


Visual Overview
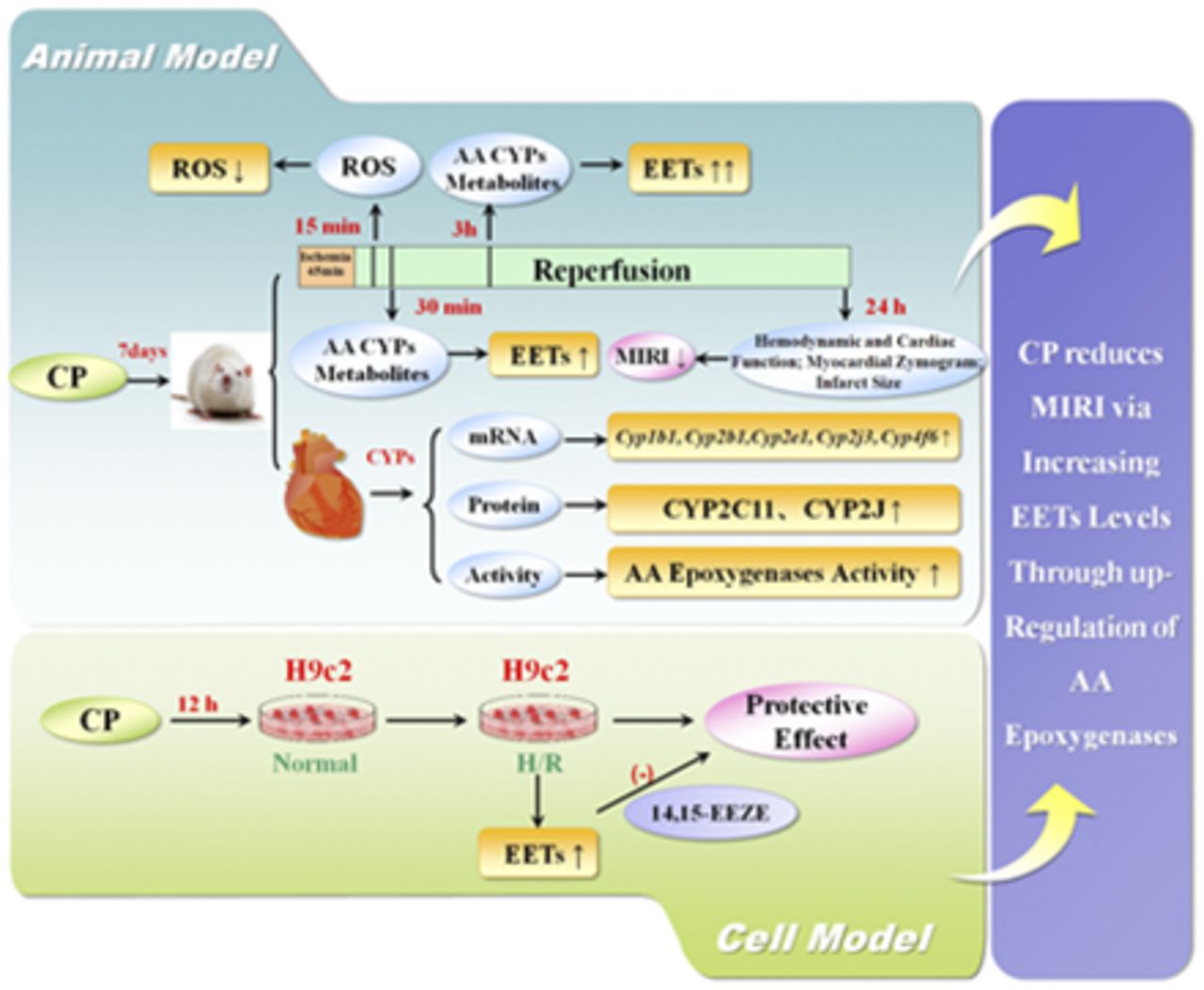
Abstract
Accumulating data suggest that epoxyeicosatrienoic acids (EETs) and 20-hydroxyeicosatetraenoic acid, both cytochrome P450 (P450) enzyme metabolites of arachidonic acid (AA), play important roles in cardiovascular diseases. For many years, the cardiotonic pill (CP), an herbal preparation derived from Salviae Miltiorrhizae Radix et Rhizoma, Notoginseng Radix et Rhizoma, and Borneolum Syntheticum, has been widely used in China for the treatment of coronary artery disease. However, its pharmacological mechanism has not been well elucidated. The purpose of this study was to investigate the chronic effects of the CP on myocardial ischemia-reperfusion injury (MIRI) and AA P450 enzyme metabolism in rats (in vivo) and H9c2 cells (in vitro). The results showed that CP dose dependently (10, 20, and 40 mg/kg/d; 7 days) mitigated MIRI in rats. The plasma concentrations of EETs in CP-treated ischemia-reperfusion (I/R) rats (40 mg/kg/d; 7 days) were significantly higher (P < 0.05) than those in controls. Cardiac Cyp1b1, Cyp2b1, Cyp2e1, Cyp2j3, and Cyp4f6 were significantly induced (P < 0.05); CYP2J and CYP2C11 proteins were upregulated (P < 0.05); and AA-epoxygenases activity was significantly increased (P < 0.05) after CP (40 mg/kg/d; 7 days) administration in rats. In H9c2 cells, the CP also increased (P < 0.05) the EET concentrations and showed protection in hypoxia-reoxygenation (H/R) cells. However, an antagonist of EETs, 14,15-epoxyeicosa-5(Z)-enoic acid, displayed a dose-dependent depression of the CP’s protective effects in H/R cells. In conclusion, upregulation of cardiac epoxygenases after multiple doses of the CP—leading to elevated concentrations of cardioprotective EETs after myocardial I/R—may be the underlying mechanism, at least in part, for the CP’s cardioprotective effect in rats.
Footnotes
- Received February 4, 2016.
- Accepted May 4, 2016.
↵1 M.X. and H.H. contributed equally to this work.
This work was sponsored by the National Natural Science Foundation of China [Grants 81130068 and 81202984]; the National Science and Technology Major Project “Creation of Major New Drugs” [Grant 2012ZX09303009-002]; the Priority Academic Program Development of Jiangsu Higher Education Institutions [PAPD]; the 12th Six Talent Peaks Project in Jiangsu Province [Grant WSN-054]; and the Leading Talents of Scientific Research in TCM of Jiangsu Province [Grant LJ200906].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}