


Visual Overview

Abstract
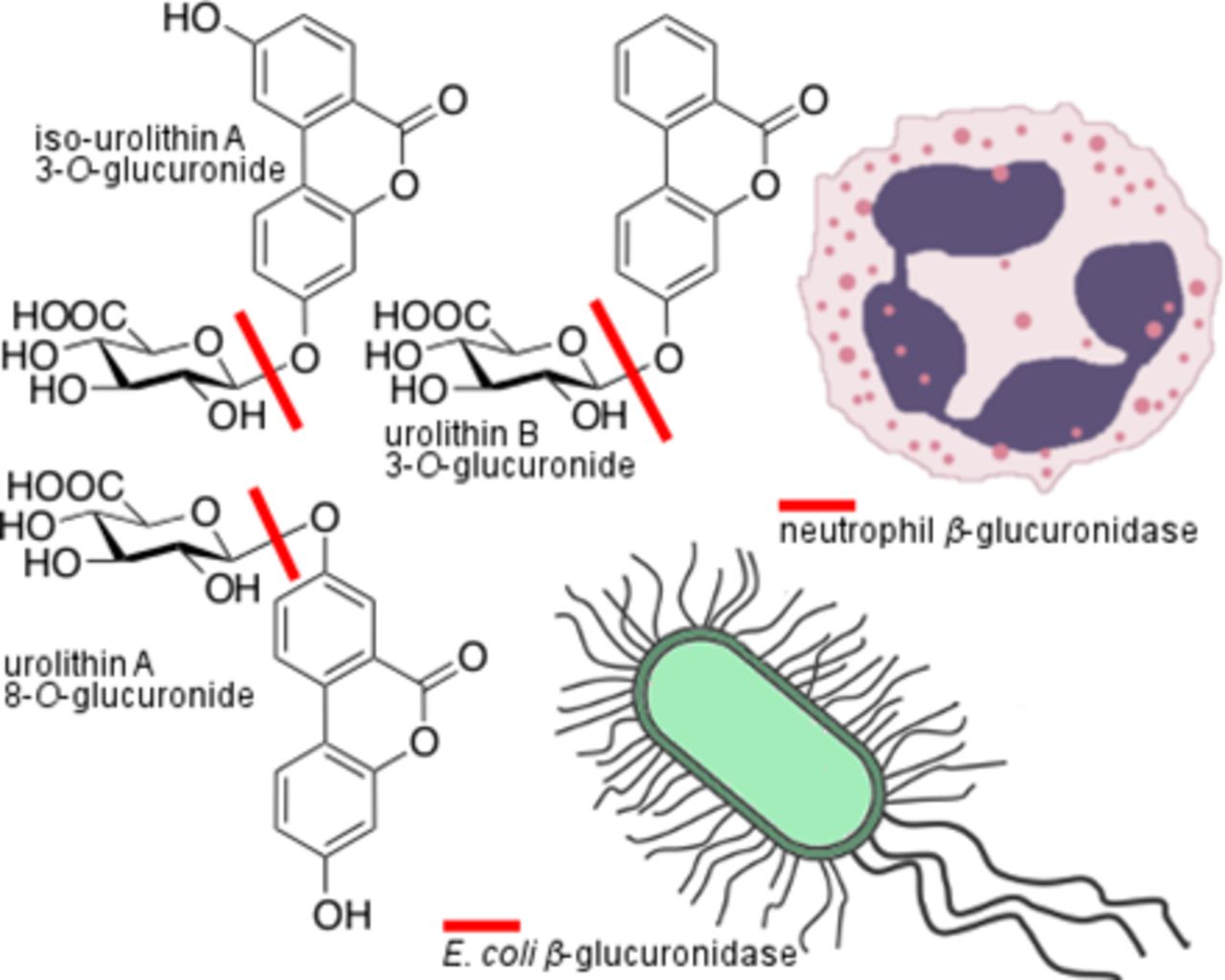
In recent years, many xenobiotics derived from natural products have been shown to undergo extensive metabolism by gut microbiota. Ellagitannins, which are high molecular polyphenols, are metabolized to dibenzo[b,d]pyran-6-one derivatives—urolithins. These compounds, in contrast with their parental compounds, have good bioavailability and are found in plasma and urine at micromolar concentrations. In vivo studies conducted for ellagitannin-containing natural products indicate their beneficial health effects toward inflammation and cancer, which are associated with the formation of urolithins. However, the great majority of in vitro experiments that have revealed the molecular mechanisms responsible for the observed effects were conducted for urolithin aglycones. These studies are thus incongruent with the results of pharmacokinetic studies that clearly indicate that glucuronide conjugates are the dominant metabolites present in plasma, tissue, and urine. The aim of this study was to isolate and structurally characterize urolithin conjugates from the urine of a volunteer who ingested ellagitannin-rich natural products, and to evaluate the potential role of β-glucuronidase–triggered cleavage in urolithin disposition. Glucuronides of urolithin A, iso-urolithin A, and urolithin B were isolated and shown to be cleaved by the β-glucuronidases released by neutrophils from azurophilic granules upon N-formylmethionine-leucyl-phenylalanine stimulation as well as by Escherichia coli standard strains and clinical isolates from patients with urinary tract infections. These results justify the hypothesis that the selective activation of urolithin glucuronides by β-glucuronidase, which are present at high concentrations at inflammation and infection sites and in the microenvironments of solid tumors, could locally increase the concentration of bioactive urolithin aglycones.
Footnotes
- Received January 24, 2017.
- Accepted March 1, 2017.
The project was financially supported by a Polish Ministry of Science and Higher Education research grant, Iuventus Plus [Grant IP2015 062274]. J.P.P. was financially supported by a Foundation for Polish Science START scholarship [START 84.2016]. The project was performed using Centre for Preclinical Research and Technology infrastructure financed by the European Regional Development Fund within the “Innovative Economy” Operational Programme for 2007–2013.
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

- Copyright © 2017 by The American Society for Pharmacology and Experimental Therapeutics
DMD articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}