Article Text

Abstract
Background: A recent study from our laboratory showed that induction of the multidrug resistance related drug efflux pump ABCG2 contributed to acquired resistance of human T cells to the disease modifying antirheumatic drug (DMARD) sulfasalazine (SSZ).
Objectives: To investigate the duration of SSZ resistance and ABCG2 expression after withdrawal of SSZ and rechallenging with SSZ, and to assess the impact of SSZ resistance on responsiveness to other DMARDs.
Methods: Human CEM cells (T cell origin) with acquired resistance to SSZ (CEM/SSZ) were characterised for (a) SSZ sensitivity and ABCG2 expression during withdrawal and rechallenge of SSZ, and (b) antiproliferative efficacy of other DMARDs.
Results: ABCG2 protein expression was stable for at least 4 weeks when CEM/SSZ cells were grown in the absence of SSZ, but gradually declined, along with SSZ resistance levels, to non-detectable levels after withdrawal of SSZ for 6 months. Rechallenging with SSZ led to a rapid (<2.5 weeks) resumption of SSZ resistance and ABCG2 expression as in the original CEM/SSZ cells. CEM/SSZ cells displayed diminished sensitivity to the DMARDs leflunomide (5.1-fold) and methotrexate (1.8-fold), were moderately more sensitive (1.6–2.0 fold) to cyclosporin A and chloroquine, and markedly more sensitive (13-fold) to the glucocorticoid dexamethasone as compared with parental CEM cells.
Conclusion: The drug efflux pump ABCG2 has a major role in conferring resistance to SSZ. The collateral sensitivity of SSZ resistant cells for some other (non-related) DMARDs may provide a further rationale for sequential mono- or combination therapies with distinct DMARDs upon decreased efficacy of SSZ.
- disease modifying antirheumatic drugs
- multidrug resistance
- sulfasalazine
- ABC, ATP binding cassette
- BCRP, breast cancer resistance protein
- DMARDs, disease modifying antirheumatic drugs
- FCS, fetal calf serum
- IC50, drug concentration required to inhibit cell growth by 50%
- MDR, multidrug resistance
- MRP1, multidrug resistance associated protein 1
- MTX, methotrexate
- NFκB, nuclear factor κB
- Pgp, P-glycoprotein
- SSZ, sulfasalazine
- TNFα, tumour necrosis factor α
Statistics from Altmetric.com
- ABC, ATP binding cassette
- BCRP, breast cancer resistance protein
- DMARDs, disease modifying antirheumatic drugs
- FCS, fetal calf serum
- IC50, drug concentration required to inhibit cell growth by 50%
- MDR, multidrug resistance
- MRP1, multidrug resistance associated protein 1
- MTX, methotrexate
- NFκB, nuclear factor κB
- Pgp, P-glycoprotein
- SSZ, sulfasalazine
- TNFα, tumour necrosis factor α
Acquired drug resistance is recognised as a common cause of treatment failure for patients with cancer after sustained treatment with cytostatic drugs.1,2 An important mechanism of acquired cellular drug resistance to anticancer drugs is the overexpression of drug efflux pumps belonging to the superfamily of adenosine triphosphate dependent binding cassette (ABC) transporters.2–6 ABC transporters with an established role in drug efflux and drug resistance include (a) P-glycoprotein (Pgp, ABCB1), which confers resistance preferentially to lipophilic drugs7; (b) members of the multidrug resistance associated protein subfamily (MRP1-6, ABCC1-6), which confer resistance to various anionic charged/glutathione conjugated drugs such as methotrexate (MTX),8 thiopurines,9,10 and steroids5,11; and (c) ABCG2, also known as breast cancer resistance protein (BCRP), a recently identified ABC transporter that confers resistance to amphiphilic drugs, including mitoxantrone and MTX.12,13
In clinical rheumatology, loss of drug efficacy is also noted for patients with rheumatoid arthritis (RA) receiving long term treatment with disease modifying antirheumatic drugs (DMARDs).14–17 Whether this loss of DMARD efficacy is related to the onset of acquired resistance to DMARDs, with mechanisms similar to anticancer drug resistance, is not known.18 Meta-analyses of DMARD treatment termination rates showed that the DMARD sulfasalazine (SSZ)19,20 had a relatively shorter lasting efficacy than other DMARDs, in particular the “gold standard” MTX.14,16–18 In general, lack of efficacy may be related to diminished anti-inflammatory and/or antiproliferative effects of DMARDs against proinflammatory cytokine releasing cells at inflammatory sites.21
To gain insight into the possible mechanism(s) of resistance to the antiproliferative/anti-inflammatory effects of the DMARD SSZ, we previously provoked acquired resistance to SSZ in an in vitro model system of human T lymphocytes by stepwise exposure of CEM (T) cells to gradually increasing concentrations of SSZ.22 This study showed that SSZ resistance in CEM (T) cells was conferred by overexpression of a drug efflux pump—notably, the multidrug resistance transporter ABCG2/BCRP. Owing to enhanced drug efflux in SSZ resistant cells (CEM/SSZ), higher concentrations of SSZ were required to inhibit the secretion of the proinflammatory cytokine tumour necrosis factor α (TNFα).22
In the present study we investigated the stability of the resistant phenotype of CEM/SSZ cells after withdrawal of SSZ in order to establish whether SSZ resistance is slowly/rapidly lost over time and reinducible after SSZ rechallenge. Moreover, we analysed to what extent SSZ resistance affected antiproliferative effects of other DMARDs and anti-inflammatory drugs. The results demonstrated that SSZ resistance, along with ABCG2 expression, was gradually lost over a period of 6 months after withdrawal of SSZ, but rapidly re-emerged, within 2 weeks, upon rechallenge with SSZ. Drug sensitivity was as follows: CEM/SSZ cells displayed lower sensitivity (fivefold) to the DMARD leflunomide, a slightly diminished sensitivity to MTX (twofold), moderately increased sensitivities for cyclosporin A and chloroquine (up to twofold), but a markedly enhanced sensitivity for dexamethasone (13-fold).
MATERIALS AND METHODS
Materials
Sulfasalazine, leflunomide, chloroquine, phenylmethylsulfonyl fluoride (PMSF), and dexamethasone were purchased from Sigma Chemical Co (St Louis, MO, USA). Protease inhibitor cocktail and Triton X-100 were from Boehringer Mannheim (Ingelheim, Germany). MG132 was from Calbiochem (Germany). MTX was a generous gift from Pharmachemie Haarlem, the Netherlands. Cyclosporin A was a gift from Novartis (Arnhem, the Netherlands). The ABCG2 inhibitor Ko14323 was kindly provided by Dr A Schinkel (Netherlands Cancer Institute, Amsterdam, the Netherlands). RPMI-1640 tissue culture medium and fetal calf serum (FCS) were obtained from Gibco Chemical Co, Grand Island NY, USA.
Cell culture and selection of SSZ resistant CEM (T) cells
SSZ resistant human CEM (T) cells were isolated as described previously.22 In short, human CEM (T) cells were cultured at an initial density of 3×105 cells/ml in RPMI-1640 medium supplemented with 10% FCS, 2 mM l-glutamine, and 100 µg/ml penicillin and streptomycin in a 5% CO2 incubator at 37°C.24,25 Cell cultures were refreshed twice weekly and SSZ was added to the cell culture medium at an initial concentration of 0.4 mmol/l. This concentration of SSZ could be stepwise increased to 1.5 mM SSZ (after 4 months) and 2.5 mM SSZ after a period of 6 months.
Extensive binding of SSZ to plasma proteins (serum albumin) is known to limit its therapeutic effect.26,27 Based on the presence of at least three classes of SSZ binding sites on serum albumin and their association rate constants28 it can be calculated that a protein concentration of about 100 µmol/l in 10% FCS can bind 320 µM SSZ. For the two selected SSZ resistant CEM cells grown in the presence of 1.5 and 2.5 mM SSZ, this would imply that 21% and 13% of SSZ, respectively, is protein bound.
It should be recognised that CEM cells are of T cell leukaemic origin, proliferating with a doubling time of 26–30 hours. Thus CEM cells may not fully representative for normal T cells or T cells applicable to RA. However, CEM cells may serve as a valuable in vitro model system for RA research as they respond to T cell stimuli, producing proinflammatory cytokines like TNFα.22 Furthermore, CEM cells have been exploited to disclose resistance mechanisms to MTX that are clinically encountered.24,29–31
Antiproliferative effects of DMARDs and other drugs were analysed by plating 1.25×105 cells/ml of parental CEM (T) cells, CEM/SSZ1.5, and CEM/SSZ2.5 cells (in the absence of SSZ) in individual wells of a 24 well plate containing up to 50 µl of drug solution. Inhibition of cell growth was determined after 72 hours’ incubation by counting cells with a haemocytometer and cell viability by trypan blue exclusion. The drug concentration required to inhibit cell growth by 50% compared with control growth is defined as the IC50 concentration.
Western blotting
For analysis of expression of ABCG2/BCRPand ABCC1/MRP1, cells were harvested in the mid-log phase of growth and washed three times with ice cold Hepes buffered saline, pH 7.4. Total cell lysates of 107 cells were suspended in 500 µl of lysis buffer containing 50 mM Tris-HCl (pH 7.6), 5 mM dithiothreitol, 20 µl protease inhibitor cocktail (one tablet/ml H2O), 20% glycerol, and 0.5% NP-40. The suspension was sonicated (MSE sonicator, amplitude 6, for 3×5 seconds with 30 seconds time intervals at 4°C) and centrifuged in an Eppendorf microcentrifuge (10 minutes, 14 000 rpm, 4°C). Protein content of the supernatant was determined by Biorad protein assay.32 Fifty microgram of total cell lysates was fractionated on a 7.5% polyacrylamide gel and transferred onto a nitrocellulose membrane. The nitrocellulose membranes were preincubated overnight at 4°C in blocking buffer (5% Biorad Blocker in TBS-T (10 mM Tris-HCl, pH 8.0, 0.15 M NaCl, 0.1% Tween-20) to prevent aspecific antibody binding. After blocking, the membranes were incubated for 1 hour at room temperature with the primary antibodies for the multidrug resistant (MDR) pumps ABCG2/BCRP (BXP21, 1:400) and MRP1 (MRPr1, 1:500) as described by Scheffer et al.32,33 As control for protein loading, β-actin (Chemicon International, Ca, USA: 1:3000) was used. After three washing steps with TBS-T, membranes were incubated for 1 hour with horseradish peroxidase labelled antirat/mouse (Dako; 1:2000 for ABCG2/BCRP and ABCC1/MRP1, and 1:3000 for β-actin) as secondary antibody. Detection of the antibody binding was measured by enhanced chemiluminescence according to the manufacturer’s instructions (Amersham International, Buckinghamshire, UK). Protein levels were determined by densitometric scanning (GelDoc and Molecular Analyst, Biorad Laboratories) of the x ray films (Hyperfim ECL, Amersham International, Buckinghamshire, UK).
Statistical analysis
Statistical significance of differences was analysed by analysis of variance using SPSS 9.0 computer software. A p value <0.05 was considered to be significantly different.
RESULTS
Onset and stability of SSZ resistance in human CEM (T) cells
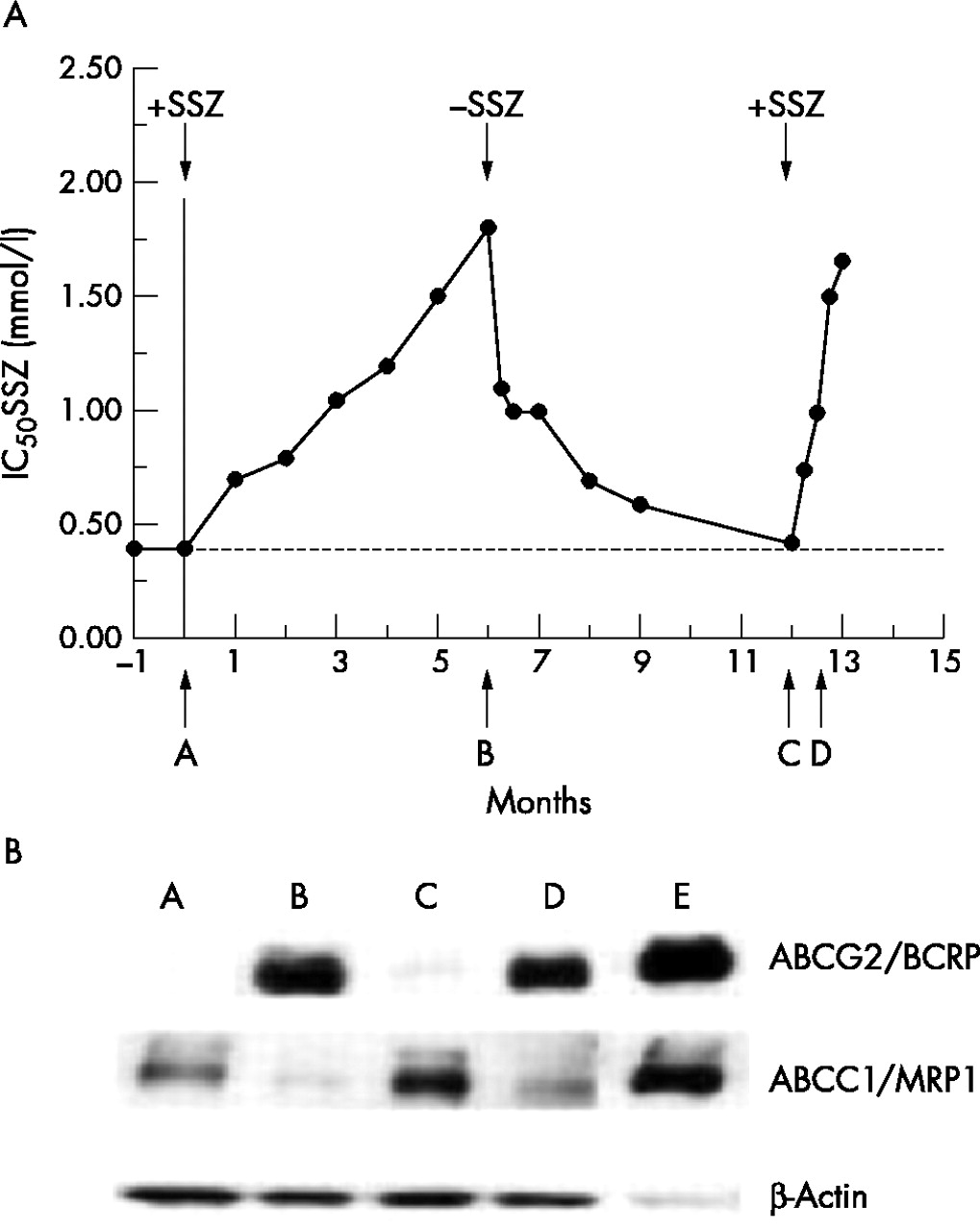
Acquired resistance to SSZ in human CEM (T) cells could be provoked by culturing cells in stepwise increasing concentrations of SSZ.22Figure 1A (point A) shows the antiproliferative effect of SSZ (IC50: concentration for 50% growth inhibition) for CEM (T) cells at the initial dosing of 0.4 mM SSZ, and for CEM (T) cells that were grown in the presence of stepwise increasing concentrations of SSZ. After a period of 4 months, CEM (T) could be maintained at an IC50 concentration of 1.5 mM SSZ (CEM/SSZ1.5) which was about fourfold higher than parental CEM (T) cells. When CEM/SSZ1.5 cells were grown in the presence of 1.5 mM SSZ for another 2 months, the IC50 value slightly increased to 1.7 mM (fig 1A, point B). When, in parallel, CEM/SSZ1.5 cells obtained after 4 months were exposed to further increasing concentrations of SSZ for 2 months, eventually CEM/SSZ2.5 cells were obtained that could grow in the presence of 2.5 mM SSZ (not shown).

(A) Time course of sensitivity to sulfasalazine (SSZ) of CEM (T) during development of SSZ resistance, after withdrawal of SSZ from CEM/SSZ1.5 cells and rechallenge of revertant CEM/SSZ1.5 cells with SSZ. Parental CEM (T) cells (point A) were exposed to a starting concentration of 0.4 mM SSZ that was gradually increased to 1.5 mM over a period of 6 months to yield SSZ resistant CEM/SSZ1.5 cells (point B). From this point on CEM/SSZ1.5 were grown in the absence of SSZ to finally yield revertant CEM/SSZ1.5 cells that displayed again parental CEM (T) cell sensitivity to SSZ (point C). When revertant cells were re-exposed to 1.5 mM SSZ for 2.5 weeks (point D), CEM/SSZ1.5 resumed their original resistance level as observed at point B. IC50 values were determined after 72 hours’ exposure of cells to SSZ. (B) ABCG2/BCRP and ABCC1/MRP1 protein expression, and mitoxantrone sensitivity of CEM (T) cells and (revertant) CEM/SSZ1.5 cells isolated at the various time points A–D. Cell lysates of 2008/MRP132 and MCF7/MR43 served as positive controls for ABCC1/MRP1 and ABCG2/BCRP, respectively (not shown). For CEM (T) and CEM/SSZ1.5 cells, 50 µg of total cell lysate was applied on the sodium dodecyl sulphate polyacrylamide gel, and for the controls 10 µg protein. ABCC1/MRP1 was detected by the monoclonal antibody MRPr1, and ABCG2/BCRP was detected by the monoclonal antibody BXP21.32,33 β-Actin was used as a control for equal protein loading.
To establish whether the SSZ resistance in CEM/SSZ1.5 cells is a transient or stable phenotype, CEM/SSZ1.5 cells (at point B) were further grown in the absence of SSZ, and IC50 values for the CEM/SSZ1.5 cells were monitored over time. After 1 week in the absence of SSZ, IC50 values for SSZ dropped by 40–50% to stay at this level for up to 4 weeks. From this point on, IC50 values for SSZ gradually declined to reach ultimately the original levels of SSZ sensitivity of parental CEM (T) cells (point C). To assess whether SSZ resistance would be reinduced upon exposure to SSZ, revertant CEM/SSZ1.5 cells were re-exposed to 1.5 mM SSZ. Within only 2.5 weeks’ exposure to SSZ (D), revertant CEM/SSZ1.5 cells resumed their original level of SSZ resistance as seen at point B. These results indicate that although SSZ resistance in CEM/SSZ is transiently lost over a period of 6 months, rechallenge of revertant cells with SSZ rapidly reinduces resistance to SSZ.
Expression of MDR transporters ABCG2/BCRP and ABCC1/MRP1 in CEM (T) and CEM/SSZ cells
Initial studies from our laboratory22 reported differential expression of MDR proteins upon development of SSZ resistance in CEM/SSZ cells: induction of ABCG2/BCRP and down regulation of ABCC1/MRP1 expression. Figure 1B illustrates the expression of ABCG2/BCRP and ABCC1/MRP1 at the various times A–D in fig 1A. Parental CEM (T) cells (point A, fig 1A) exhibited undetectable levels of expression of ABCG2/BCRP protein, but had an appreciable level of ABCC1/MRP1. In contrast, SSZ resistant CEM cells isolated at 1.5 mM SSZ (CEM/SSZ1.5 cells) (fig 1A, point B) showed a marked induction of ABCG2/BCRP expression and down regulation of ABCC1/MRP1 expression. After withdrawal of SSZ from the cell culture of CEM/SSZ1.5 for a period of 6 months (fig 1A, point C) ABCG2/BCRP and ABCC1/MRP1 expression reverted to the original levels of parental CEM (T) cells. Notably, when revertant CEM/SSZ1.5 were re-exposed to 1.5 mM SSZ for 2.5 weeks (fig 1A, point D), ABCG2/BCRP expression was rapidly up regulated, while ABCC1/MRP1 was again down regulated. As a control, fluctuations in the expression of ABCG2/BCRP paralleled the growth inhibitory effects (IC50 values) for the cytostatic drug mitoxantrone, which is a known substrate for efflux by ABCG2/BCRP6,34 (data not shown).
Because fig 1A showed that changes in IC50 values for CEM/SSZ1.5 cells were particularly noted within the first 10 weeks after withdrawal of SSZ (fig 1A, point B→C), we fine tuned the analysis of IC50 values for SSZ and changes in expression of ABCG2/BCRP and ABCC1/MRP1 during this period of 10 weeks. IC50 values for SSZ dropped by about 40–50% after growth of CEM/SSZ1.5 cells in the absence of SSZ for 1 week (fig 2A). From this point on, IC50 values for SSZ only slowly declined. At 70 days after withdrawal of SSZ from CEM/SSZ1.5 cells, resistance to SSZ could still be noted compared with parental CEM (T) cells. Figure 2B shows the expression of ABCG2/BCRP and ABCC1/MRP1 in parental CEM (T) cells (lane A) compared with CEM/SSZ1.5 cells before (lane B) and after withdrawal of SSZ for 3 days, 7 days, 4 weeks, and 10 weeks (lanes C–F, respectively). ABCG2/BCRP protein expression in CEM/SSZ1.5 cells appeared to be stable for at least 4 weeks after withdrawal of SSZ, after which a diminished, but still detectable, level of expression could be seen after 10 weeks. ABCC1/MRP1 expression, which was down regulated in CEM/SSZ1.5 cells in the presence of SSZ, was rapidly reinduced within 3–7 days after withdrawal of SSZ, even overshooting the level of ABCC1/MRP1 expression in parental CEM (T) cells after 70 days of SSZ withdrawal from CEM/SSZ1.5 cells. These results indicate that ABCG2/BCRP and ABCC1/MRP1 follow an inverse pattern of expression after SSZ exposure/withdrawal.

{kind=link}
{kind=link}
(A) Antiproliferative effect of SSZ against CEM/SSZ1.5 cells isolated at various times (0–10 weeks) after withdrawal of SSZ from CEM/SSZ1.5 cells from point B in fig 1A: 0 days (closed circles), 3 days (open triangles), 1 week (closed triangles), 4 weeks (open diamonds), 10 weeks (closed squares). Antiproliferative effects/growth inhibition for SSZ were evaluated after 72 hours’ exposure to SSZ and compared with those for parental CEM (T) cells (open circles). (B) Expression of ABCG2/BCRP and ABCC1/MRP1 protein of CEM/SSZ1.5 cells during withdrawal of SSZ for up to 10 weeks as in fig 2A. Lane A: parental CEM (T) cells; lane B: CEM/SSZ1.5 cells before withdrawal of SSZ; lanes C–F: CEM/SSZ1.5 cells 3 days, 7 days, 4 weeks, and 10 weeks, respectively, after withdrawal of SSZ. Note: sample E for ABCC1/MRP1 was not tested. (C) Reversal of SSZ resistance in CEM/SSZ1.5 cells by the ABCG2/BCRP blocker Ko143 before and after 4 weeks’ withdrawal of SSZ. Symbols: (open triangle) parental CEM (T) cells, (open circles) CEM/SSZ1.5 cells, (closed circles) CEM/SSZ cells + 0.5 µM Ko143, (open squares) CEM/SSZ1.5 cells after 4 weeks’ withdrawal of SSZ, (closed squares) CEM/SSZ1.5 cells after 4 weeks withdrawal of SSZ + 0.5 µM SSZ. Antiproliferative effects of SSZ were evaluated after 72 hours’ exposure to SSZ in the absence or presence of Ko143.
Given the apparent discrepancy of the rapid partial loss of SSZ resistance within 1 week of withdrawal of SSZ from CEM/SSZ1.5 cells (fig 2A) and the stable expression of ABCG2/BCRP during this period (fig 2B, lanes B–D), we further investigated the functional contribution of ABCG2/BCRP in SSZ resistance by blocking ABCG2/BCRP with a specific inhibitor, Ko143.23 Blocking of ABCG2/BCRP reversed SSZ resistance in CEM/SSZ1.5 cells by 50–60%, but not completely, to parental CEM (T) cell sensitivity (fig 2C). After growth of CEM/SSZ1.5 cells in the absence of SSZ for 4 weeks, when IC50 values for SSZ had stabilised at about 50% of the original CEM/SSZ1.5 cells, Ko143 fully reversed the residual SSZ resistance of CEM/SSZ1.5 cells to the SSZ sensitivity of CEM (T) cells (fig 2C). These results indicate that SSZ resistance in CEM/SSZ1.5 is mediated by at least two components, each contributing about 50%. One component, is rapidly lost within one week after withdrawal of SSZ, the other component, which stabilised after 1–4 weeks’ growth of CEM/SSZ1.5 cells in the absence of SSZ, can be fully accounted for by ABCG2/BCRP. The latter component is then gradually lost over period of 5 months’ culture in the absence of SSZ.
Sensitivity profile of CEM/SSZ cells for DMARDs and other anti-inflammatory drugs
In clinical rheumatology, SSZ is applied sequentially in mono- and/or combination therapies with other DMARDs.19,35,20 For this reason, we investigated whether the onset of SSZ resistance affects the sensitivity for other clinically active DMARDS, as well as other drugs with potentially anti-inflammatory properties. Drug sensitivities were evaluated by their antiproliferative effects for CEM/SSZ1.5 and CEM/SSZ2.5 cells as compared with parental CEM (T) cells (table 1).
Antiproliferative effects of DMARDs and other drugs against CEM (T) and CEM/SSZ cells
Cross resistance to mitoxantrone, a topoisomerase II inhibitor that is a prototypical substrate for ABCG2/BCRP34,36 for both CEM/SSZ1.5 (4.3-fold, p = 0.002 v CEM ) and CEM/SSZ2.5 cells (6.4-fold, p<0.001 v CEM), was studied. As expected, mitoxantrone cross resistance was fully reversed by the ABCG2/BCRP blocker Ko143. Furthermore, cross resistance of CEM/SSZ cells was noted for the DMARDs leflunomide (up to 5.1-fold for CEM/SSZ2.5, p = 0.005 v CEM) and MTX (up to 1.8-fold for CEM/SSZ2.5, p = 0.027 v CEM). Because SSZ mediates part of its anti-inflammatory effects by targeting the NFκB signalling pathway,37,38 we also determined the drug sensitivity for MG132, an inhibitor of NFκB activation through inhibition of 26S-proteasome.39 A low level of cross resistance for CEM/SSZ cells was noted for MG132 (1.6-fold, p = 0.004 for CEM/SSZ2.5 and p<0.001 for CEM/SSZ1.5 v CEM). Interestingly, enhanced sensitivity of CEM/SSZ cells was noted for the DMARDs chloroquine (up to 1.9-fold CEM/SSZ2.5, p = 0.043 v CEM) and cyclosporin A (up to 1.6-fold, p⩽0.001 for CEM/SSZ1.5 and p = 0.028 for CEM/SSZ2.5 v CEM). CEM/SSZ cells displayed even more hypersensitivity (up to 13-fold, p = 0.002 for CEM/SSZ1.5 and p = 0.003 for CEM/SSZ2.5 v CEM) to the glucocorticoid dexamethasone. These results indicate that induction of SSZ resistance can coincide with a differentially altered DMARD sensitivity pattern, including, on the one hand, cross resistance to leflunomide and MTX, but on the other, enhanced sensitivity to cyclosporin A and chloroquine, and, most notably, to dexamethasone.
DISCUSSION
Characterisation of human CEM (T) cells with acquired resistance to the DMARD SSZ showed differential expression of two MDR efflux pumps: up regulation of ABCG2/BCRP and down regulation of ABCC1/MRP1. This phenotype could be reverted (slowly) and reinduced (rapidly) after SSZ withdrawal and SSZ re-exposure, respectively. Beyond this, CEM/SSZ cells were characterised by enhanced sensitivity to the DMARDs cyclosporin A and chloroquine, and most notably to dexamethasone, which may provide possible strategies to circumvent resistance to SSZ.
SSZ is commonly used in various DMARD regimens for RA treatment, including monotherapy or combination therapy with other DMARDs such as MTX, hydroxychloroquine or in the COBRA regimen.19,20,35,40 SSZ can induce antiproliferative/apoptotic effects41 as well as anti-inflammatory effects by inhibiting activation of NFκB,37 which leads to the decreased production/secretion of proinflammatory cytokines such as TNFα.38 Although SSZ is a potent DMARD, its efficacy in long term treatment seems to be more compromised than that of other DMARDs, such as MTX.14,16,18 Because lack of efficacy may be related to either persistent/renewed disease activity and/or side effects of DMARDs, we suggested that diminished SSZ efficacy might also be associated with the development of resistance. Indeed, as previously reported,22 we found that the onset of SSZ resistance in human CEM (T) cells coincided with the induction of the MDR pump ABCG2/BCRP. Thus far, ABCG2/BCRP induction had only been noted upon development of a selected group of topoisomerase inhibitors such as mitoxantrone, doxorubicin, and topotecan.12,34,42–44
ABCG2/BCRP expression in CEM/SSZ cells did not fully account for the resistant phenotype as about 50% of the SSZ resistance was lost within one week after withdrawal of SSZ, whereas ABCG2/BCRP expression was unchanged. The nature of this latter component has not been identified, but may be related to SSZ-induced transient alterations in the NFκB signalling pathway38 that controls the transcription of anti-apoptotic and proinflammatory cytokine/chemokine genes.45–47 Consistent with this hypothesis is the observation of a two- to threefold enhancement of TNFα production/secretion by CEM/SSZ cells compared with CEM (T) cells.22 It remains to be established whether this type of SSZ resistance-induced effect may also be responsible for the altered antiproliferative effects (table 1) of NFκB signalling pathway drugs, including leflunomide,48 MG132,39 and dexamethasone.45,49,50
The gradual loss of ABCG2/BCRP expression from cell cultures of CEM/SSZ1.5 cells after 6 months’ (approximately 50 passages) withdrawal of SSZ is consistent with a study by Maliepaard et al, who noted the loss of ABCG2/BCRP expression from a topotecan resistant ovarian carcinoma cell line over 30 passages in drug-free medium.51 These results indicate that ABCG2/BCRP expression gradually declines when the selective pressure is absent, but can be rapidly resumed upon renewed exposure to the selective drug.
Besides the effects on ABCG2/BCRP expression, it is of interest to note that SSZ exposure/resistance had down regulatory effects on the expression of ABCC1/MRP1. Although this MDR transporter has an established role in extrusion of various toxic drugs,2,5,11 it also has important immunological functions in, for instance, dendritic cells, where, by exporting the cysteinyl leucotriene LTC4, it mediates the signalling for chemokine CCL19 chemotaxis and migration of dendritic cells to the lymph nodes.52 Because SSZ is an inhibitor of LTC synthetase,53 impaired synthesis of LTC may parallel reduced expression of its transporter ABCC1/MRP1. Another example of ABCC1/MRP1 down/up regulation was recently identified by our laboratory after depletion/repletion of cellular folate status54 in CEM (T) cells. Consistent with the notion that SSZ can also exert antifolate effects,55 we obtained preliminary evidence that the differential expression of ABCC1/MRP1 is indeed correlated with the cellular folate status in CEM/SSZ cells (Jansen G, Scheper RJ, Dijkmans BAC, unpublished data).
The present study also provides insight into strategies that may or may not be successful to circumvent/reverse SSZ resistance. Firstly, temporary discontinuation of DMARD treatment after an initial response and resumption of the treatment at the time of progressive disease proved to be effective.56 Whether a similar strategy is also effective for patients with RA for whom DMARD treatment has failed, has not been investigated. When such a clinical strategy was mimicked in vitro—discontinuation of SSZ treatment after development of SSZ resistance and rechallenging with SSZ at the time SSZ resistance had apparently disappeared—it did not seem to be effective. SSZ resistance, along with ABCG2/BCRP overexpression, was rapidly reinduced (<2.5 weeks). Secondly, development of resistance to SSZ may be accompanied by adverse effects of collateral resistance to other DMARDs (for example, leflunomide) which might be used for sequential monotherapy. Potentially positive strategies to target SSZ resistance may include the use of (a) blockers of ABCG2/BCRP that serve as chemosensitisers to reverse SSZ resistance (for example, Ko143); (b) DMARDs whose antiproliferative activity was not impaired during development of SSZ resistance, such as cyclosporin A and chloroquine. Because chloroquine is a substrate for efflux by ABCC1/MRP1, a reduced expression of ABCC1/MRP1, as seen in CEM/SSZ cells, may confer enhanced sensitivity to chloroquine,57 which could increase the efficacy of hydroxychloroquine containing treatments.20 Finally, most interesting was the observation that CEM/SSZ cells displayed hypersensitivity (13-fold) to the glucocorticoid dexamethasone. One previous report has described collateral sensitivity to dexamethasone (10-fold) in human myeloma cells with acquired resistance to doxorubicin.58 The mechanistic basis for the markedly enhanced dexamethasone sensitivity is not clear, but is not associated with an increase in glucocorticoid receptor levels in CEM/SSZ cells compared with CEM (T) cells (not shown).
Altogether, this study shows that exposure/resistance to SSZ can have differential effects on the expression of at least two important MDR pumps: ABCG2/BCRP and ABCC1/MRP1. As these MDR pumps are normally expressed on various peripheral blood cell types,59–61 further research is warranted to identify the expression levels of these MDR pumps during treatment of patients with RA with DMARDs which include SSZ. Thus far, only a few studies have investigated blood samples of patients with RA for differential expression of the MDR transporter Pgp.62–65 Although these studies included limited numbers of patients, data were supportive for a role of Pgp in DMARD efficacy. Our present study, though based on in vitro data using a T cell model system, demonstrates also that MDR pumps other than Pgp may be relevant for DMARD resistance. Once specific MDR proteins are identified for their contributing role in DMARD (in)efficacy, this information may be exploited to use specific blockers for the various MDR pumps2 as chemosensitisers for DMARD activity.
Acknowledgments
J van der Heijden is a recipient of the 2002 Arthron Young Investigator Fellowship.
This study is supported by a grant (NR03-1-402) from the Dutch Rheumatism Association (Nationaal Reumafonds).