


Abstract
Corynoline is a 1,3-benzodioxole-containing isoquinoline alkaloid isolated from Corydalis bugeana Turcz., a traditional herbal medicine. Corynoline has reportedly demonstrated multiple pharmacologic properties. Previous studies have also shown that corynoline induced cytotoxicity and inhibited cytochrome P450 (CYP) enzymes, but the mechanisms of the adverse effects remain unknown. The major objective of the present study was to identify reactive metabolites of corynoline responsible for the cytotoxicity and enzyme inhibition. Three oxidative metabolites (M1–M3) were detected by liquid chromatography–tandem mass spectrometry in rat liver microsomal incubations after exposure to corynoline. M1 and M2 were two isomers of catechol derivatives, and M3 was a di-catechol. The M1–M3 metabolites were also observed in urine of rats given corynoline. A total of four N-acetylcysteine (NAC) conjugates (M4–M7) were detected in microsomes containing corynoline, NAC, and NADPH. Apparently, M4 and M5 were derived from M1, M6 resulted from M2, and M7 was a M3-derived NAC conjugate. This indicates that corynoline was bioactivated to ortho-quinone derivatives. No corynoline-derived NAC conjugates (M4–M7) were detected in urine of rats given corynoline; however, three corresponding cysteinylglycine conjugates (M8–M10) were observed instead. Recombinant P450 enzyme incubations demonstrated that the CYPs 2C9, 3A4, and 2C19 were mainly involved in metabolic activation of corynoline. The metabolism study facilitates the understanding of corynoline-induced cytotoxicity and P450 enzyme inhibition.
Introduction
Corydalis bugeana Turcz., a perennial herb, belongs to Corydalis plants of the Papaveraceae family (Xie et al., 2004). The dried whole plant is referred to in traditional herbal medicine as C. Bungeanae Herba (Kudiding in Chinese) and is officially included in the 2010 version of the Chinese Pharmacopoeia. Kudiding has been widely used in China and other Asian nations for the treatment of various infectious diseases. The chemical components found in Kudiding include alkaloids, steroids, and flavonoid glycosides (Yang et al., 2014). Isoquinoline alkaloids are reportedly the major components occurring in the herb (Niu et al., 2011). Corynoline (Fig. 1) is one of the isoquinoline alkaloids and was first isolated and characterized from Papaveraceae in 1968 (Naruto et al., 1968). The alkaloid has been proven to show potent anti-inflammatory and/or immunosuppressive activity (Kamigauchi et al., 2005). A recent study demonstrated that corynoline was an inhibitor of prolyl oligopeptidase, which is reportedly associated with schizophrenia, bipolar affective disorder, and related neuropsychiatric disorders (Cahlíková et al., 2015). Despite this, corynoline was found to show time-dependent inhibition of CYP3A4 (Fang et al., 2011), and it has been reported to be toxic to cultured human A549, SK-OV-3, SK-MEL-2, and HCT15 cells (Choi et al., 2007). The observed enzyme inhibition and cytotoxicity raise the concern on the safe use of corynoline and its remedies.

Structure of corynoline.
The toxicities of many xenobiotics are mediated by an initial cytochrome P450 (CYP)–dependent generation of reactive electrophilic intermediates that react with nucleophilic sites of macromolecules, such as proteins and nucleic acids. The modification of the biomolecules is suggested to trigger the process of toxicities. Many structural toxicophores have been documented based on structure-toxicity relationship studies (Kalgutkar and Soglia, 2005; Baillie, 2006), which enables us to avoid these functional groups in the practice of designing drugs. 1,3-Benzodioxole group is such a structure generally considered as an important structural alert. Many 1,3-benzodioxole-containing compounds have been reported to induce P450-dependent toxicities and P450 enzyme inactivation. The P450-mediated metabolic activation generally starts with O-demethylenation of those 1,3-benzodioxole-containing compounds leading to ring opening and formation of a catechol metabolite (Hutzler et al., 2006), followed by oxidation to produce electrophilic ortho-quinones (Hutzler et al., 2008). For example, the generation of an ortho-quinone metabolite derived from 3,4-methylenedioxymethamphetamine is reportedly responsible for toxicities induced by 3,4-methylenedioxymethamphetamine (Tucker et al., 1994; Antolino-Lobo et al., 2010). Sitaxentan, a selective endothelin-A receptor antagonist, was withdrawn from the global market by Pfizer in December 2010 due to its idiosyncratic hepatotoxicity. A similar ortho-quinone metabolite of sitaxentan was recently characterized and was suggested to be associated with the reported hepatic toxicity (Erve et al., 2013).
Corynoline contains two 1,3-benzodioxole functional groups located in the two sides of the structure. The reported cytotoxicity of corynoline and CYP3A4 inactivation led us to seek electrophilic reactive metabolites of corynoline. The objectives of the present study were 1) characterization of reactive electrophilic metabolites of corynoline using in vitro and in vivo models; 2) elucidation of the pathways of corynoline bioactivation; and 3) identification of cytochrome P450 enzymes responsible for metabolic activation of corynoline.
Materials and Methods
Chemicals and Materials.
Corynoline with purity of >98% was obtained from Chengdu Pufeide Biologic Technology Co., Ltd. (Chengdu, China). N-Acetylcysteine (NAC) and NADPH were purchased from Sigma-Aldrich (St. Louis, MO). Recombinant human P450 enzymes were acquired from BD Gentest (Woburn, MA). Rat liver microsomes (Sprague-Dawley, male) were prepared in our laboratory according to a previously published method (Lin et al., 2014). Diazepam was purchased from the National Institute for the Control of Pharmaceutical and Biologic Products (Shenyang, China). All organic solvents were purchased from Fisher Scientific (Springfield, NJ). Distilled water was purchased from Wahaha Co. Ltd. (Hangzhou, China). All of the reagents and solvents were analytical or high-performance liquid chromatography grade.
Microsomal Incubations.
Corynoline (62.5 μM) and rat liver microsomes (4.0 mg/protein/ml) were mixed in potassium phosphate buffer (pH 7.4), followed by addition of NAC (10 mM) dissolved in phosphate buffer as a trapping agent. The total volume of the incubation mixtures was 200 μl. The reactions were initiated by addition of NADPH (1.0 mM). The negative control without NADPH was included to ensure that the metabolism of corynoline was NADPH dependent. The reaction mixtures were incubated at 37°C in a gentle shaking water bath for 60 minutes.
Recombinant Human P450 Incubations.
Corynoline was individually incubated with human recombinant CYPs (100 nM), including the CYPs 1A2, 2B6, 2C9, 2C19, 2D6, 2E1, 3A4, and 3A5. Other conditions were equivalent to the microsomal incubation experiments. A total normalized rate method was applied. The rates of metabolite formation in individual incubations with recombinant P450 enzymes were multiplied by the mean specific content of the corresponding P450 enzyme in human liver microsomes to obtain the normalized reaction rates of each enzyme.
Sample Preparation for Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS) Analyses.
The incubation reactions were terminated by mixing with double volumes of ice-cold acetonitrile, followed by addition of diazepam (1.25 nM). The mixtures were vortexed for 3 minutes and centrifuged at 16,000 rpm for 10 minutes at 4°C. The resulting supernatants (1 μl) were injected into LC-MS/MS for analyses.
Animal Studies.
Male Sprague-Dawley rats (200 ± 20 g) were obtained from the Animal Center of Shenyang Pharmaceutical University. All animal manipulations met the requirements approved by the Ethics Review Committee for Animal Experimentation of Shenyang Pharmaceutical University (Shenyang, China). All rats were kept in a controlled environment (room temperature and moderate humidity) under 12 hour dark/light cycles and fed on commercial rat chow and water ad libitum. Rats were individually placed in metabolism cages. Blank urine samples were collected after a 12-hour overnight fast with free access to water prior to the experiment. Rats were i.p. administered with corynoline dissolved in 1,2-propanediol/saline (1:4, v/v) at 20 mg/kg. Urine samples were collected during the time periods of 0–12, 12–24, and 24–48 hours after administration. During the experiments, the rats were allowed unlimited access to food and water. The collected urine samples were stored at −80°C until analysis.
Preparation of Urine Samples for LC-MS/MS.
Triple volumes of acetonitrile were added to the urine samples. Each sample was vortexed for 3 minutes and then centrifuged at 16,000 rpm for 10 minutes at 4°C. The supernatants were harvested and blown dry under a nitrogen stream at 40°C. The resulting samples were reconstituted with 100 μl acetonitrile/water (50/50, v/v). After centrifugation, the supernatants (1 μl) were analyzed by LC-MS/MS.
Chemical Synthesis of Corynoline NAC Conjugates.
Chemical demethylenation of corynoline was performed, based on a published protocol (Cano et al., 2013). Briefly, corynoline (2 mg) and AlCl3 (2.2 mg) were mixed in 4 ml of toluene. The mixture was refluxed for 20 hours at 110°C. After cooling down to room temperature, 0.5 ml water was added, followed by evaporation of the organic solvent under vacuum. The resulting solid was dissolved in CH2Cl2 (4 ml) and mixed with 2 mg 2,3-dichloro-5,6-dicyano-1,4-benzoquinone. The mixture was stirred at 4°C for 4 hours. After evaporation of the solvent, the crude products were dissolved in 4 ml H2O and mixed with NAC (2 mg). The mixture was stirred at room temperature for 10 hours. The resulting reaction solution was centrifuged at 16,000 rpm for 5 minutes, and the supernatants (1 μl) were subjected to LC-MS/MS for analysis.
LC-MS/MS Analysis Method.
Identification of metabolites was conducted on an AB SCIEX Instruments 4000 hybrid triple quadrupole-liner ion trap (Q-Trap) mass spectrometer (Applied Biosystems, Foster City, CA) equipped online with an ekspert ultraLC 100 system (Applied Biosystems). Chromatographic separation was achieved on an Ultimate XB-C18 column (2.1 × 100 mm, 3 μm) (Welch Scientific, Inc., Shanghai, China). Gradient elution was performed with acetonitrile containing 0.1% formic acid (solvent A) and water containing 0.1% formic acid (solvent B) at a constant flow rate of 0.4 ml/min. The volume of injection was 1 μl. Analyses were carried out within 14 minutes under the following gradients: 0–2 minutes, 5% A; 2–10 minutes, 5%–80% A; 10 to 11 minutes, 80%–90% A; 11–13 minutes, 90%–5% A; and 13 to 14 minutes, 5% A. Mass spectrometric analyses were conducted using an electrospray ionization source in positive-ion mode. The 4000 Q-Trap system was operated with a turbo ion spray using the following conditions: gas 1, 50 psi; gas 2, 50 psi; curtain gas (N2), 25 psi; heater temperature, 650°C; ion spray voltage, 5500 V; declustering potential, 70 V; entrance potential, 10 V, and cell exit potential, 3 V. For targeted analysis, multiple-reaction monitoring (MRM) and enhanced product ion (EPI) scanning were used to selectively monitor with ion transitions m/z 356 → 289 and 344 → 277 (oxidative metabolites), m/z 517 → 388 and 505 → 376 (NAC conjugates), and m/z 285 → 193 (diazepam, internal standard), respectively. The collision energy was set at 50 eV with a spread of 3 eV. All data were analyzed using the AB SCIEX Analyst 1.6.1 software (Applied Biosystems).
Results
Mass Spectrometric Behavior of Corynoline.
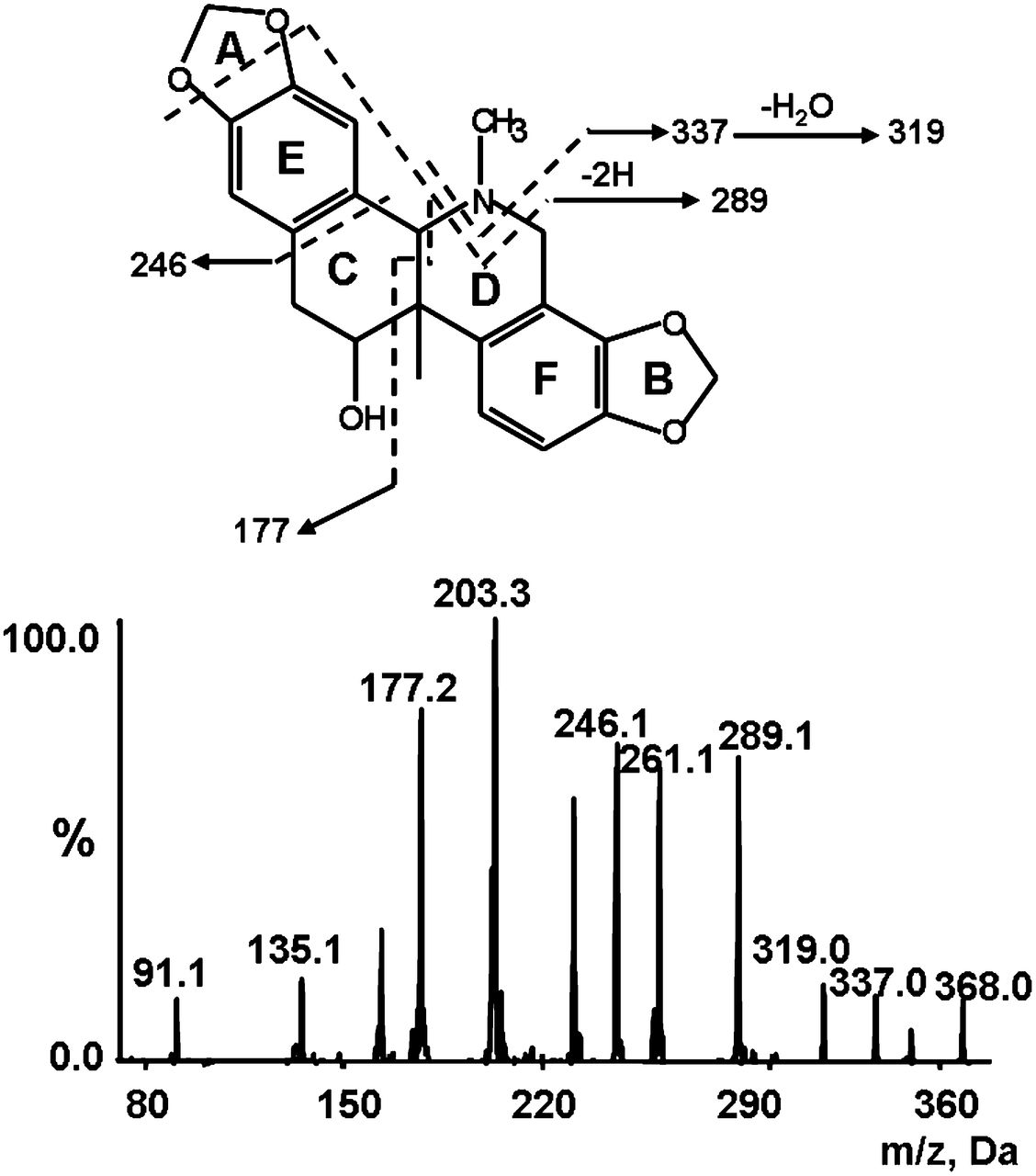
To facilitate metabolite identification, we started with mass spectrometric analysis of the parent compound corynoline. Corynoline was analyzed in positive ionization mode due to higher sensitivity and ionization efficiency and more fragment ions obtained. The parent drug displayed fragment ions of m/z 350, 337, 319, 289, 246, and 177 with [M+H]+ of m/z 368 (Fig. 2), which we proposed were derived from a loss of H2O, CH5N, CH7ON, C2H9O2N, C7H6O2, and C11H13O2N, respectively. These fragment ions represented integrated benzodioxole moieties (A and B rings, Fig. 2).

MS/MS spectrum of corynoline.
In Vitro Metabolic Activation of Corynoline.
Three oxidative metabolites (M1–M3) and four NAC conjugates (M4–M7) were detected in microsomal incubations containing corynoline, NADPH, and NAC (Scheme 1). No such metabolites were observed in the control group, which lacked NADPH (Fig. 3A; Fig. 4A; Fig. 5A), indicating that the generation of the metabolites was NADPH dependent.

Proposed pathways for the formation of corynoline-drived ortho-quinones by P450-mediated oxidation and synthetic reactions of corynoline.

Identification of metabolites M1, M2, and M3. Extracted ion (m/z 356/289 for M1 and M2, and 344/277 for M3) chromatograms obtained from LC-Q-Trap/MS analysis of rat liver microsome incubations containing corynoline and NAC in the absence (A) or presence (B) of NADPH. (C) Extracted ion chromatograms obtained from LC-Q-Trap/MS analysis of synthetic M1, M2, and M3. (D) Extracted ion chromatograms of M1, M2, and M3 obtained from LC-Q-Trap/MS analysis of urine samples of rats treated with corynoline. MS/MS spectra of M1 (E), M2 (F), and M3 (G) generated in microsomal incubations. MS/MS spectra of synthetic M1 (H), M2 (I), and M3 (J). MS/MS spectra of M1 (K), M2 (L), and M3 (M) in the urine samples of rats given corynoline.
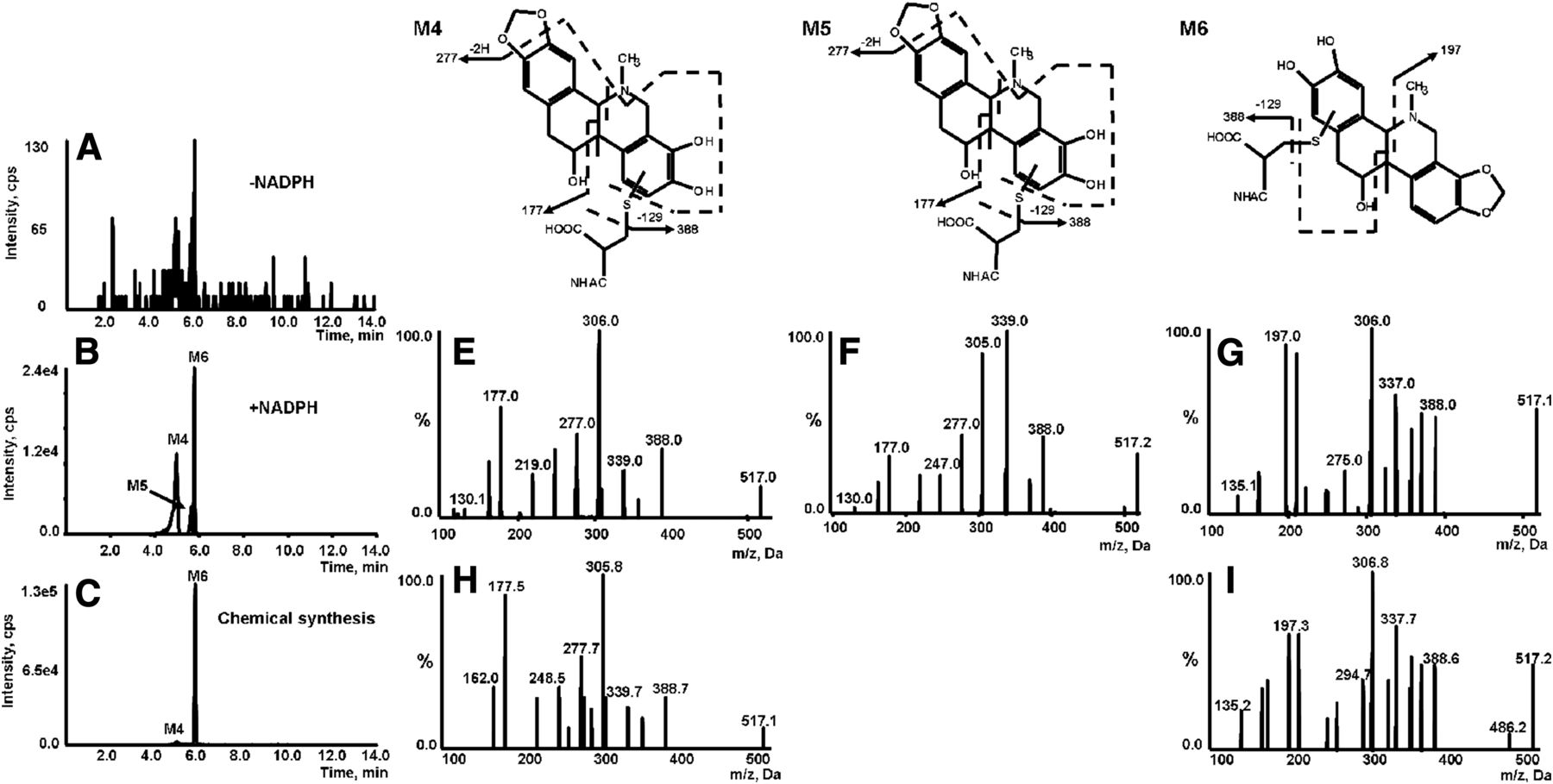
Identification of metabolites M4, M5, and M6. Extracted ion (m/z 517/388 for M4, M5, and M6) chromatograms obtained from LC-Q-Trap/MS analysis of rat liver microsome incubations containing corynoline and NAC in the absence (A) or presence (B) of NADPH. (C) Extracted ion chromatograms obtained from LC-Q-Trap/MS analysis of synthetic M4 and M6. MS/MS spectra of M4 (E), M5 (F), and M6 (G) generated in microsomal incubations. MS/MS spectra of synthetic M4 (H) and M6 (I).

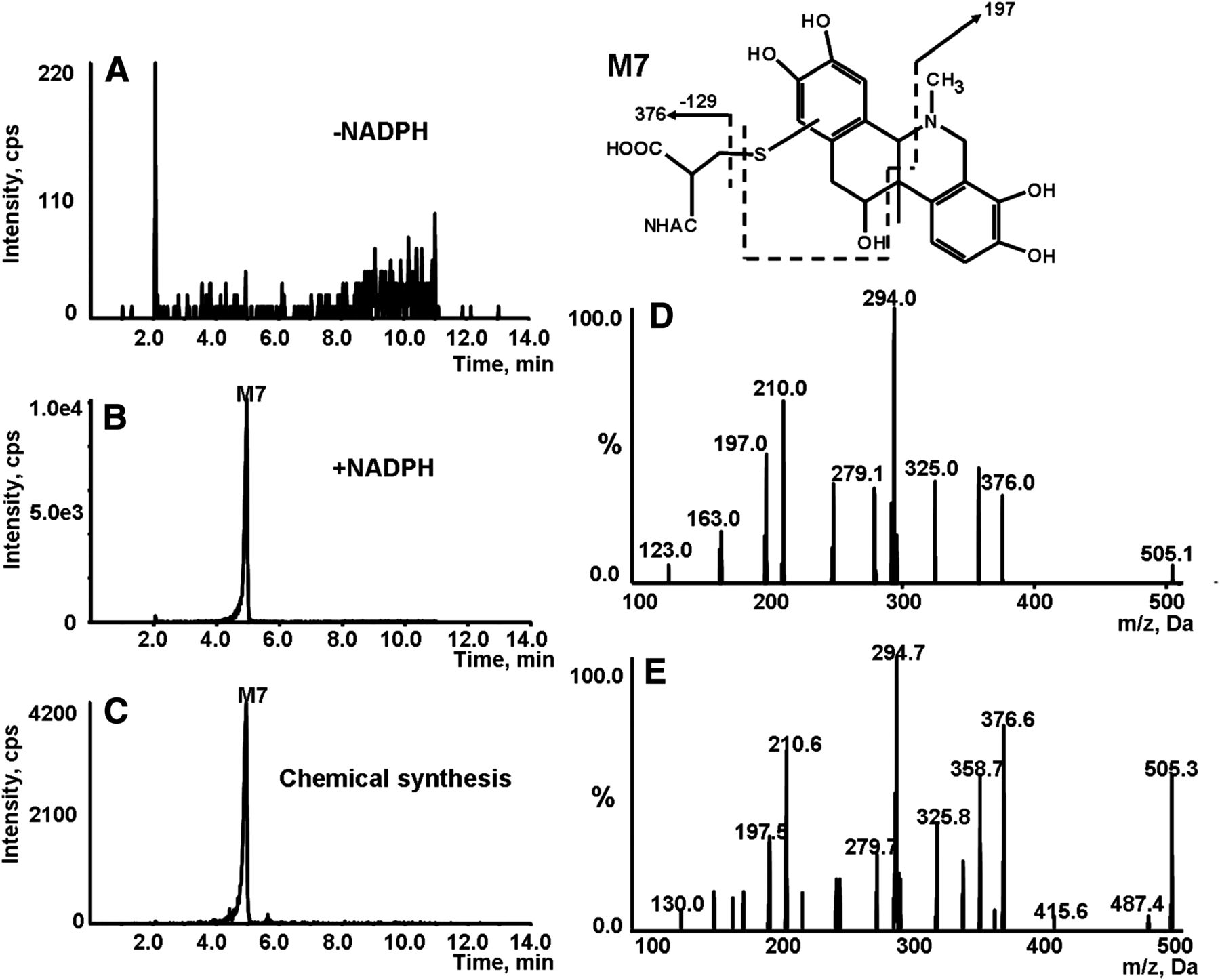
Identification of metabolite M7. Extracted ion (m/z 505/376 for M7) chromatogram obtained from LC-Q-Trap/MS analysis of microsomal incubations containing NAC in the absence (A) or presence (B) of NADPH. (C) Extracted ion chromatogram obtained from LC-Q-Trap/MS analysis of synthetic M7. (D) MS/MS spectrum of M7 generated in microsomal incubations. (E) MS/MS spectrum of synthetic M7.
M1 and M2 eluted out at 5.52 and 5.80 minutes showed their molecular ions at m/z 356, 12 Da lower than m/z 368 of [M+H]+ of the parent compound, indicating the demethylenation on one of the 1,3-benzodioxole rings of corynoline. The MS/MS spectra of M1 and M2 were obtained by MRM-EPI scanning (ion transition m/z 356/289) (Fig. 3, E and F). The product ions of M1 and M2 at m/z 338 were derived from the loss of H2O. M1 displayed characteristic fragment ions of m/z 307, 277, and 234, resulting from a loss of CH7ON, C2H9O2N, and C7H6O2, which were 12 Da less than those produced from the parent drug, indicating that the B ring underwent the process of demethylenation. The observation of product ions at m/z 177 suggests that the A-E-C ring system was retained intact. This provided additional evidence for the occurrence of demethylenation on the B ring. Unlike M1, M2 failed to show the fragment at m/z 177. Instead, it displayed a product ion at m/z 165 (12 Da shorter than m/z 177). This indicates that the formation of M2 resulted from demethylenation of the A ring of corynoline. Thus, we proposed that M1 was a catechol metabolite with a loss of methylene group on the B ring, and M2 was the other positional isomer (Scheme 1).
M3 (retention time = 4.82 minutes) exhibited [M+H]+ at m/z 344, 24 Da less than that of corynoline (Fig. 3B), suggesting that corynoline experienced a loss of two methylene groups during the microsomal reaction. The product ions at m/z 277, 234, and 165 observed in the MS/MS spectrum acquired through MRM-EPI scanning (ion transition m/z 344/277) responded to a loss of CH9O2N, C6H6O2, and C10H13O2N, respectively (Fig. 3G). These ions were all 12 Da shorter relative to that of the parent compound, suggesting that both the A and B rings suffered demethylenation. Based on the fragmentation pattern, we proposed that M3 was the di-catechol metabolite of corynoline (Scheme 1).
Identification of Corynoline NAC Conjugates.
The M4, M5, and M6 metabolites were eluted out at 5.00, 5.64, and 5.78 minutes, respectively (Fig. 4B). The three metabolites shared the same [M+H]+ at m/z 517, suggesting NAC incorporation in M1 and/or M2. The MS/MS spectra acquired by MRM-EPI scanning (ion transition m/z 517/388) showed the indicative characteristic neutral loss of 129 Da associated with the cleavage of the NAC moiety (Fig. 4, E–G). The product ions of M4 and M5 at m/z 177 and 277 (loss of C15H20O5N2S and C7H16O5N2S) were found to be the same as that of M1, indicating that the A ring was retained unchanged. This led us to propose that NAC was conjugated on the F ring. However, no such ions at m/z 177 and 277 were observed in the MS/MS spectrum of M6. In addition, the ion of m/z 197 (with sulfur retained on the catechol), found as a dominant fragment ion of M6, was 32 Da (sulfur) higher than m/z 165, a characteristic fragment of M2 observed in its MS/MS spectrum (Fig. 3F). This indicates that NAC was conjugated on the E ring. Taken together, M4 and M5 were proposed to be NAC conjugates derived from M1, and M6 was a NAC conjugate resulting from M2.
M7 eluted out at the retention time of 4.95 minutes and revealed [M+H]+ of m/z 505 (Fig. 5B), along with the characteristic neutral loss of 129 Da. The molecular ion observed matches the molecular weight of the NAC conjugate derived from M3, the di-catechol metabolite of corynoline. The MS/MS spectrum of M7 obtained by MRM-EPI scanning (ion transition m/z 505/376) demonstrated a product ion at m/z 197, a fragment resulting from the C-E ring system (Fig. 5D), signaling that NAC was incorporated on the E ring as described previously for M6. Additional evidence for the assignment included that M7 failed to display fragments at m/z 177 and 289, the characteristic fragments of M1 and M2. Thus, we proposed that M7 was derived from M3 conjugated with NAC on the E ring.
Chemical synthesis was performed to further characterize the metabolites detected in microsomal incubations. Demethylenation was achieved by refluxing of corynoline in toluene in the presence of AlCl3. The resulting demethylenated products showed the same chromatographic and mass spectrometric identities (Fig. 3, C and H–J) of M1–M3 produced in microsomal incubations. The synthetic demethylenated products were further oxidized with 2,3-dichloro-5,6-dicyano-1,4-benzoquinone, followed by reaction with NAC. The reactions offered three products corresponding to M4, M6, and M7 (NAC conjugates) observed in microsomal incubations (Fig. 4, C, H, and I; Fig. 5, C and E). Unfortunately, we were unable to obtain enough amounts of the individual products for nuclear magnetic resonance characterization due to the complexity and poor yields of the reactions.
Bioactivation of Corynoline in Rats.
To investigate bioactivation of corynoline in vivo, urinary metabolites collected after an i.p. injection of corynoline in rats were analyzed by LC-MS/MS in MRM-EPI mode. Oxidative metabolites M1, M2, and M3 were found in the urine after administration of corynoline, and no such metabolites were observed in the urine before the treatment with corynoline (data not shown). The retention time (Fig. 3D) and major fragment ions (Fig. 3, K–M) of the three metabolites detected in the urine were in agreement with those produced in microsomal incubations.
We failed to detect NAC conjugates (M4–M7) in the urine samples. Instead, we found three metabolites (M8–M10) with molecular ions of m/z 532, which matches the molecular weight of cysteinylglycine (Cys-Gly) conjugates derived from M1 and/or M2. These conjugates resulted from the loss of the glutamyl portion from the corresponding glutathione conjugates. As expected, no such conjugates were detected in the blank urine (data not shown).
The M8–M10 conjugates were detected at retention times of 4.57, 5.23, and 5.58 minutes, respectively (Fig. 6A). The MS/MS spectra of the three metabolites were obtained through MRM-EPI scanning (ion transition m/z 532/356) (Fig. 6, B–D). M8–M10 displayed product ions at m/z 356 resulting from the loss of the Cys-Gly portion. M8 and M9 exhibited product ions of m/z 177 and 277 as M1 did, indicating that the two conjugates were derived from M1. M10 showed product ions at m/z 289 and 165, which were observed in the MS/MS spectrum of M2, suggesting that M10 originated from M2. Thus, we proposed that M8 and M9 were Cys-Gly conjugates derived from M1, and M10 was a Cys-Gly conjugate resulting from M2.

Identification of metabolites M8, M9, and M10. (A) Extracted ion (m/z 532/356 for M8, M9, and M10) chromatograms obtained from LC-MS/MS analysis of urine samples of rat treated with corynoline. MS/MS spectra of M8 (B), M9 (C), and M10 (D) obtained from LC-Q-Trap/MS analysis of urine samples of rats given corynoline.
Identification of Human Recombinant CYPs Responsible for Metabolic Activation of Corynoline.
To identify which individual human recombinant CYPs are mainly involved in metabolic activation of corynoline, eight CYP isoforms, including the CYPs 1A2, 2B6, 2C9, 2C19, 2D6, 2E1, 3A4, and 3A5, were individually incubated with corynoline in the presence of NAC as the trapping agent, followed by monitoring the formation of M4 and M5. We found that multiple CYP isoforms mediated M4 and M5 formation. Among them, the CYPs 2C9, 3A4, and 2C19 were the primary enzymes contributing to the metabolic activation of corynoline (Fig. 7).

Individual human recombinant CYP enzymes involved in bioactivation of corynoline. Corynoline was incubated with individual human recombinant CYP isoforms in the presence of NADPH and NAC. The catalytic capabilities of the enzymes were evaluated by monitoring the formation of M4 and M5 after normalization based on the relative content of the corresponding P450 enzyme in human liver microsomes. Data represent the mean ± S.D. (n = 3).
Discussion
With the rapidly growing global interest in natural products as medical remedies, dietary supplements, and natural foods, more and more cases of natural product poisonings have been reported. Some natural products are toxic and may cause severe adverse effects, including fetal problems in humans. Thus, safe consumption of such natural products is becoming a public concern. Corynoline is one of the major components in C. Bungeanae Herba, which is widely used as a traditional herbal medicine. Although corynoline was reported to cause cytotoxicity (Choi et al., 2007), metabolism of corynoline has not been carefully investigated. Corynoline contains two 1,3-benzodioxole groups, and many 1,3-benzodioxole-containing compounds were found to induce toxicities (Erve et al., 2013). Metabolic activation is suggested to initiate the process of the toxicities induced by these toxic compounds. Specifically, the bioactivation possibly starts with O-dealkylation, by which the 1,3-benzodioxole group is biotransformed to the corresponding catechol, and sequential oxidation of the catechol produces an ortho-quinone. The resulting quinone may be responsible for the toxicities. Three O-dealkylation metabolites were detected in microsomal incubations with corynoline. The three oxidative metabolites were characterized as two isomeric catechols (M1 and M2) and one di-catechol (M3), respectively. Additionally, a total of four NAC conjugates (M4–M7) were identified in NAC- and corynoline-fortified microsomal reactions, and these conjugates were derived from the respective M1, M2, and M3 metabolites. The formation of the demethylenated metabolites and the NAC conjugates was found to require the presence of NADPH in the microsomal incubations. The data clearly indicate the production of corynoline-drived ortho-quinone intermediates and the participation of cytochrome P450 enzymes in metabolic activation of corynoline.
Chemical synthesis was executed to verify the metabolite identification work. 1,3-Benzodioxole is basically an acetal derivative that can be hydrolyzed to produce a catechol and an aldehyde. The synthetic work was started with refluxing of corynoline in toluene catalyzed by Lewis acid AlCl3. Apparently, the hydrolytic reaction was not selective and gave three products that displayed the identical retention time and mass spectra as that of M1–M3 observed in microsomal incubations containing corynoline and NADPH. The resulting hydrolytic products were further oxidized by 2,3-dichloro-5,6-dicyano-1,4-benzoquinone, followed by reaction with NAC. LC-MS/MS analysis demonstrated that the resultant reactions offered three NAC conjugates that elicited the same chromatographic and mass spectrometric behaviors as that of M4, M6, and M7 observed in the corresponding microsomal incubations. The synthetic work allowed us to confirm the structures of M1–M7 (except for M5). Unexpectedly, the product corresponding to M5 was not found in the chemical reaction mixture. The reason for the failed formation of the product remains unknown.
Corynoline was reported to display time-dependent inhibition on CYP3A4 (Fang et al., 2011). Time-dependent inhibition is often a criterion for irreversible enzyme inhibition. Covalent modification of the P450 apoprotein, alkylation of the heme moiety, or destruction of the heme group via the metabolic intermediate complex (Erve et al., 2013) is considered to trigger the enzyme inactivation. Our observation of ortho-quinone intermediate provided the evidence for a possible mechanism by which the reactive species alkylated the apoprotein and/or heme of the host enzyme. In addition, we cannot exclude that the 1,3-benzodioxole moieties were biotransformed to the corresponding reactive carbenes that chelate the iron of the P450 heme group to form metabolite-intermediate complexes, resulting in the interference of the functioning of the heme.
Urinary NAC conjugates (mercapturic acids) are often used as biomarkers of exposure to electrophilic agents. These conjugates arise from the corresponding glutathione conjugates. The process of conversion from glutathione conjugates to NAC conjugates involves sequential removal of the Glu residue (to produce Cys-Gly conjugates) and Gly portion, catalyzed by γ-glutamyltranspeptidase and Cys-Gly dipeptidase, respectively. The resulting Cys conjugates are N-acetylated to form NAC conjugates in the kidney before excretion. To investigate the bioactivation process of corynoline in vivo, we analyzed the urine samples of rats treated with corynoline. We failed to detect any NAC and Cys conjugates derived from corynoline in the urine. Instead, Cys-Gly conjugates (M8–M10) resulting from M1 and M2 were found in the urine. The absence of the Cys and NAC conjugates may be attributed to the following: 1) the corresponding Cys-Gly conjugates are not the substrates of Cys-Gly dipeptidase, and 2) these conjugates are the inhibitors of the dipeptidase. Despite this, the observation of the urinary Cys-Gly conjugates provided in vivo evidence for metabolic activation of corynoline.
In conclusion, the present study demonstrated that corynoline was metabolized to three catechols and was further oxidized to the corresponding ortho-quinones in vitro and in vivo. CYPs 2C9, 3A4, and 2C19 mainly contributed to the metabolic activation of corynoline. The results provided important information about the potential risk of adverse effects in consumption of corynoline and corynoline-containing remedies.
Authorship Contributions
Participated in research design: Zheng, Peng.
Conducted experiments: Mao, Peng.
Performed data analysis: Mao.
Wrote or contributed to the writing of the manuscript: Mao, Zheng.
Footnotes
- Received June 10, 2015.
- Accepted August 4, 2015.
This work was supported in part by the National Natural Science Foundation of China [Grants 81430086 and 81373471].
Abbreviations
- CYP
- cytochrome P450
- Cys-Gly
- cysteinylglycine
- EPI
- enhanced product ion
- LC-MS/MS
- liquid chromatography–tandem mass spectrometry
- MRM
- multiple-reaction monitoring
- NAC
- N-acetylcysteine
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics
References
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}