow rate: 0.3 mL/min, column temperature: 30°C.
Vincristine: flow rate: 0.3 mL/min, column temperature: 30°C.
Rhodamine 123: flow rate: 0.5 mL/min, column temperature: 40°C.
Glibenclamide: flow rate: 0.5 mL/min, column temperature: 40°C.
Mass Spectrometry Conditions.
A high-resolution mass spectrum (Q-Exactive Focus MS, Sunnyvale, CA) was used to analyze samples in positive electrospray ionization mode, and data were collected and processed using Trace Finder 4.0 software. Source parameters adopted were as follows: sheath gas flow rate: 35 arb; aux gas flow rate: 10 arb; sweep gas flow rate: 1 arb; spray voltage: 3.5 kV; capillary temp: 260°C; s-lens RF level: 50%; aux gas heater temp: 300°C. Rhodamine 123: 345.12337 (m/z), Rhodamine 6G (IS): 443.23292 (m/z), Vincristine: 825.40692 (m/z), Vinblastine (IS): 811.42766 (m/z), Prazosin: 384.16992 (m/z), Terazosin (IS): 387.19675 (m/z), Ubenimex: 309.18088 (m/z), Cilostazol (IS): 370.22375 (m/z), Glibenclamide: 494.15110 (m/z), and Glipizide (IS): 446.18565 (m/z).
Western Blot.
Total protein samples were harvested from liver tissue and homogenated, and Caco-2 cells were lysed by ice-cold lysis buffer followed by content determination. The protein samples were separated by electrophoresis on 10% SDS polyacrylamide gels, and then transferred to a polyvinylidene fluoride membrane. Tris-Buffered Saline Tween-20 was used to block membranes with 5% skimmed milk powder for 1.5 hours. The polyvinylidene fluoride membranes were incubated with the primary antibodies of β-Actin (1:500, mouse monoclonal, Abcam, ab8226), MDR1 (1:1000, rabbit monoclonal, Abcam, ab170904), MRP2 (1:1000, rabbit polyclonal, Sigma-Aldrich, M8316), BCRP (1:500, rabbit monoclonal, Abcam, ab207732), PEPT1 (1:500, rabbit polyclonal, Thermo Fisher, PA5-37010), and OATP2B1 (1:500, rabbit polyclonal, Novus, NBP-1-59811), respectively, at 4°C overnight. The membranes were incubated with a 1:10000 dilution of the second antibody at 37°C for 1 hour. Specific protein bands were detected by the ECL system (Amersham imager 600, Boston, MA).
Quantitative Real-time Polymerase Chain Reaction.
Liver tissue and Caco-2 cells were homogenized, and total RNA was isolated using TRIzol reagent. The quality of the RNA solutions was assessed using a NanoDrop 2000c spectrophotometer (Thermo Fisher Scientific). cDNA was synthesized according to the manufacturer’s protocol. Relative mRNA expression was analyzed using the BIO-RAD CFX Connect Real-Time PCR System (Bio-Rad Laboratories, Inc.). For mature let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148b-3p, and miR-148a-3p detection, reverse transcription was performed using poly(A)-tailed total RNA, a reverse transcription primer, and reverse transcriptase. Products were amplified at 95°C for 5 minutes, followed by 40 cycles at 95°C for 15 seconds, 60°C for 20 seconds, and 72°C for 40 seconds. Fold induction values were calculated according to the Eq. 2–ΔΔCt, where ΔCt represents the difference in cycle threshold numbers between the target gene and the control gene β-actin or U6, and ΔΔCt represents the relative change in the difference between the control and treatment groups. The primers are listed in Tables 2–4. The sequence of primer was: reverse 5′-AGTGCAGGGTCCGAGGTATT-3′ for all miRNAs. qPCR efficiency over all samples was calculated according to the Minimum Information for Publication of Quantitative Real-Time PCR Experiments guidelines.
Primer sequences corresponding to the rat genes examined with a real-time PCR analysis
Primer sequences corresponding to the Caco-2 cell genes examined with a real-time PCR analysis
Primer sequences corresponding to the cell miRNAs examined with a real-time PCR analysis
RNA Sequencing for Caco-2 Cells.
Briefly, total RNA was collected and lysed from Caco-2 cells with Trizol reagent after exposure to hypoxia or normoxia with Trizol reagent. The RNA quality was evaluated by a spectrophotometer (Thermo Fisher Scientific NanoDrop 2000c). A library for Illumina (HiSEq. 2500, San Diego, CA) platform was constructed consisting of miRNA enrichment, cDNA generation, and end repair to generate blunt ends, A-tailing, adaptor ligation, and PCR amplification. The differential miRNAs were considered upregulated if fold change was ≥2 and P-value was ≤ .05, and downregulated if fold change was ≤0.5 and P-value was ≤0.05.
miRNA Transfection.
The Caco-2 cells were plated at a density of 3 × 105 cells per well in 6-well plates and incubated at 37°C in a humidified 5% CO2 incubator for 24 hours. The Caco-2 cells were transfected with 769 nmol/L mimics or inhibitors of miR-873-5p or negative control (NC) RNA, as control using Lipofectamine 30EC00 and OPTI-MEM reduced serum medium according to the manufacturer's protocol. The miR-873-5p mimics and inhibitors were chemically synthesized by KeyGEN BioTECH.
Statistical Analysis.
All numerical data were expressed as mean ±SD. One-way analysis of variance (ANOVA) was used to analyze protein and mRNA expressions in rats and Caco-2 cells, followed by post hoc tests using least significant difference (LSD). Independent-samples t test was used for uptake and transport assay, and the difference of P < 0.05 was considered statistically significant. The rank-sum test was used for variance nonhomogeneity. All the statistical analyses were performed with the Statistical Package for the Social Sciences, version 13.0 (SPSS, Chicago, IL).
Results
Expression of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 in Rats.
As shown in Fig. 1A, the mRNA expression of MDR1 was significantly increased in the AMH and AHH groups, but decreased in the CHH group, compared with that in the P group. Compared with the P group, the mRNA expression of MRP2 was increased in the AMH and CHH groups, and mRNA expression of BCRP was increased in the AMH and AHH groups. Influx transporters also significantly changed in high-altitude hypoxia. The mRNA expression of PEPT1 was higher in the AMH, AHH, and CHH groups and lower in the CMH group, compared with that in the P group. High-altitude hypoxia did not affect the mRNA expression of OATP2B1.

Expression of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 in rats. (A) mRNA expression and (B) protein expression. Data were expressed as mean ±SD (n = 10). The data were analyzed using ANOVA, followed by posthoc tests using LSD tests. *P < 0.05 versus the P group. The P group (altitude: 390 m, PaO2: 20 kPa): rats living in the city of Xi’an. The AMH group (altitude: 2,800 m, PaO2: 15.1 kPa) and the CMH group ( altitude: 2,800 m, PaO2: 15.1 kPa): rats living at the city of Xi’an, but they received a 24-hour acute exposure and a 30-day chronic exposure to a middle-altitude area (Gonghe County). The AHH group (altitude: 4,300 m, PaO2: 12.4 kPa) and the CHH group (altitude: 4,300 m, PaO2: 12.4 kPa): rats living in the city of Xi’an, but the rats in the AHH group were acutely exposed for 24 hours to high altitudes, and the rats in the CHH group were chronically exposed for 30 days to high-altitude areas (Maduo County).
Rats received a 24-hour acute exposure and a 30-day chronic exposure to middle- and high-altitude levels. As shown in Fig. 1B, changes in MDR1 protein expression were in conformity to the corresponding changes in mRNA expression. The protein expression of MDR1 was significantly decreased in high-altitude hypoxia. MRP2 protein expression was decreased in the AMH group but increased in the CHH group. BCRP and PEPT1 protein expressions increased in high-altitude hypoxia, whereas the expression of OATP2B1 did not significantly change.
Functions of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 in Rats.
Figure 2 shows the absorption of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide in the intestine expressed in the form of Peff and Ka. Comparable data were obtained from these two parameters. The experiments were performed using several specific substrates. Rhodamine 123 is a specific substrate for MDR1, vincristine for MRP2, prazosin for BCRP, ubenimex for PEPT1, and glibenclamide is specific for OATP2B1. Our data showed that the Peff and Ka form of rhodamine 123 decreased when the rats were exposed to acute high-altitude hypoxia. Compared with the P group, the Peff and Ka form of vincristine in the AMH group significantly increased but decreased in the CMH and CHH groups. The Peff and Ka form of prazosin, ubenimex, and glibenclamide increased when the rats were exposed to hypoxia, but the Peff and Ka form of prazosin in the AHH group significantly decreased. The above results suggested that the activity of MDR1, PEPT1, and OATP2B1 increased under hypoxic conditions, and BCRP decreased in the CMH and CHH groups; however, it increased in the AHH group. The activity of MRP2 was found to be reduced after exposure to acute moderate altitude hypoxia but increased after exposure to chronic moderate- and high-altitude hypoxia.

Function of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 in rats. Left panel: intestinal absorption expressed in Peff (the effective permeability coefficient) of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide. Right panel: intestinal absorption expressed in Ka (the absorption rate constant) of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide. Data were expressed as mean ±SD (n = 6). The data were analyzed using ANOVA, followed by posthoc tests using LSD tests. *P < 0.05 versus the P group. The P group (altitude: 390 m, PaO2: 20 kPa): rats living in the city of Xi’an. The AMH group (altitude: 2,800 m, PaO2: 15.1 kPa) and the CMH group (altitude: 2,800 m, PaO2: 15.1 kPa): rats living in the city of Xi’an, but they received a 24-hour acute exposure and a 30-day chronic exposure to a middle-altitude area (Gonghe County). The AHH group (altitude: 4,300 m, PaO2: 12.4 kPa) and the CHH group (altitude: 4,300 m, PaO2: 12.4 kPa): rats living in the city of Xi’an, but the rats in the AHH group were acutely exposed to high altitudes for 24 hours, and the rats in the CHH group were chronically exposed to high altitudes areas for 30 days (Maduo County).
Expression of MDR1, MRP2, BCRP, PEPT1, OATP2B1, and PXR in Caco-2 Cells.
After Caco-2 cells were treated with 5% oxygen concentration for 2, 6, 12, 24, and 48 hours and 2%, 5%, and 10% oxygen concentration for 24 hours, protein and mRNA expressions were assessed by Western blot and qPCR. As shown in Fig. 3A, compared with the normoxia group, the mRNA expression of MDR1 in the 5%-2h group was increased, BCRP increased in the 10%-24h, 5%-2h, and 5%-6h group, PEPT1 increased in the 2%-24h group, but OATP2B1 decreased in the 5%-6h group. There was no significant difference in the mRNA expression of MRP2 under hypoxia. In addition to the 5%-2h group, the mRNA expression of PXR decreased in the 2%-24h, 10%-24h, 5%-6h, 5%-12h, 5%-24h, and 5%-48h group.

Expression of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 in Caco-2 cells. (A) mRNA expression and (B) protein expression. Normoxia: Caco-2 cells were cultured in a normoxic humidified incubator. Caco-2 cells were exposed to 2% O2 for 24 hours (2%-24 hour), 10% O2 for 24 hours (10%-24 hours), 5% O2 for 24 hours (5%-24 hours), 5% O2 for 2 hours (5%-2 hours), 5% O2 for 6 hours (5%-6 hours), 5% O2 for 12 hours (5%-12 hours), 5% O2 for 24 hours (5%-24 hours), and 5% O2 for 48 hours (5%-48 hours). Data were expressed as mean ± D (n = 3). The data were analyzed using ANOVA followed by posthoc tests using LSD tests. *P < 0.05 versus the normoxia group.
Figure 3B shows the protein expression of MDR1, PEPT1, and OATP2B1, which significantly increased in the 2%-24h, 10%-24h, 5%-2h, 5%-6h, 5%-12h, 5%-24h, and 5%-48h group, compared with the normoxia group. BCRP increased in the 2%-24h, 10%-24h, 5%-6h, 5%-12h, 5%-24h, and 5%-48h group. The protein expression of MRP2 and PXR significantly decreased in 2%-24h, 10%-24h, 5%-2h, 5%-6h, 5%-12h, 5%-24h, and 5%-48h group, compared with the normoxia group. We investigated the effect of time and oxygen concentration on the drug transporters after 24-hour treatment with 2% oxygen concentration, which had the greatest effect on MDR1, BCRP, PEPT1, and OATP2B1 protein expression. Therefore, 2% oxygen concentration was used for the subsequent uptake and transport experiment.
Effect of Hypoxia on the Uptake and Transport of Rhodamine 123, Vincristine, Prazosin, Ubenimex, and Glibenclamide.
Similar to the in vivo experiment, the functions of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 were evaluated by analyzing the uptake and transport of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide in vitro. As shown in Fig. 4, compared with the normoxia group, the uptake of rhodamine 123 significantly increased at 90 minutes, vincristine increased at 30, 60, and 90 minutes, and ubenimex significantly increased at 120 minutes. The uptake of glibenclamide decreased at 30 minutes but increased at 120 minutes. The uptake value did not significantly differ for prazosin. Figure 5 shows that the Papp of rhodamine 123 and ubenimex was significantly increased at 30, 60, 90, and 120 minutes under hypoxic conditions in the influx direction. The Papp of vincristine and glibenclamide significantly decreased at 30 minutes but increased at 60 minutes and 90 minutes under hypoxic conditions in the influx direction. The Papp of prazosin significantly decreased at 120 minutes under hypoxic conditions in the influx direction. Figure 6 shows that Papp of rhodamine 123 significantly increased at 30, 60, and 120 minutes, but vincristine decreased at 90 minutes under hypoxic conditions in the efflux direction. The Papp of prazosin and ubenimex significantly increased at 30, 60, 90, and 120 minutes under hypoxic conditions in the efflux direction. The Papp of glibenclamide significantly increased at 30 and 60 minute under hypoxic conditions in the efflux direction. The above-mentioned results suggest that the activity of efflux transporters MDR1 and BCRP increased, but the activity of MRP2 decreased under hypoxia. The activity of influx transporters PEPT1 was also found to be increased; however, the activity of OATP2B1 increased at 60 and 90 minutes and decreased at 30 minutes under hypoxia. Additionally, transport from AP to BL of rhodamin123 as well as transport from BL to AP of ubenimex was also increased by hypoxia, which suggested that the activity of influx transporters for rhodamin123 and efflux transporters for ubenimex also increased under hypoxia.

Effect of hypoxia on the uptake of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide in Caco-2 cell model. Caco-2 cells were incubated at 37°C with 2% O2 for 24 hours in the 6-well plates, and then the samples were collected at different time points within 120 minutes. Data were expressed as mean ±SD (n = 3). The data were analyzed using an independent sample t test. *P < 0.05 versus the normoxia group.

Effect of hypoxia on the transport (AP-BL) of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide in Caco-2 cell model. (A) rhodamine 123; (B) vincristine; (C) prazosin; (D) ubenimex; and (E) glibenclamide. Caco-2 cells were incubated at 37°C with 2% O2 for 24 hours in the apical or basolateral side of the transwell insert, and then the samples were collected at different time points within 120 minutes. AP-BL: apical-to-basolateral direction transport. Data were expressed as mean ±SD (n = 3). The data were analyzed using an independent sample t test. *P < 0.05 versus the normoxia group.

Effect of hypoxia on the transport (BL-AP) of rhodamine 123, vincristine, prazosin, ubenimex, and glibenclamide in Caco-2 cell model. (A) rhodamine 123; (B) vincristine; (C) prazosin; (D) ubenimex; and (E) glibenclamide. Caco-2 cells were incubated at 37°C with 2% O2 for 24 hours in the apical or basolateral side of the transwell insert, and then the samples were collected at different time points within 120 minutes. BL-AP: basolateral-to-apical direction transport. Data were expressed as mean ±SD (n = 3). The data were analyzed using an independent sample t test. *P < 0.05 versus the normoxia group.
Effect of Hypoxia on miRNAs’ Expression Profile in Caco-2 Cells.
In a systematic approach to identifying miRNAs involved in hypoxia, we applied the well-established model of hypoxia treatment in Caco-2 cells. We then applied sequencing analysis to compare miRNA expression levels between hypoxia and normoxia and found that 27 miRNAs were upregulated and 56 miRNAs were downregulated in the hypoxic conditions (Fig. 7).

Expression profiles of miRNA under hypoxic conditions in Caco-2 cells. (A) microarray heat map; (B) volcano plot; and (C) scatter diagram. Caco-2 cells were incubated at 37°C with 2% O2 for 24 hours in 6-well plates. The expression levels of miRNAs are shown using the indicated pseudocolor scale from -10 (green) to 10 (red) relative to the value for Caco-2 cells in the normoxia group. The color represents the relative mRNA expression levels, green indicating downregulated miRNAs, red indicating upregulated miRNAs, and black indicating unchanged miRNAs.
miR-873-5p May Play a Key Role in the Regulation of MDR1 and PXR under Hypoxia.
let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148b-3p, and miR-148a-3p were regarded as possible regulators when MDR1 and PXR changed under hypoxia by using Miranda, a microRNA target prediction database, in which we found binding sites of let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148b-3p, and miR-148a-3p in PXR 3′ UTR. As shown in Fig. 8, the mRNA expression of let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148b-3p, and miR-148a-3p significantly decreased in hypoxic conditions. According to the literature reports (Wu et al., 2016), miR-873-5p can regulate MDR1. Therefore, miR-873-5p was selected for the transfection experiment.

The expression of let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148b-3p, and miR-148a-3p under hypoxia. Caco-2 cells were incubated at 37°C with 2% O2 for 24 hours in 6-well plates. The expressions of let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148b-3p, and miR-148a-3p were detected by qPCR. Data were expressed as mean ±SD (n = 3). The data were analyzed using an independent sample t test. *P < 0.05 versus the control group.
Identification of miR-873-5p that Regulates the Expression of MDR1 and PXR.
To investigate whether miR-873-5p has a function in the downregulation of MDR1 and PXR expression, Caco-2 cells were cultured in a medium and then cells were transfected with mimics and inhibitors of miR-873-5p or NC. Figure 9 shows that miR-873-5p expression significantly increased in the mimic group but decreased in the inhibitor group. The expressions of mRNA and protein of MDR1 and PXR increased after the treatment of Caco-2 cells with an inhibitor of miR-873-5p and decreased in the mimic group.

miR-873-5p regulates the expression of MDR1 and PXR in Caco-2 cells. Caco-2 cells were transfected with NC RNA, mimics, and inhibitors of miR-873-5p. The mRNA expression of (A) miR-873-5p, (B) MDR1, and (C) PXR was determined by qPCR. (D) Protein expression of MDR1 and PXR was detected by Western blot. Data were expressed as mean ±SD (n = 3). The data were analyzed using ANOVA followed by posthoc tests using LSD tests. *P < 0.05 versus the control group.
Discussion
Drug metabolizing enzymes and transporters are the main factors affecting drug metabolism. In our previous studies, we found that high-altitude hypoxia affects the activity and expression of cytochrome P450 (Zhou et al., 2018; Duan et al., 2020; Duan et al., 2021). To date, only a few reports have addressed the ways to resolve the problem of changes in the functions and expression levels of drug transporters under high-altitude hypoxic environments. To further study the effects of high-altitude hypoxia on drug transporters, changes in the functions and expression levels of MDR1, MRP2, BCRP, PEPT1, and OATP2B1 were examined in rats and Caco-2 cells. The results suggested that high-altitude hypoxia increased the expression and activity of most drug transporters.
Fradette et al. found that hypoxia increased the protein and mRNA expression of P-gp, but hypoxia did not affect the mRNA expression of BCRP in rats subjected to a Fio2 of 8% for 48 hours compared with those in rabbits breathing 21% Fio2 (Fradette et al., 2007). Lazarowski et al. showed that hypoxia induced neuronal MDR1 expression after experimental focal hypoxia by local injection of CoCl2 into the frontoparietal cortex in male adult rats (Lazarowski et al., 2007). Zhang et al. found that MDR1 was significantly changed and directly regulated by Kruppel Like Factor 8 under hypoxia (Zhang et al., 2014), which is consistent with our findings. Another study found that the mRNA expression of MDR1 was decreased by hypoxia, and the Peff value of the substrate was significantly increased (Li et al., 2016). Models of hypoxia that are induced by low-pressure cabins or chemical methods can only simulate hypoxia, and these methods cannot simulate the conditions of hypothermia, dry climate, or intense radiation, which means these methods cannot reflect a real high-altitude environment.
The structure and function of Caco-2 cells are similar to intestinal epithelial cells, and they share the same results in studies concerning in vivo and in vitro drug transporter functions. Efflux transporters, such as MDR1, MRP2, and BCRP, play the role of an intestinal barrier in the small intestine, discharging intracellular substances into the intestinal cavity, whereas uptake transporters, such as PEPT1 and OATP2B1, are widely expressed in the small intestine and are capable of promoting the absorption of drugs (Liu, 2019). Under the influence of hypoxic conditions, the improved efflux functions of MDR1 and BCRP may weaken drug absorption and cause drug resistance, whereas the enhanced uptake functions of PEPT1 and OATP2B1 may promote drug absorption as well as improve bioavailability. The significantly decreased efflux function of MRP2 may reduce the occurrence of multidrug resistance.
Studies have reported an association between the uptake activity of PEPT1 and pH. If the pH of the drugs that are absorbed in the small intestine can be adjusted to the most appropriate range of PEPT1, their bioavailability can be improved (Wu et al., 2020). Firstly, the structure of intestinal microbiota changed significantly under hypoxic conditions, and intestinal pH was also changed accordingly (Liu et al., 2021), which significantly affected the uptake activity of PEPT1. Secondly, the results of this study showed that different oxygen concentrations had different effects on the protein expression of PEPT1. The 2%-24h group had the most significant effect, followed by 5%-48h, 5%-24h, 5%-12h, and 5%-6h groups. If the optimal oxygen concentration and the optimal pH could be found, the bioavailability of PEPT1 substrate drugs could be significantly improved, and the problem of the low absorption rate of some oral drugs could be solved. This experiment only studied the effects of different oxygen concentrations on the protein expression of transporters and not on the functions of transporters, which will be covered in subsequent experiments.
The expression regulation of genes is a complex process that comprises transcriptional regulation and posttranscriptional regulation. The former is further divided into DNA regulation at the genetic level and epigenetic regulation, whereas the latter is further divided into the regulation of RNA processing and maturation process, translational regulation, and regulation of protein processing (Zhu et al., 2012). In this experiment, the proteins of MDR1, MRP2, and OATP2B1 have different variation trends of expression from those of mRNA, probably due to posttranscriptional regulation.
Bioinformatics analysis showed that multiple miRNAs could target the PXR gene, and PXR is an important member of the nuclear receptor family. Studies have shown a strong link between PXR and MDR1 (Gu et al., 2013; Li et al., 2017). Previous studies by the authors have demonstrated a significant reduction of PXR expression in rats and HepG2 cells under hypoxic conditions (Duan et al., 2020). This study showed that the expression of PXR in Caco-2 cells under hypoxic conditions was also reduced significantly, which is in line with the previous studies. It is important to study the significance of the relationship of the miRNA-PXR-MDR1 axis.
A high-throughput sequencing technique was adopted to detect the changes of miRNAs under hypoxic conditions. Eighty-three miRNAs were observed to be changed significantly, of which the expression of 27 miRNAs was found to be increased, and the expression of 56 was found to be decreased. We conducted target predictions on those miRNAs using miRanda and selected let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148a-3p, and miR-148b-3p for their potentials to interact with PXR. qPCR was then used to demonstrate the expression reduction of let-7f-5p, let-7g-5p, miR-522-3p, miR-873-5p, miR-148a-3p, and miR-148b-3p under hypoxic condition. Wu et al. (Wu et al., 2016) investigated the multidrug resistance mechanism in ovarian cancer cells and found that miR-873 mediated multidrug resistance by targeting MDR1, confirming that miR-873 could regulate the expression of MDR1. Takagi et al. (Takagi et al., 2008) confirmed that miR-148a has a regulatory effect on PXR. A drug resistance study of HT29 colon cancer cells found that miR-522 could target ABCB5 to reverse amycin-induced drug resistance (Yang et al., 2015). The regulatory effects of let-7f, let-7g, miR-522, and miR-148 on MDR1 have not been reported yet. According to literature reports and bioinformatics analysis results, we found that miR-873-5p might target MDR1 and PXR to regulate drug metabolism.miR-873-5p-mimic and inhibitor transfection cells were used to verify the regulatory effects of miR-873-5p on PXR and MDR1. It was found that PXR expression significantly decreased in miR-873-5p overexpression cells and significantly increased in miR-873-5p low-expression cells, confirming that miR-873-5p could negatively regulate PXR expression. Numerous studies have confirmed that the dimer of the nuclear receptor PXR and its molecular chaperone directly bind the DNA to MDR1 or recruit other DNA transcription factors to activate the genetic transcription of MDR1 (Gu et al., 2013; Feng et al., 2018; Moscovitz et al., 2018). According to the above-mentioned results, this study speculates that miR-873-5p may target PXR to regulate the expression of MDR1. However, Fig. 3 shows the opposite trend as overexpression of MDR1 occurs along with downregulation of PXR. It is suggested that the progress of drug transporters regulation under hypoxia might involve several signaling pathways and not a single signaling pathway; this is an integral component of gene regulatory networks and requires further study. In miR-873-5p overexpression and low-expression cells, MDR1 expression was also significantly decreased and increased, respectively, confirming that miR-873-5p also negatively regulated MDR1 expression in a direct manner. Besides, in the family of nuclear receptors, constitutive androstane receptor (CAR) could regulate MRPs (Lv and Huang, 2020), PXR could regulate MRPs (Mazerska et al., 2016), farnesoid X receptor (FXR) could regulate OATPs (Malagnino et al., 2019), and hepatocyte nuclear factor 4-alpha (HNF4α) culd regulate the expression of MDR1 (Vagnerová et al., 2019). Furthermore, nuclear receptors could also interact with each other. For example, FXR and peroxisome proliferator-activated receptor could interact (Li et al., 2020), and HNF4α could regulate the expression of PXR, CAR, and hepatocyte nuclear factor 1-alpha (Pavek, 2016; Tian et al., 2018; Rosenthal et al., 2019); such an interaction greatly complicates the regulation of drug transporters. Apart from the regulatory effect of miR-873-5p on MDR1 confirmed by this study, miR-27a, miR-145, miR-491, etc., could all act on MDR1 (Zhao et al., 2017; Kong et al., 2019; Shi et al., 2020). It was observed that miRNA had a regulatory effect on CAR, and it is also interesting to note that CAR could, in turn, downregulate the expression of miRNA, forming a two-way regulation (Kazantseva et al., 2015). The regulation of multiple miRNAs on transporters and nuclear receptors, the interactions among nuclear receptors, the regulation of nuclear receptors on transporters, and the negative feedback of nuclear receptors on miRNAs constitute a multifactor, multitarget, interactive mechanism of drug transporters regulation, with miRNA being the key regulatory factor.
In vivo experiments have confirmed that the transport function of MDR1 in small intestines of rats significantly increased under high-altitude hypoxic environment. In vitro experiments have proven that the protein and mRNA expressions of MDR1 significantly increased and the efflux activity improved considerably under hypoxic conditions. The results mentioned above suggest that hypoxia is an inducer of MDR1 and may lead to drug interactions. Tumor cells are also a microhypoxic environment and an important reason for the occurrence of multidrug resistance. The increase of MDR1 under high-altitude hypoxic conditions may also enhance the incidence of multidrug resistance and reduce the efficacy. Therefore, high-altitude hypoxia may not only affect the physiologic disposition of drugs but also facilitate interactions between drugs and increase the incidence of multidrug resistance.
This experiment determined MDR1 protein and mRNA expressions in miR-873-5p overexpression and low-expression Caco-2 cells, respectively, but did not determine the transporter performance of MDR1. Our future research will focus on establishing miR-873-5p overexpression and low-expression Caco-2 monolayer models under hypoxic conditions, and transport experiments will also be conducted on MDR1 substrate drugs to confirm further whether miR-873-5p targets MDR1 to regulate the metabolic process of drugs. MDR1 is a key drug transporter in the liver, kidneys, and intestine. It can pump intracellular drugs out of cells with the help of adenosine triphosphate, thereby reducing the concentration of intracellular drugs (Zhang et al., 2020). However, MRP2, BCRP, PEPT1, and OATP2B1 also play a key role in regulating the physiologic disposition of drugs under hypoxic conditions. Whether miRNA target MRP2, BCRP, PEPT1, and OATP2B1 is still unclear, which needs further study.
The results of this study may be helpful for rational pharmaceutical use in high-altitude environments. It is suggested that patients should be closely monitored when they are living in or traveling to high-altitude areas, and the dosages of some drugs that are transported by MDR1, MRP2, BCRP, PEPT1, and OATP2B1 should be reevaluated by therapeutic drug monitoring.
Authorship Contributions
Participated in research design: Duan, Bai, Li.
Conducted experiments: Duan, Bai, Yang, Zhou, Gu, Zhu, La.
Contributed new reagents or analytic tools: Liu, Wang.
Performed data analysis: Duan, Li.
Wrote or contributed to the writing of the manuscript: Duan, Bai, Li.
Footnotes
- Received September 17, 2021.
- Accepted November 12, 2021.
↵1 Y.D. and X.B. contributed equally to this work.
This work was supported by the National Natural Science Foundation of China [Grant 81760673 and 81460568], the Qinghai Innovation Platform Construction Project [Grant 2021-ZJ-T03], and the Key Research Foundation of Development and Transformation of Qinghai Province [Grant 2021-SF-158].
No author has an actual or perceived conflict of interest with the contents of this article.
Abbreviations
- ADME
- absorption, distribution, metabolism, and excretion
- AHH
- acute high-altitude hypoxia
- AMH
- acute moderate-altitude hypoxia
- AP
- apical
- BCRP
- breast cancer resistance protein
- BL
- basolateral
- Caco-2
- colon cancer
- CAR
- constitutive androstane receptor
- CHH
- chronic high-altitude hypoxia
- CMH
- chronic moderate-altitude hypoxia
- CYP450
- cytochrome P450
- HBSS
- Hank’s balanced salt solution
- HPLC
- high-performance liquid chromatography system
- IS
- internal standard
- LSD
- least significant difference
- MDR1
- multidrug resistance protein 1
- miRNA
- microRNA
- MRP2
- multidrug resistance-associated protein 2
- NC
- negative control
- OATP2B1
- organic anion-transporting polypeptides 2B1
- P
- plain
- PEPT1
- peptide transport 1
- PXR
- pregnane X receptor
- Copyright © 2022 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}