


Visual Overview

Abstract
The common marmoset (Callithrix jacchus), a small New World monkey, has the potential for use in human drug development due to its evolutionary closeness to humans. Four novel cDNAs, encoding cytochrome P450 (P450) 2C18, 2C19, 2C58, and 2C76, were cloned from marmoset livers to characterize P450 2C molecular properties, including previously reported P450 2C8. The deduced amino acid sequence showed high sequence identities (>86%) with those of human P450 2Cs, except for marmoset P450 2C76, which has a low sequence identity (∼70%) with any human P450 2Cs. Phylogenetic analysis showed that marmoset P450 2Cs were more closely clustered with those of humans and macaques than other species investigated. Quantitative polymerase chain reaction analysis showed that all of the marmoset P450 2C mRNAs were predominantly expressed in liver as opposed to the other tissues tested. Marmoset P450 2C proteins were detected in liver by immunoblotting using antibodies against human P450 2Cs. Among marmoset P450 2Cs heterologously expressed in Escherichia coli, marmoset P450 2C19 efficiently catalyzed human P450 2C substrates, S-warfarin, diclofenac, tolbutamide, flurbiprofen, and omeprazole. Marmoset P450 2C19 had high Vmax and low Km values for S-warfarin 7-hydroxylation that were comparable to those in human liver microsomes, indicating warfarin stereoselectivity similar to findings in humans. Faster in vivo S-warfarin clearance than R-warfarin after intravenous administration of racemic warfarin (0.2 mg/kg) to marmosets was consistent with the in vitro kinetic parameters. These results indicated that marmoset P450 2C enzymes had functional characteristics similar to those of humans, and that P450 2C–dependent metabolic properties are likewise similar between marmosets and humans.
Introduction
The cytochrome P450 (P450) superfamily comprises 57 functional genes and 58 pseudogenes in humans (Nelson et al., 2004) and encodes enzymes mainly involved in the oxidative metabolism of drugs, toxic chemicals, and endogenous compounds. The human P450 2C enzymes, including P450 2C8, 2C9, and 2C19, metabolize 24% of all pharmaceutical drugs, such as diazepam, ibuprofen, phenytoin, tolbutamide, warfarin, and omeprazole (Rendic and Guengerich, 2014).
Due to their genetic similarity to humans, nonhuman primates are used for preclinical studies to predict the toxicity and pharmacokinetics of new drugs. The Old World monkeys such as cynomolgus monkeys (Macaca fascicularis) and rhesus monkeys (Macaca mulatta) are the most commonly used among nonhuman primates. The common marmoset (Callithrix jacchus), another New World monkey, is beneficial in various research fields such as neuroscience, stem cell research, drug metabolism, toxicology, and immune and autoimmune diseases (Kishi et al., 2014; Tokuno et al., 2015) because of its small body size, availability, high reproductive efficiency, and early sexual maturity (Sasaki, 2015).
In the marmoset P450 2C subfamily, P450 2C8 cDNA has been identified and characterized. Marmoset P450 2C8 shares high sequence identity (87%) with human P450 2C8 and catalyzes tolbutamide methyl hydroxylation (Narimatsu et al., 2006). In cynomolgus monkeys, five P450 2C cDNAs (namely, P450 2C8, 2C9, 2C18, 2C19, and 2C76) have been identified (Uno et al., 2011a). Monkey P450 2C9 and 2C19 share similar metabolic properties with human P450 2C9 and 2C19 in the metabolism of tolbutamide, S-mephenytoin, flurbiprofen, and diclofenac (Uno et al., 2011b). P450 2C19 stereoselectively catalyzes R-warfarin 7-hydroxylation (Hosoi et al., 2012). Cynomolgus monkey P450 2C76, not orthologous to human P450s, catalyzes tolbutamide methyl hydroxylation and is partly responsible for the differences in drug metabolism between macaques and humans (Uno et al., 2011a). P450 2C93 is a functional enzyme in rhesus monkeys, but not in cynomolgus monkeys (Uno et al., 2011b).
We previously reported gene expressions of marmoset P450 1A, 2A, 2B, 2C, 2D, 2E, 2J, and 3A in livers (Shimizu et al., 2014) and enzymatic characteristics of marmoset P450 3A, 2D, and 2A isoforms (Uehara et al., 2015a,b,c). The characterization of marmoset P450 enzymes is useful for understanding the metabolic properties of marmosets. In the present study, we report the identification of four novel P450 2C cDNAs isolated from marmoset livers. Four novel marmoset P450 2Cs and previously reported P450 2C8 (Narimatsu et al., 2006) were analyzed for their amino acid sequence identity, tissue distribution of mRNA expression, and drug-metabolizing activity compared with human and/or cynomolgus monkey P450 2Cs.
Materials and Methods
Chemicals and Enzymes.
Ethoxyresorufin, resorufin, 7-hydroxycoumarin, 7-pentoxyresorufin, diclofenac sodium, 4′-hydroxydiclofenac tolbutamide, flurbiprofen, S/R-warfarin, 7-hydroxywarfarin, chlorzoxazone, 6-hydroxychlorzoxazone, and 1′-hydroxymidazolam were purchased from Sigma-Aldrich (Tokyo, Japan). Coumarin, 7-hydroxycoumarin, paclitaxel, omeprazole, and midazolam were purchased from Wako Pure Chemicals (Osaka, Japan). 6α-Hydroxypaclitaxel, methylhydroxy tolbutamide, and bufuralol were purchased from Corning Life Sciences (Woburn, MA). 4′-Hydroxyflurbiprofen, 5-hydroxyomeprazole, and 1′-hydroxybufuralol were purchased from Toronto Research Chemicals (Toronto, Canada). Oligonucleotides were synthesized by Greiner Japan (Tokyo, Japan). Pooled liver microsomes from marmosets, cynomolgus monkeys, and humans were purchased from Corning Life Sciences. Marmoset liver microsomes were also prepared from individual marmoset tissue samples as described previously (Uehara et al., 2014). The other chemicals were obtained in the highest grade commercially available.
Animals and Tissue or Plasma Preparations.
Four marmosets (males, >2 years old) were purchased from CLEA Japan (Tokyo, Japan). The animals were kept in cages (40 × 610 × 1578 mm) at 24–27°C, 40–60% relative air humidity with a 12/12-hour light/dark cycle, and had free access to a balanced diet (CMS-1M; CLEA Japan) with added vitamins and water. This study was approved by the animal ethics committee and the gene recombination experiment safety management committee of the Central Institute for Experimental Animals, and was carried out according to the Guidelines for Proper Conduct of Animal Experiments by the Science Council of Japan (2006). Animal care was conducted in accordance with the recommendation of the Guide for the Care and Use of Laboratory Animals (National Research Council of the National Academies, 2011). Tissue samples including brain, lung, liver, kidney, and small intestine samples were collected from 16 marmosets (9 males and 7 females) after euthanasia by exsanguination under ketamine (60 mg/kg) and isoflurane deep anesthesia as described previously (Shimizu et al., 2014).
Stocked marmoset plasma samples (Uehara et al., 2015a), after intravenous administration of five-drug composites of caffeine, warfarin, omeprazole, metoprolol, and midazolam (cassette dosing) at doses of 0.20 mg kg−1 each, were reanalyzed for S/R-warfarin using liquid chromatography–tandem mass spectrometry [Nexera UHPLC System (Shimadzu, Kyoto, Japan); TSQ Vantage (Thermo Fisher Scientific Inc., Waltham, MA)] as described previously (Utoh et al., 2015). In brief, S/R-warfarin was determined separately using a Chiralcel OD-RH column (Daicel, Tokyo, Japan) with 6% (v/v) methanol and 54% (v/v) acetonitrile in 0.040% (v/v) formic acid for 7 minutes. The mass transitions (m/z) monitored were 307 → 250 for S/R-warfarin.
Sequence Analysis.
Methods for cDNA cloning are shown in the Supplemental Methods. The amino acid sequences of marmoset P450 2Cs were compared with known P450 2C amino acid sequences of various species from GenBank using BLAST (National Center for Biotechnology Information). Multiple alignment of amino acid sequences was performed using Genetyx (Software Development, Tokyo, Japan). Analyses of the marmoset, rhesus monkey, and human genome data were performed using BLAST-like alignment tool (BLAT). The phylogenic tree was constructed by the neighbor-joining method using DNASIS Pro (Hitachi Software, Tokyo, Japan). The P450 2C amino acid sequences from GenBank were used, including marmoset P450 2C8 (NP_001191366); human P450 2A6 (NP_000753), 2C8 (NP_000755), 2C9 (NP_000762), 2C18 (NP_000763), and 2C19 (NP_000760); cynomolgus monkey P450 2C8 (NP_001270692), 2C9 (NP_001274554), 2C18 (ABB87194), 2C19 (NP_001270219), and 2C76 (NP_001271718); rhesus monkey P450 2C8 (NP_001035300), 2C9 (NP_001035329), 2C18 (NP_001180995), 2C19 (NP_001035301), 2C76 (NP_001171259), and 2C93 (NP_001232885); dog P450 2C21 (NP_001183973) and 2C41 (NP_001003334); rabbit P450 2C1 (P00180), 2C2 (NP_001164584), 2C3 (NP_001164736), 2C4 (NP_001177360), 2C5 (NP_001164397), 2C14 (NP_001164591), 2C16 (NP_001164593), and 2C30 (NP_001164737); pig P450 2C33 (NP_999579), 2C42 (NP_001161307), and 2C49 (NP_999585); rat P450 2C6 (NP_001013926), 2C7 (NP_058854), 2C11 (NP_062057), 2C12 (NP_113760), 2C13 (NP_612523), 2C22 (NP_612521), 2C23 (NP_114027), and 2C24 (NP_001258283); and mouse P450 2C29 (NP_031841), 2C37 (NP_034131), 2C38 (NP_034132), 2C39 (NP_034133), 2C40 (NP_034134), 2C44 (NP_001001446), 2C50 (NP_598905), 2C54 (NP_996260), 2C55 (NP_082365), 2C65 (NP_082467), 2C66 (NP_001011707), and 2C70 (NP_663474). The marmoset P450 2C18, 2C19, 2C58, and 2C76 amino acid sequences were deduced from the cDNAs identified in this study.
Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction.
The mRNA expressions of marmoset P450 2C genes in brain, lung, liver, kidney, and small intestine were measured by quantitative real-time reverse transcription (RT)-polymerase chain reaction (PCR). RNA isolation and cDNA synthesis were carried out as described earlier. In brief, total RNA samples were pooled in equal quantities from six marmosets for each tissue. The RT samples were prepared in a total volume of 20 μl containing 1 μg of pooled total RNA, SuperScript III RT reverse transcriptase, and random primers (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. PCR amplification was performed in triplicate with a total volume of 20 μl using Fast SYBR Green Master Mix (Applied Biosystems, Foster City, CA) on an ABI PRISM 7500 sequence detection system (Applied Biosystems). The PCR primers were used at a final concentration of 200 nM, including marmoset CYP2C8 (cjCYP2C8; 5qrt1) 5′-CAGTAAAGGAAGCCCTGATTGATA-3′ and cjCYP2C8 (3qrt1) 5′-ATTGCTGGAAATGATTCCAAGAT-3′ for P450 2C8, cjCYP2C19L (5qrt1) 5′-CAGAGATACATTGACCTCATTCCTACTAAC-3′ and cjCYP2C19L (3qrt1) 5′-AGGCCCTCTCCTACACACATC-3′ for P450 2C18, cjCYP2C26L (5qrt1) 5′-ACCAAGAATCGGTAGACATTAACAAC-3′ and cjCYP2C26L (3qrt1) 5′-TCCAAACAAGTCTAATGCAGTGTTTAC-3′ for P450 2C19, cjCYP2C20L (5qrt1) 5′-GGCAACTTTAAGAAAAGTGACCAT-3′ and cjCYP2C20L (3qrt1) 5′-GGGTGGCAAGGAAAAAATCC-3′ for P450 2C58, and cjCYP2C21L (5qrt1) 5′-AAAGGGCACAACAATATTAGCAGAT-3′ and cjCYP2C21L (3qrt1) 5′-GAACTGGGGTGGTGTCAATATC-3′ for P450 2C76. PCR cycling conditions were 2 minutes at 50°C and 10 minutes at 95°C, followed by 40 cycles at 95°C for 15 seconds and at 60°C for 1 minute. A calibration curve was generated using each target marmoset P450 2C cDNA, and the amplification efficiency was confirmed. Marmoset P450 2C mRNA expression levels were normalized with 18S ribosomal RNA level.
Expression and Preparation of Recombinant P450 2Cs in Escherichia coli.
Marmoset P450 2Cs were expressed by a heterologous expression system in Escherichia coli using the methods described previously (Uehara et al., 2010). In brief, to enhance protein expression, marmoset P450 2C cDNAs were modified at the N terminus by PCR using forward and reverse primers containing the restriction sites of the NdeI and XbaI sites (underlined), respectively. PCR primers were cjCYP2C8/20L (5exp1bov) 5′- GGAATTCCATATGGCTCTGTTATTAGCAGTTTTTCTCTGTCTCTCTTTTTTGCTTCTCTTTTCAC-3′ and cjCYP2C8 (3exp1) 5′- GCTCTAGATCAGACAGGAATGAAGCAGATCTGGTA-3′ for P450 2C8/2C58, cjCYP2C19L (5exp1bov) 5′- GGAATTCCATATGGCTCTGTTATTAGCAGTTTTTTTCTGTCTCTCCTGTTTGCTTCTCCTT-3′ and cjCYP2C19L (3exp1) 5′- GCTCTAGATCAGACTGGAATGAAGCAGAGCTGGTA-3′ for P450 2C18, cjCYP2C26L (5exp1bov) 5′- GGAATTCCATATGGCTCTGTTATTAGCAGTTTTTCTCTCTCTCTCCTGTTTGCTTCTCCTT-3′ and cjCYP2C26L (3exp1) 5′- GCTCTAGATCTTCAGATAGGAATGAAGCACAACTGAT-3′ for P450 2C19, and cjCYP2C21L (5exp1bov) 5′- GGAATTCCATATGGCTCTGTTATTAGCAGTTTTTACTTGTCTTTCTTGTCTGACTCTTCTTTTTCTG-3′ and cjCYP2C21L (3exp1) 5′- GCTCTAGATCATCAGACTGGAATAAAACAAAGCTCATAG-3′ for P450 2C76. PCR products were purified, double digested, and ligated with the pCW vector, which contained the human NADPH-P450 reductase cDNA, and sequences of the inserts were confirmed by sequencing. Plasmids for expression of human P450 2C8, 2C9, and 2C19 and cynomolgus monkey P450 2C19 were constructed as described previously (Yamazaki et al., 2002; Uno et al., 2006). The membrane fractions were prepared from E. coli DH5α expressing P450 2C proteins: the concentrations of P450 protein and NADPH-P450 reductase were measured as described previously (Yamazaki et al., 2002). The detection of P450 2C proteins in recombinant and marmoset liver microsomes was performed by immunoblotting as shown in the Supplemental Methods.
Activity Characterization and Kinetic Analysis.
Ethoxyresorufin O-deethylation, 7-ethoxycoumarin O-deethylation, coumarin 7-hydroxylation, 7-pentoxyresorufin O-deethylation, paclitaxel 6α-hydroxylation, diclofenac 4′-hydroxylation, tolbutamide methylhydroxylation, flurbiprofen 4-hydroxylation, S/R-warfarin 7-hydroxylation, omeprazole 5-hydroxylation, chlorzoxazone 6-hydroxylation, bufuralol 1′-hydroxylation, and midazolam 1′-hydroxylation by recombinant P450 2C proteins and liver microsomes from marmosets, cynomolgus monkeys, and humans were measured as described previously (Yamazaki et al., 2002; Uno et al., 2011b) with some minor changes. In brief, each mixture containing recombinant protein (20 pmol/ml) or liver microsomes (0.40 mg/ml), a NADPH-generating system (0.25 mM NADP+, 2.5 mM glucose 6-phosphate, and 0.25 units/ml glucose 6-phosphate dehydrogenase), and substrate (10 μM ethoxyresorufin, 100 μM 7-ethoxycoumarin, 100 μM coumarin, 10 μM 7-pentoxyresorufin, 100 μM paclitaxel, 100 μM diclofenac, 2500 μM tolbutamide, 100 μM flurbiprofen, 100 μM S- and R-warfarin, 100 μM omeprazole, 500 μM chlorzoxazone, 100 μM bufuralol, or 100 μM midazolam) in 100 mM potassium phosphate buffer (pH 7.4) was incubated at 37°C for the optimal time (10 minutes for ethoxyresorufin 7-ethoxycoumarin, coumarin, 7-pentoxyresorufin, and midazolam; 15 minutes for paclitaxel, tolbutamide, diclofenac, flurbiprofen, S- and R-warfarin, omeprazole, chlorzoxazone, and bufuralol). For metabolites from 7-ethoxycoumarin, coumarin, flurbiprofen, S- and R-warfarin, bufuralol, and midazolam, reactions were stopped by addition of 25 μl of 60% HClO4 (w/v) or 0.4 ml of methanol. Reaction mixtures were centrifuged at 10,000g for 5 minutes, and metabolites of supernatants were analyzed directly by high-performance liquid chromatography (HPLC) with fluorescence or a UV detector. For metabolites of other substrates, reactions were stopped with ethyl acetate and centrifuged at 2500g for 5 minutes. The organic phases were evaporated and the residues were dissolved in the HPLC mobile phase, and then subjects were analyzed by HPLC with a UV detector. For kinetic analysis, reactions were performed over a range of concentrations for each substrate (5–400 μM paclitaxel, 5–2500 μM tolbutamide, 1–800 μM S/R-warfarin, 0.5–300 μM flurbiprofen, and 1–1000 μM omeprazole). Data were fitted to an integrated form of the Michaelis-Menten equation using KaleidaGraph (Synergy Software, Reading, PA).
Results
Identification of Novel Marmoset P450 2C Genes.
To identify marmoset P450 2C cDNAs except for P450 2C8, which was reported previously (Narimatsu et al., 2006), a search for the sequences highly homologous to human P450 2C cDNAs was performed on the marmoset genome data using BLAT. To clone the four novel marmoset P450 2C cDNAs, along with P450 2C8, RT-PCR was performed with total marmoset liver RNA to amplify the region containing the open reading frame of marmoset P450 2C cDNA. Marmoset P450 2C18, 2C19, 2C58, and 2C76 cDNAs, encoding the proteins of 490 amino acids for P450 2C18, 2C19, and 2C58 or 489 amino acids for P450 2C76, were obtained (Fig. 1). The deduced amino acids of all marmoset P450 2C cDNAs contained the six potential substrate recognition sites and heme-binding region. Marmoset P450 2C18, 2C19, and 2C58 showed high sequence identities to human P450 2C18 (93%), 2C9/2C19 (86%), and 2C8 (87%) at the amino acid level (Table 1). Marmoset P450 2C76 showed high sequence identity (87%) with cynomolgus monkey P450 2C76, but low sequence identity with human P450 2Cs (68–70%).

Multiple sequence alignment of the deduced amino acid sequences of marmoset P450 2Cs. Amino acid sequences of marmoset P450 2Cs (2C8, 2C18, 2C19, 2C58, and 2C76) and human (h) P450 2Cs (2C8, 2C9, 2C18, and 2C19) were aligned using Genetyx. Six substrate recognition sites (SRSs) are marked by a solid line. The heme-binding domain is marked by a broken line. An asterisk indicates perfectly conserved amino acids. A dot indicates conservatively changed amino acids.
Similarities of the deduced amino acid sequences of marmoset P450 2Cs with cynomolgus monkey and human P450 2C isoforms
The BLAST program was used to compare sequences with marmoset, cynomolgus monkey, and human P450 2C isoforms as described in Materials and Methods.
Phylogenetic analysis of P450 2C amino acid sequences in marmosets and other species indicated that marmoset P450 2C18, 2C19, and 2C58 were closely clustered with human P450 2Cs, together with cynomolgus monkey and rhesus monkey P450 2Cs, unlike dog, pig, rabbit, rat, and mouse P450 2Cs (Fig. 2), whereas marmoset P450 2C76 was closely clustered with cynomolgus monkey and rhesus monkey P450 2C76.

Phylogenetic tree of marmoset P450 2C amino acid sequences. The neighbor-joining method was used to create a phylogenetic tree with P450 2C amino acid sequences of marmoset (cj), human (h), cynomolgus monkey (mf), rhesus monkey (mm), dog (d), pig (p), rabbit (rab), rat (r), and mouse (m), together with human P450 2A6 as an outgroup. Branched length is proportional to the number of amino acid substitutions. The scale bar indicates 10% amino acid substitution in the sequence.
The genomic location of the P450 2C genes in the marmoset genome was determined by analyzing the marmoset genome data using BLAT, together with the macaque (rhesus monkey) and human genomes. The analysis showed that P450 2C8, 2C18, 2C19, 2C58, and 2C76 formed the P450 2C cluster in marmoset chromosome 12, indicating that the P450 2C cluster was conserved among marmosets, macaques, and humans with some differences (Fig. 3). In the macaque genome, seven genes, P450 2C8, 2C9, 2C18, 2C19, 2C58P, 2C76, and 2C93, were found, whereas five genes, P450 2C8, 2C9, 2C18, 2C19, and 2C58P, were located in the human genome. P450 2C58 was the pseudogene in human and macaque, but not in marmoset. In the marmoset and macaque genomes, P450 2C76 was found in the location corresponding to the intergenic region in the human genome.
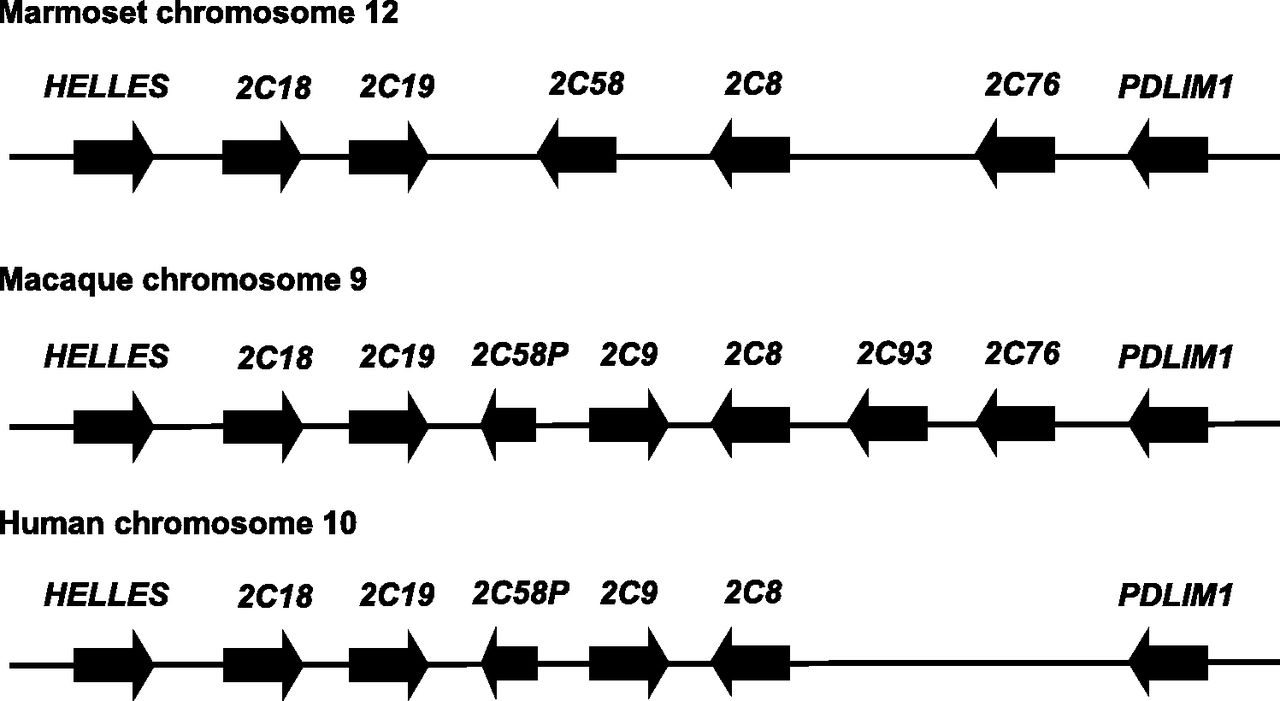
Genomic structures of P450 2C cluster in marmosets, macaques, and humans. The genomic structures were analyzed using the marmoset, macaque (rhesus monkey), and human genome data on BLAT. P450 2C genes were located between HELLES and PDLIM1 in the marmoset genome, similar to rhesus monkey and human. The length of the genes and the distance between the genes are not proportionate to actual measurement.
Tissue Expression Pattern of Marmoset P450 2Cs.
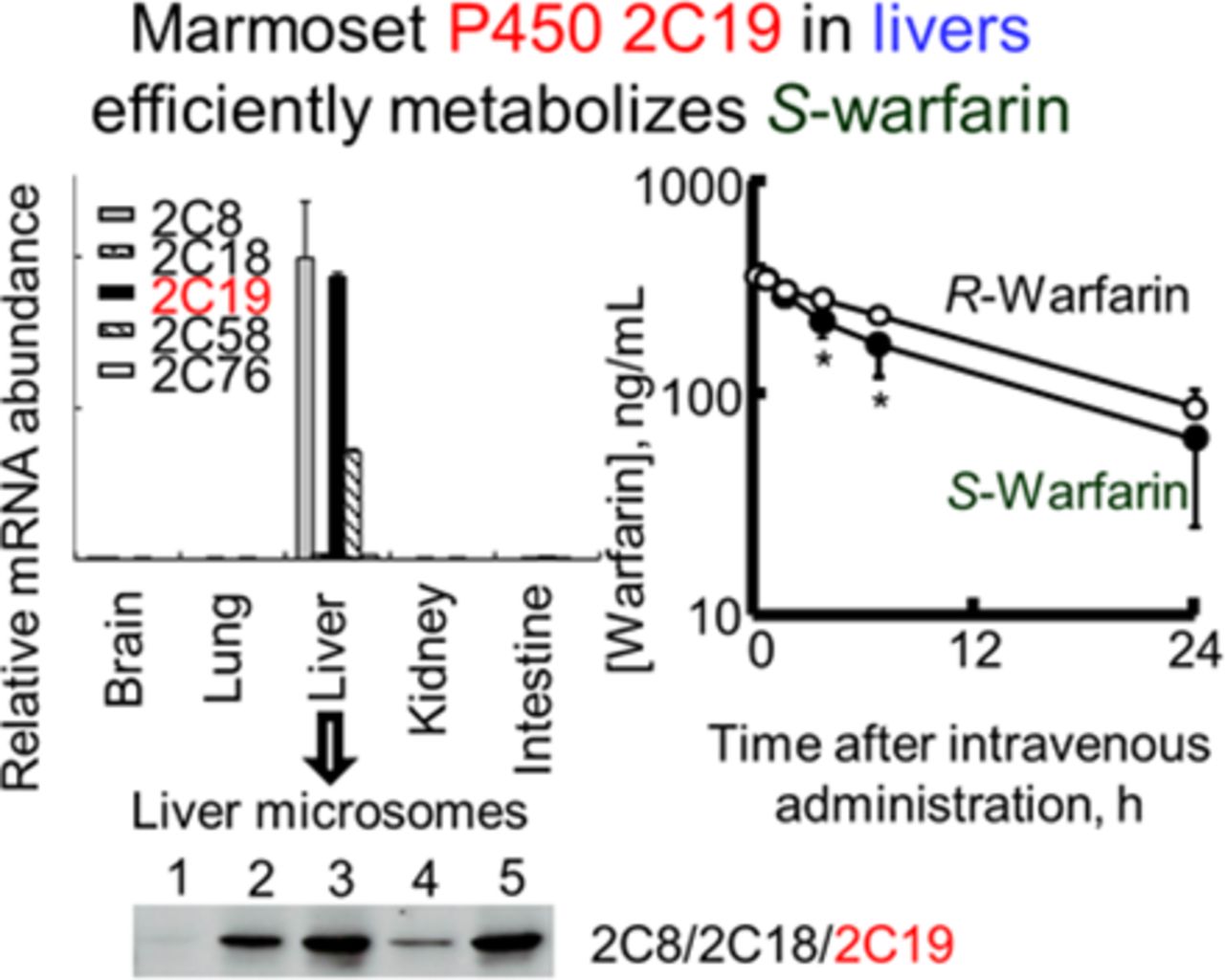
To measure the expression level of P450 2C mRNAs in marmoset tissues, real-time RT-PCR was performed with gene-specific primers using total RNA of brains, kidneys, livers, lungs, and small intestines. Among the five tissues examined, all five marmoset P450 2C mRNAs were expressed predominantly in the liver, where P450 2C8 mRNA was most abundant, followed by P450 2C19, 2C58, 2C18, and 2C76 mRNA (Fig. 4).

Expression of P450 2C mRNAs in marmoset tissues. Expression of marmoset P450 2C mRNAs in brains, lungs, livers, kidneys, and small intestines was measured by real-time RT-PCR. The raw data were normalized to 18S ribosomal RNA expression level. Values are presented as the means plus standard deviations (n = 3). Expression level of P450 2C8 mRNA was adjusted to 1, and expression levels of other P450 2C mRNAs are shown as the expression values relative to P450 2C8 mRNA.
Expression of marmoset P450 2C proteins in livers was confirmed by immunoblotting using anti-human or anti-cynomolgus monkey P450 2C antibodies. The cross-reactivity of each P450 2C antibody was investigated using recombinant proteins, including marmoset P450 2C8, 2C18, 2C19, 2C58, and 2C76 (Supplemental Fig. 1A). Recombinant marmoset P450 2C8 and 2C58; 2C8, 2C18, and 2C19; and 2C76 were detected by anti-human P450 2C8, anti-human P450 2C9, and anti-cynomolgus monkey P450 2C76 antibodies, respectively. Immunoblot analysis using five individual marmoset liver microsomes detected bands of approximately 56 kDa (Supplemental Fig. 1B), indicating that marmoset P450 2C proteins were expressed in marmoset liver with a large individual variation.
Kinetic Analysis for Drug Oxidations Mediated by Marmoset P450 2Cs.
To investigate catalytic function of marmoset P450 2C enzymes, ethoxyresorufin O-deethylation, 7-ethoxycoumarin O-deethylation, coumarin 7-hydroxylation, 7-pentoxyresorufin O-deethylation, paclitaxel 6α-hydroxylation, diclofenac 4′-hydroxylation, tolbutamide methyl hydroxylation, flurbiprofen 4-hydroxylation, S/R-warfarin 7-hydroxylation, omeprazole 5-hydroxylation, chlorzoxazone 6-hydroxylation, bufuralol 1′-hydroxylation, and midazolam 1′-hydroxylation were measured using recombinant marmoset P450 2C enzymes (Table 2). Marmoset P450 2C enzymes substantially metabolized human P450 2C probe substrates, paclitaxel (P450 2C8), diclofenac (P450 2C18, 2C19, and 2C76), tolbutamide (P450 2C8, 2C18, 2C19, 2C58, and 2C76), flurbiprofen (P450 2C8, 2C19, and 2C58), S- and R-warfarin (P450 2C18 and 2C19), and omeprazole (P450 2C19). Marmoset P450 2C enzymes metabolized other non-P450 2C human substrates, ethoxyresorufin, 7-ethoxycoumarin, and 7-pentoxyresorufin (marmoset P450 2C8, 2C18, 2C19, 2C58, and 2C76); coumarin and chlorzoxazone (P450 2C8 and 2C58); and bufuralol (P450 2C8, 2C18, 2C19, and 2C58).
Catalytic activities by recombinant marmoset P450 2C enzymes
Kinetic analysis for paclitaxel 6α-hydroxylation, tolbutamide methyl hydroxylation, flurbiprofen 4-hydroxylation, and omeprazole 5-hydroxylation was performed with recombinant P450 2C enzymes and liver microsomes from marmosets and humans (Table 3). Marmoset P450 2C8 indicated low affinity (Km, 42 μM) and capacity (Vmax, 1.6 min−1) for paclitaxel 6α-hydroxylation, showing a lower Vmax/Km value (0.038 μM−1 min−1) than that of human P450 2C8 (7.2 μM−1 min−1). Marmoset liver microsomes showed an apparent Km value of 44 μM toward paclitaxel 6α-hydroxylation, similar to marmoset P450 2C8. For tolbutamide methyl hydroxylation, marmoset P450 2C19 showed an apparent Km (217 μM) lower than that of marmoset P450 2C8 (2210 μM) and 2C58 (1320 μM) and human P450 2C9 (284 μM), resulting in a Vmax/Km value (0.092 μM−1 min−1) lower than that of human P450 2C9 (0.19 μM−1 min−1). Marmoset liver microsomes showed an apparent Km value of 373 μM toward tolbutamide methyl hydroxylation, similar to marmoset P450 2C19. For flurbiprofen 4-hydroxylation, marmoset P450 2C8, 2C19, and 2C58 showed apparent Km values of 155, 161, and 85 μM, and Vmax/Km values of 0.0090, 0.075, and 0.0094 μM−1 min−1, respectively, indicating low affinity and Vmax/Km values of marmoset P450 2C enzymes for flurbiprofen 4-hydroxylation compared with human P450 2C9 (Km, 15 μM; Vmax/Km, 1.8 μM−1 min−1). Marmoset liver microsomes also showed an apparent Km value of 34 μM and a Vmax/Km value of 0.0021 μM−1 min−1 compared with those of human liver microsomes. For omeprazole 5-hydroxylation, marmoset P450 2C19 showed an apparent Km value of 23 μM and Vmax/Km value of 2.3 μM−1 min−1, similar to human P450 2C9. Marmoset liver microsomes showed a lower Km value of 30 μM toward omeprazole 5-hydroxylation, similar to marmoset P450 2C19.
Kinetic parameters for oxidation activities of paclitaxel, tolbutamide, flurbiprofen, and omeprazole by recombinant P450 2C enzymes and liver microsomes from marmosets and humans
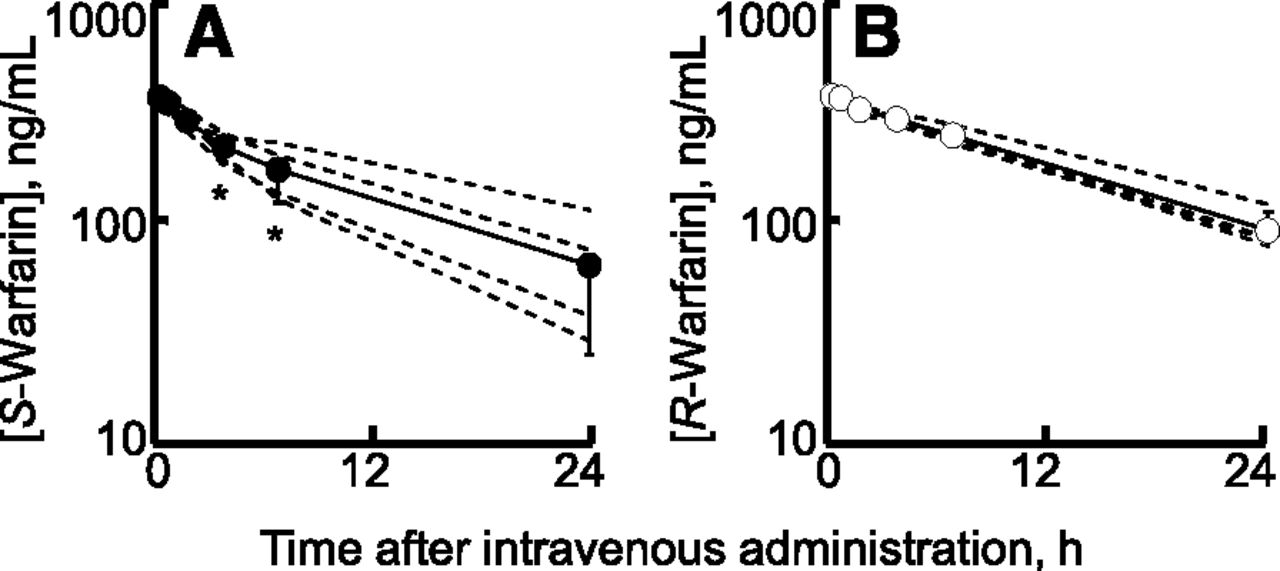
S/R-warfarin oxidation was further analyzed both in vitro (Table 4) and in vivo (Fig. 5) in marmosets. Recombinant marmoset P450 2C19 had a higher affinity (Km, 14 μM) and Vmax/Km (14 mM−1 min−1) for S-warfarin 7-hydroxylation than those for R-warfarin 7-hydroxylation (Km, 755 μM; Vmax/Km, 0.26 mM−1 min−1), but was comparable to those for marmoset liver microsomes (Table 4). Similarly, human P450 2C9 had a low Km value of 3.1 μM and a high Vmax/Km value of 55 mM−1 min−1 for S-warfarin 7-hydroxylation compared with those for R-warfarin 7-hydroxylation (Km, 123 μM; Vmax/Km, 0.057 mM−1 min−1). By contrast, cynomolgus monkey P450 2C19 had a high Km value of 124 μM and a low Vmax/Km value of 10 mM−1 min−1 for S-warfarin 7-hydroxylation compared with those for R-warfarin 7-hydroxylation (Km, 51 μM; Vmax/Km, 137 mM−1 min−1). To investigate the pharmacokinetics of S- and R-warfarin in marmosets, plasma concentrations of S- and R-warfarin after a single intravenous administration at doses of 0.20 mg racemic warfarin kg−1 each to four male marmosets were measured using liquid chromatography–tandem mass spectrometry. Plasma concentrations of S-warfarin decreased significantly faster than R-warfarin in a monophasic manner in marmosets (Fig. 5). Calculated mean clearance values (and ranges) in four marmosets for S- and R-warfarin were 22.2 (13.0–29.8) and 15.3 (12.8–17.0) ml/h/kg, respectively.
Kinetic parameters for 7-hydroxylation of racemic, S-, and R-warfarin by recombinant P450 2C enzymes and liver microsomes from marmosets, cynomolgus monkeys, and humans

Plasma concentrations of S- and R-warfarin after intravenous administration of racemic warfarin (0.2 mg/kg) in marmosets. Plasma concentrations of S-warfarin (A) and R-warfarin (B) after intravenous administration of racemic warfarin (0.2 mg/kg) were measured in four marmosets. The average and individual values are marked by solid and broken lines, respectively. Results are expressed with circles (means) and bars (±S.D.s) obtained with four marmosets (*P < 0.05, two-way analysis of variance with Bonferroni post test).
Discussion
Human P450 2C enzymes in liver account for the metabolism of approximately 20% of the drugs on the market, and thus P450 2C enzymes are important in drug metabolism. Despite the potential importance of marmosets in drug development, marmoset P450 2C enzymes have not been fully identified and characterized. We previously reported that a number of P450 2C genes were expressed in marmoset livers (Shimizu et al., 2014). In the present study, four novel P450 2C cDNAs were cloned from marmoset livers (Figs. 1 and 3) based on the marmoset genome data. Marmoset P450 2C18, 2C19, and 2C58 had high sequence identities (>86%) at the amino acid level with human P450 2Cs (Table 1), whereas marmoset P450 2C76 had a high sequence identity (87%) with cynomolgus monkey P450 2C76. In phylogenetic analysis, marmoset P450 2C18, 2C19, and 2C58 were closely clustered with human, cynomolgus, and rhesus monkey P450 2Cs, whereas marmoset P450 2C76 was clustered with cynomolgus and rhesus monkey P450 2C76 (Fig. 2). All marmoset P450 2C mRNAs were abundant in livers (Fig. 4), where P450 2C proteins were also expressed (Supplemental Fig. 1). Marmoset P450 2C enzymes metabolized human P450 2C9 and 2C19 probe substrates, as well as non–P450 2C substrates (Table 2). Marmoset P450 2C19 catalyzed many drug oxidations, including S-warfarin 7-hydroxylation, diclofenac 4′-hydroxylation, tolbutamide methyl hydroxylation, flurbiprofen 4-hydroxylation, and omeprazole 5-hydroxylation (Tables 3 and 4), and substantially catalyzed bufuralol 1′-hydroxylation, which is also catalyzed by marmoset P450 2D6 and 2D7 (Uehara et al., 2015b). Similarly, human P450 2C19 catalyzes bufuralol 1′-hydroxylation with low efficiency compared with human P450 2D6 (Mankowski, 1999). Marmoset P450 2C19 might be partly responsible for bufuralol 1′-hydroxylation in livers. Marmoset P450 2C8, 2C18, 2C19, 2C58, and 2C76 also catalyzes 7-pentoxyresorufin O-deethylation. Similarly, cynomolgus monkey P450 2C9 and 2C19 metabolize many human P450 2C substrates, but also show some limited differences in drug oxidations of human P450 2C substrates (Hosaka et al., 2015b). Interestingly, marmoset P450 2C58 had the highest catalytic activities among five marmoset P450s tested toward non-P450 2C substrates, such as ethoxyresorufin, 7-ethoxycoumarin, coumarin, 7-pentoxyresorufin, and chlorzoxazone, whereas cynomolgus monkey P450 2C58 and human P450 2C58 were both pseudogenes. These findings might account for some limited species differences in drug-metabolizing enzymes between marmosets and cynomolgus monkeys or humans. Although monkey-specific P450 2C76 had a high impact on drug oxidations (Hosaka et al., 2015a), marmoset P450 2C76 showed few unique characteristics of human P450 2C substrates under the present conditions. These findings suggested that the substrate selectivity of marmoset P450 2C19 may reflect the functional similarity of the P450 2C enzyme between marmosets and humans.
Marmoset liver microsomes had efficiently catalyzed 7-hydroxylation for S-warfarin, but not for R-warfarin (Table 4), similar to those of humans but not those of cynomolgus monkeys. We reported previously that S-warfarin 7-hydroxylation in human liver microsomes was mainly catalyzed by P450 2C9, although cynomolgus monkey P450 2C19 preferentially catalyzed 7-hydroxylation of R-warfarin (Hosoi et al., 2012). Among all marmoset P450 2C enzymes, marmoset P450 2C19 efficiently catalyzed 7-hydroxylation for S-warfarin, but not R-warfarin, similar to human P450 2C9 (Table 4). Additionally, plasma concentrations of S-warfarin after intravenous administration of warfarin decreased faster than R-warfarin (Fig. 5), suggesting that S-warfarin was stereoselectively metabolized in marmoset livers, comparable to human livers. However, the metabolic clearances of S-warfarin showed large interindividual variations in comparison with those of R-warfarin, suggesting the possible genetic polymorphism of marmoset P450 2C19 involved in drug oxidations in marmosets. The similarity of stereoselectivity for warfarin 7-hydroxylation between marmosets and humans indicated that the drug-metabolizing properties of marmoset P450 2C19 were similar to humans under the present conditions.
Marmoset P450 2C19 showed 86% amino acid sequence identity with both human P450 2C9 and 2C19 (Table 1). Marmoset P450 2C19 efficiently metabolized the human P450 2C9 and 2C19 probe substrates, tolbutamide, flurbiprofen, S-warfarin, and omeprazole (Tables 3–4). Human P450 2C9 and 2C19 amino acid sequences showed 91% sequence identities; however, metabolic activities and/or substrate selectivity are generally different. Some critical key residues for substrate specificity have been identified by studies with chimeric constructs and amino acid substitutions between human P450 2C19 and 2C9. Human P450 2C19 His99, Pro220, and Thr221 reportedly have been key residues for omeprazole 5-hydroxylation (Ibeanu et al., 1996). Moreover, human P450 2C19 triple mutant (Glu241Lys/Asn286Ser/Ile289Asn) has reportedly shown high catalytic efficiency for warfarin metabolism for both S- and R-warfarin with a broadened regioselectivity (Jung et al., 1998). In the case of marmoset P450 2C19, Pro220, Lys241, and Asn289 might partly account for the impaired catalytic potential of P450 2C19 with both omeprazole 5-hydroxylation (human P450 2C19 selective activity) and S-warfarin 7-hydroxylation (human P450 2C9 selective activity). In this context, further study would be interesting.
Human P450 2C8, 2C9, and 2C19 proteins are expressed in human livers (Goldstein et al., 1994). In contrast, human P450 2C18 is expressed at the mRNA level, but its expression has not been detected at the protein level in any tissues (Klose et al., 1999). Regarding gene expression modulations, hepatic expression of human P450 2C genes is constitutively regulated by a number of liver transcription factors (Jover et al., 1998; Bort et al., 2004; Ferguson et al., 2005; Kamiyama et al., 2007; Wortham et al., 2007; Surapureddi et al., 2008). The human P450 2C8 gene has been post-transcriptionally regulated by microRNAs (Zhang et al., 2012). Investigating the transcriptional regulation of marmoset P450 2C19, which is important for drug metabolism as a functional counterpart of both human P450 2C9 and 2C19, is of great interest.
In terms of marmoset P450 2C expression level, all marmoset P450 2C mRNAs were predominantly expressed in livers (Fig. 4). Two marmoset P450 2C–like proteins have been detected in livers (Igarashi et al., 1997; Schulz et al., 2001) and induced by phenobarbital and rifampicin treatments (Schulz et al., 2001): tolbutamide methyl hydroxylation activities in liver microsomes have been greatly increased by phenobarbital (4.3-fold) and rifampicin (6.1-fold) (Schulz et al., 2001). Similarly, all three human P450 2C genes have been induced in the liver by phenobarbital, rifampicin, and dexamethasone (Raucy et al., 2002; Madan et al., 2003), which have been mediated through the xenobiotic-induced transcriptional activation by the nuclear receptors (Ferguson et al., 2002, 2005; Gerbal-Chaloin et al., 2002). In the present study, marmoset P450 2C8 and 2C19 were major tolbutamide methyl hydroxylases in marmoset livers. Detailed analysis is needed to elucidate that marmoset liver P450 2C8 and/or 2C19 may be induced by phenobarbital and rifampicin via these nuclear receptors. The similarity of tissue expression patterns possibly by common transcriptional regulation and induction properties in P450 2C genes might suggest that marmosets are potentially a suitable model for preclinical safety testing for humans.
Human P450 genes are highly polymorphic (http://www.cypalleles.ki.se/), and these genetic variants would cause interindividual differences in pharmacokinetics of drugs. Whole-genome sequencing has shown the genetic divergence of marmosets among multiple colonies (Marmoset Genome Sequencing and Analysis Consortium, 2014). Indeed, in our preliminary study, genetic variants of marmoset P450 2C19 were found with 20 marmoset genomes by direct sequencing (data not shown). Genetic polymorphisms of marmoset P450 2C genes might account for the interanimal variability in P450 2C–dependent drug metabolism.
In conclusion, four novel marmoset P450s, 2C18, 2C19, 2C58, and 2C76, were predominantly expressed in marmoset livers. Among five marmoset P450 2C enzymes, marmoset P450 2C19 efficiently metabolized human P450 2C9 and 2C19 probe substrates. Additionally, marmoset P450 2C19 preferentially metabolized S-warfarin both in vitro and in vivo, similar to the stereoselectivity of human liver microsomal P450 2C9. These results indicate that marmoset P450 2C enzymes had functional characteristics similar to those of humans, indicating that P450 2C–dependent metabolic properties are relatively similar between marmosets and humans.
Acknowledgments
The authors thank Drs. Norie Murayama and Masayuki Mogi for their technical help.
Authorship Contributions
Participated in research design: Uehara, Uno, Yamazaki.
Conducted experiments: Uehara, Uno, Kawano, Toda.
Contributed new reagents or analytic tools: Inoue, Sasaki.
Performed data analysis: Uehara, Shimizu, Uno, Utoh.
Wrote or contributed to the writing of the manuscript: Uehara, Uno, Yamazaki.
Footnotes
- Received June 25, 2015.
- Accepted July 29, 2015.
S.U. and Y.U. contributed equally to this work.
This work resulted from “Construction of System for Spread of Primate Model Animals” under the Strategic Research Program for Brain Sciences of the Japan Agency for Medical Research and Development. S.U. was also supported partly by Japan Society for the Promotion of Science Grant-in-Aid for Young Scientists (B) [Grant 15K18934].
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- BLAT
- BLAST-like alignment tool
- cj
- marmoset
- HPLC
- high-performance liquid chromatography
- P450
- cytochrome P450
- PCR
- polymerase chain reaction
- RT
- reverse transcription
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}