


Abstract
The anticancer prodrug ifosfamide (IFA) contains a chiral phosphorous atom and is administered clinically as a racemic mixture ofR and S enantiomers. Animal model studies and clinical data indicate enantioselective differences in cytochrome P-450 (CYP) metabolism, pharmacokinetics, and therapeutic efficacy between the two enantiomers; however, the metabolism of individual IFA enantiomers has not been fully characterized. The role of CYP enzymes in the stereoselective metabolism of R-IFA andS-IFA was investigated by monitoring the formation of both 4-hydroxy (activated) and N-dechloroethyl (DCl) (inactive, neurotoxic) metabolites. In the 4-hydroxylation reaction, cDNA-expressed CYPs 3A4 and 3A5 preferentially metabolizedR-IFA, whereas CYP2B6 was more active towardS-IFA. Enantioselective IFA 4-hydroxylation (R > S) was observed with six of eight human liver samples. In the N-dechloroethylation reaction, CYPs 3A4 and 2B6 both catalyzed a significantly higher intrinsic metabolic clearance (Vmax/Km) ofS-IFA compared with R-IFA. Striking P-450 form specificity in the formation of individual DCl metabolites was evident. CYPs 3A4 and 3A5 preferentially produced (R)N2-DCl-IFA and (R)N3-DCl-IFA (derived from R-IFA and S-IFA, respectively), whereas CYP2B6 correspondingly formed (S)N3-DCl-IFA and (S)N2-DCl-IFA. In human liver microsomes, the CYP3A-specific inhibitor troleandomycin suppressed (R)N2- and (R)N3-DCl-IFA formation by ≥80%, whereas (S)N2- and (S)N3-DCl-IFA formation were selectively inhibited (≥85%) by a CYP2B6-specific monoclonal antibody. The overall extent of IFAN-dechloroethylation varied with the CYP3A4 and CYP2B6 content of each liver, but was significantly lower forR-IFA (32 ± 13%) than for S-IFA (62 ± 17%, n = 8; p < .001) in all livers examined. R-IFA thus has more favorable liver metabolic properties than S-IFA with respect to less extensive N-dechloroethylation and more rapid 4-hydroxylation, indicating that R-IFA may have a distinct clinical advantage over racemic IFA.
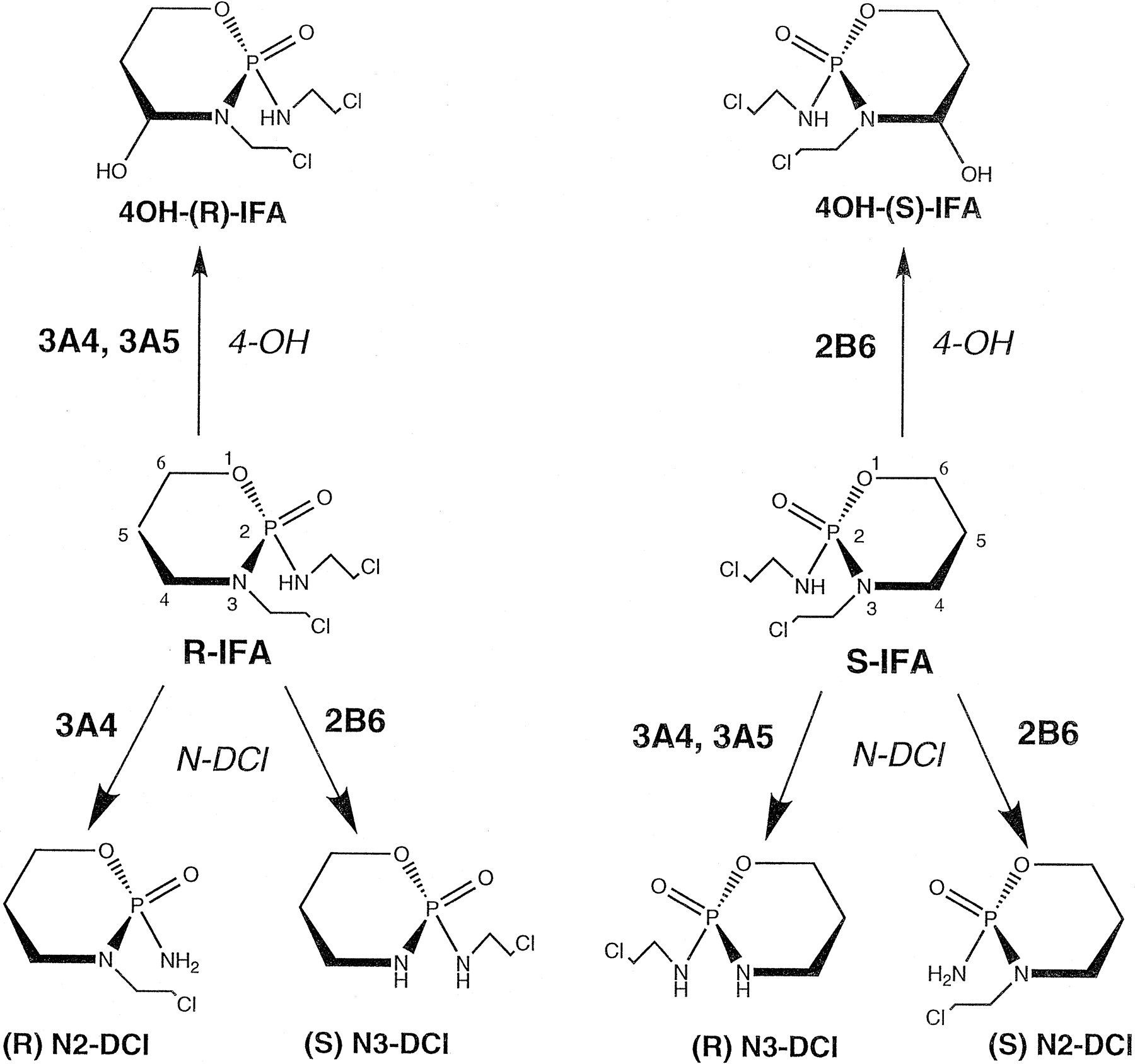
Ifosfamide (IFA)1 and its structural analog cyclophosphamide (CPA) belong to a group of oxazaphosphorine alkylating agents widely used in the therapeutic management of a variety of human malignancies (Fleming, 1997; Wright, 1997). IFA and CPA are both prodrugs that require activation by liver cytochrome P-450 (CYP) enzymes for therapeutic effect (Sladek, 1994). The primary activated metabolite of IFA, 4-OH-IFA, and its ring-opened tautomer isoaldophosphamide are readily transported across the cell membrane into the cytoplasm, where they can be metabolized to the inactive carboxyisophosphamide by aldehyde dehydrogenase. In tumor tissues, where aldehyde dehydrogenase activity is often lower than in host tissues, a correspondingly higher fraction of aldoisophosphamide escapes inactivation and can undergo general base-catalyzed, nonenzymatic elimination to yield acrolein and isophosphoramide mustard. The latter species is a DNA cross-linking derivative that is responsible for the cytotoxic activity of IFA (Wright, 1997). Alternative metabolic pathways include P-450-catalyzed dechloroethylation of IFA at N2 or N3 (Fig.1). These reactions yield equimolar amounts of the neurotoxic metabolite chloroacetaldehyde and the corresponding N-dechloroethylated (DCl) metabolite, 2-DCI-IFA or 3-DCl-IFA, both of which lack significant host toxicity and antitumor activity (Sladek, 1994).

Pathways of IFA metabolism: stereoselective 4-hydroxylation (4-OH) and N-dechloroethylation (N-DCl) of R-IFA and S-IFA catalyzed by CYPs 3A4, 3A5, and 2B6.
Shown at the bottom are the four DCl-IFA metabolites (R2 and S3, derived from R-IFA, andR3 and S2, derived fromS-IFA). The 4-hydroxy metabolites shown at top both exist as cis and trans isomers of the 4-hydroxy group that rapidly interconvert via the ring-opened aldoisophosphamide.
In spite of their similarities in chemical structure and mechanism of action, CPA and IFA differ significantly with respect to their pharmacokinetic and pharmacodynamic properties (Wright, 1997; Fleming, 1997). IFA exhibits greater cytotoxicity than CPA in some tumor models, produces less myelosuppression than CPA, and induces little cross-resistance (Hilgard et al., 1983; Berger et al., 1990). In early clinical trials, IFA was found to be severely urotoxic because of high concentrations of excreted acrolein, a problem that was substantially overcome with the introduction of sodium 2-mercaptoethane sulfonate, an effective uroprotective agent (Fleming, 1997). Chloroacetaldehyde is a major metabolite of IFA that contributes to urotoxicity and also the neurotoxicity associated with this drug (Goren et al., 1986; Pohl et al., 1989). In contrast to CPA, which is metabolized predominantly by ring 4-hydroxylation, with IFA as much as 50% of the administered dose is dechloroethylated by side chain oxidation (Kurowski and Wagner, 1993; Boddy et al., 1995a). The efficacy of IFA is thus determined by a delicate balance between two alternative, CYP-catalyzed metabolic pathways: a therapeutically beneficial drug activation/cytotoxic pathway (4-hydroxylation) and an undesirable drug deactivation/toxification pathway (N-dechloroethylation).
IFA is a chiral molecule that contains an asymmetric phosphorus atom and thus exists in two enantiomeric forms, R-IFA andS-IFA. The individual DCl metabolites, 2-DCl-IFA and 3-DCl-IFA, are also chiral and exist in both R andS forms (Fig. 1). Clinical preparations of IFA are racemic mixtures (50:50) of the two enantiomers (rac-IFA). Clinical and rodent model studies have reported significant differences in the stereoselective metabolism and disposition of the enantiomers, withS-IFA more extensively cleared by theN-dechloroethylation pathway than R-IFA (Wainer et al., 1994a,b; Granvil et al., 1996a). Differences in the enantiomeric composition of the 2- and 3-DCl-IFA metabolites present in urine of two rhabdomyosarcoma patients have been reported (Misiura et al., 1983), and significant interindividual differences in the stereoselectivity of urinary 3-DCl-IFA have been seen in a larger group of pelvic carcinoma patients (Granvil et al., 1996a; Wainer et al., 1996). Enantioselective excretion of IFA and its DCl metabolites was observed following a single 3-h infusion of drug in a pattern that suggests the involvement of at least two P-450 enzymes with distinct stereochemical selectivity toward IFA DCl (Wainer et al., 1996). Support for this proposal is provided by the differential responsiveness of the individual pathways of IFAN-dechloroethylation to the liver P-450-inducing agent phenobarbital in the rat model (Granvil et al., 1994; Lu et al., 1998). Rat CYP2B and CYP3A enzymes both contribute to liver microsomal IFAN-dechloroehylation, with CYP3A enzymes playing a more significant role in the case of CPA (Yu and Waxman, 1996; Brain et al., 1998; Yu et al., 1999), however, the catalytic roles of the corresponding human P-450 enzymes in the metabolism of the individual enantiomers of IFA have not been detailed.
Distinct subsets of liver P-450 enzymes contribute to the activation and inactivation of IFA (Chang et al., 1993; Weber and Waxman, 1993; Yu and Waxman, 1996; Roy et al., 1999), raising the possibility that these alternative pathways of P-450-catalyzed IFA metabolism can be modulated in a therapeutically beneficial manner using P-450 form-specific inhibitors and inducers (Brain et al., 1998). Another strategy proposed for increasing the therapeutic index of IFA is based on the administration of R-IFA in place of rac-IFA currently used in the clinic (Wainer et al., 1995), and derives support from the clinical and rodent model studies, summarized above, indicating thatR-IFA is less extensively N-dechloroethylated than S-IFA. The principal aims of the present study were to further investigate this proposal using human liver microsomal systems and cDNA-expressed human P-450 enzymes with the following two goals: 1) to establish the metabolic fates of R-IFA andS-IFA to better evaluate the proposed therapeutic utility of using one enantiomer compared with the other; and 2) to identify the principal human P-450 enzymes responsible for the enantioselective metabolism of R-IFA and S-IFA via both the 4-hydroxylation and the N-dechloroethylation pathways. Our findings strongly support the proposed use of R-IFA in place of rac-IFA, and further demonstrate that P-450s 3A4 and 2B6 both catalyze IFA N-dechloroethylation, but with unique stereochemical selectivities, enabling one to predict the contribution of each enzyme to overall human liver microsomal IFAN-dechloroethylation on the basis of the stereochemistry of the individual dechloroethylated metabolites.
Experimental Procedures
Materials.
Racemic IFA was obtained from the Drug Synthesis and Chemistry Branch, National Cancer Institute (Bethesda, MD). Purified R-IFA andS-IFA were obtained from Chiroscience, Ltd. (Cambridge, UK). DCl metabolites of IFA (N2-DCl-IFA and N3-DCl-IFA) and 4-hydroperoxy-IFA were gifts from Dr. J. Pohl (ASTA Pharma AG, Bielefeld, Germany). Troleandomycin (TAO) was obtained from Pfizer, Inc. (Brooklyn, NY). NADPH and trofosphamide were purchased from Sigma Chemical Co. (St. Louis. MO). All other specialty chemicals were obtained from Aldrich Chemical Co. (Milwaukee, WI). Anti-CYP2B6 monoclonal antibody (mAb-2B6) and Supersomes containing human P-450s expressed in a Baculovirus expression system containing NADPH P-450 reductase (OR) with or without cytochromeb5 (as indicated) were obtained from Gentest Corporation (Woburn, MA). Supersomes preparations used in these experiments were as follows: CYP2B6 + OR +b5, P255, lot 1; CYP3A4 + OR +b5, P202, lot 10; CYP3A5 + OR +b5, P235, lot E43; CYP3A5 + OR, P235, lot 6.
Microsomes.
Microsomes containing cDNA-expressed human P-450s were from Baculovirus-infected insect cells (Supersomes) engineered to express individual human P-450 cDNAs and human NADPH P-450 reductase. Some of the Supersomes also contained cytochromeb5, as indicated in the figures and tables. Supersomes generally exhibited specific P-450 catalytic activities severalfold higher than the average human liver microsomal sample when calculated on a per milligram protein basis. Negative controls corresponding to Supersome microsomes, which do not contain expressed P-450s, were run in parallel in each enzymatic incubation. A panel of 17 human liver microsomes, designated HLS2 to HLS36 (Roy et al., 1999), was prepared as described (Waxman et al., 1988) using livers obtained from organ donors. Inhibition studies used TAO for selective inhibition of CYP3A enzymes and mAb-2B6 (Yang et al., 1998) for selective inhibition of CYP2B6 at concentrations and under conditions shown to be maximally inhibitory (Roy et al., 1999). Microsomal protein concentrations were determined by the Bradford method with BSA as standard. P-450 form-diagnostic activities for 10 human liver microsomal P-450s were previously assayed in this same panel of livers as reported elsewhere (Roy et al., 1999).
IFA 4-Hydroxylation Assay.
Microsomal IFA 4-hydroxylase activity was assayed by monitoring formation of acrolein using an HPLC method with fluorescence detection (Bohnenstengel et al., 1997), with minor modifications (Huang and Waxman, 1999). Microsomal incubations were carried out under conditions where product formation was linear with respect to time and substrate consumption was <20%, as verified by analysis of residual substrate by gas chromatography (see below). Microsomal assays contained either 1 mM R-IFA or 1 mM S-IFA or, in some cases, 2 mM rac-IFA substrate, as indicated. These concentrations can be compared with peak plasma levels of 0.4 mM IFA (Boddy et al., 1995b) or 0.75 mM IFA (Lokiec et al., 1996) determined in clinical pharmacokinetic studies after bolus administration of IFA at 3 or 4 g/m2, respectively. Enzyme incubations included 1 mM NADPH and either 100 to 150 μg human liver microsomal protein or 20 to 40 pmol of cDNA-expressed P-450 protein in a total volume of 0.1 ml of 100 mM potassium phosphate buffer (pH 7.4) containing 0.1 mM EDTA. Complete incubation mixtures minus NADPH were preincubated for 4 min at 37°C, then initiated by addition of NADPH to 1 mM. Reactions were terminated after 20 min by the sequential addition of 40 μl of ice-cold 5% (w/v) zinc sulfate, 40 μl of ice-cold saturated barium hydroxide, and 20 μl of ice-cold 0.01 M hydrochloric acid. Following centrifugation, the analyte was derivatized with 3-aminophenol and hydroxylamine hydrochloride to yield 7-hydroxyquinoline, which was subsequently quantitated by reverse-phase HPLC on a Phenomenex C18 column (Phenomenex, Belmont, CA) coupled with a Waters 474 Scanning Fluorescence Detector set to at excitation and emission wavelengths of 350 and 515 nm, respectively. Standard curves were generated in parallel by incubating 4-hydroperoxy-IFA (0–20 μM) with BSA under the same assay conditions.
IFA N-Dechloroethylation Assay.
IFA N-dechloroethylation proceeds via an initial α-carbon hydroxylation that generates chloroacetaldehyde and dechloroethylated IFA (either 2-DCl-IFA or 3-DCl-IFA; see Fig. 1) in a molar ratio of 1:1 (Martino et al., 1992). Gas chromatography (described below) was used to separate and quantitate the individual stereoisomers of 2- and 3-DCl-IFA, whereas HPLC was used to monitor chloroacetaldehyde formation, which represents total IFA N-dechloroethylase activity. To monitor chloroacetaldehyde formation, IFA metabolism reactions were run for 20 min using the same incubation protocol and reaction termination steps described above for the 4-hydroxylation assay. This allowed for quantification of both acrolein and chloroacetaldehyde formation in a single enzyme assay. This improved the quantitation of IFA partitioning between the two alternate metabolic pathways (see Table 3). Chloroacetaldehyde was derivatized with adenosine to form the fluorescent adenosine derivative N6-ethenoadenosine (Nithipatikom et al., 1994) which was resolved by reverse-phase HPLC on a C18 column developed with 20% methanol at 1 ml/min and quantitated by fluorometry (excitation 270 nm, emission 411 nm) in an assay developed by Dr. Zeqi Huang of this laboratory (Huang and Waxman, 1999). Negative controls, consisting of Supersomes without expressed P-450 or BSA, were included with each series of incubations to determine background activity values, which were subtracted from each sample. Reactions to be analyzed for individual dechloroethylated metabolites of IFA were terminated following 45 min of incubation by addition of 1 ml of ice-cold methylene chloride with vortexing for 15 s, followed by centrifugation at 16,000g for 15 min at 4°C. The internal standard trophosphamide (10 μl of 1 mM trophosphamide stock dissolved in ethyl acetate) was added to the incubation tubes before transferring the extract to a test tube. The incubation tubes were extracted a second time with 1 ml of ice-cold methylene chloride and both extracts were then combined and dried under a gentle stream of nitrogen at room temperature. The dried residues were reconstituted in ethyl acetate and analyzed by gas chromatography.
Stereoselective metabolism of IFA: contributions of the 4-hydroxylation and N-dechloroethylation pathways
Gas Chromatography.
Individual DCl metabolites of R- and S-IFA were analyzed on a Hewlett-Packard model 6890 gas chromatograph (Hewlett-Packard, Palo Alto, CA) outfitted with a model 7673 autosampler and a nitrogen-phosphorus specific detector. An Alltech Chirasil-Val capillary column (Alltech Associates, Deerfield, IL), 0.53 mm i.d. × 20 mm long with a film thickness of 0.6 μm was used for the enantioselective separations (Boos et al., 1991; Granville et al., 1993). The carrier gas, helium, was metered through the column at a constant flow rate of 3.2 ml/min with additional helium makeup gas supplied to the detector to give a combined flow of 36 ml/min. The splitless mode injector and detector temperatures were 240 and 250°C, respectively. The oven temperature was held at 65°C for 3 min after injection, then increased to 180°C at 15°C/min, held until 30 min postinjection, increased to 198°C at 12°C/min, held until 41.5 min postinjection, increased to 210°C at 12°C/min, held until 46.5 min after injection, and finally, returned to 65°C. Retention times were as follows: S-IFA, 36.5 min; R-IFA, 37.7 min;S2-DCl, 25.5 min; R2-DCl, 26.5 min; S3-DCl, 33.1 min; R3-DCl, 34.9 min; and trofosfamide (internal standard), 41.5 min. Assays were carried out under conditions in which substrate depletion during the enzymatic reactions was ≤20%.
Validation of the gas chromatographic assay for detection and quantitation of each of the four DCl metabolites or IFA with respect to linearity of detection response, accuracy, intraassay and interassay precision, limit of detection, limit of quantitation, and metabolite recovery was carried out using standard methods. Metabolite detection was linear (r2 ≥ 0.993) over the range of 7.5 to 240 μM. Accuracy values ranged from 1.2 to 2.4% at 240 μM metabolite and from 1.9 to 3.7% at 7.5 μM metabolite. Mean interassay variation was <5%. Limit of detection was 0.2 to 0.4 μM and limit of quantitation (defined as the smallest sample concentration giving a relative S.D. of <5% and deviation from nominal concentration of <10%) was 6.9 to 7.2 μM. Metabolite recoveries under the assay conditions ranged from 83 to 88%.
Enzyme Kinetics and Data Analysis.
Initial reaction rates obtained at five to seven substrate concentrations (ranging from 0.25–5.0 mM IFA) were used to calculate apparent Km andVmax values using the software program Enzyme Kinetics (ChemSW Inc., Fairfield, CA). Data presented in Table 1(below) are based on Michaelis-Menten double reciprocal plots, which were linear (r > 0.98) for each of the major metabolites. Very similar kinetic values were obtained on regraphing the data using Eadie-Hofstee or Hanes plots or by direct linear plots of the data, as determined using the Enzyme Kinetics software package. Linear regression analysis of theN-dechloroethylation data obtained from a panel of human liver microsomes (n = 17 samples) was performed using the option “Analysis” in the software program GraphPad Prism (GraphPad Software Inc., San Diego, CA). The two-tailed Student’st test for paired sample means was employed to compare mean values between groups using the Data Analysis package of Microsoft Excel.
Kinetic analysis of CYPs 3A4 and 2B6 towards N-dechloroethylation of IFA enantiomers
Results
Stereoselective IFA Metabolism by cDNA-Expressed Human P-450s4-Hydroxylation.
CYPs 3A4, 3A5, and 2B6 together catalyzed ∼75% of human liver microsomal IFA 4-hydroxylation (Roy et al., 1999). The 4-hydroxylation of R-, S-, and rac-IFA was assayed using these three human CYPs expressed individually in a Baculovirus expression system (Supersomes) (Fig. 2). CYP3A4 exhibited ∼3-fold higher 4-hydroxylase activity with R-IFA compared with S-IFA. rac-IFA (2 mM) was less actively metabolized by CYP3A4 than would be predicted based on the sum of the activities observed with 1 mM R-IFA and 1 mM S-IFA alone (Fig. 2; rac-IFA). CYP3A5 exhibited an even more striking specificity for R-IFA, catalyzing very little 4-hydroxylation ofS-IFA. In contrast, CYP2B6 exhibited substantially greater 4-hydroxylation activity with S-IFA compared withR-IFA. CYP2C19, which makes only a minor contribution toward human liver microsomal IFA 4-hydroxylation (Roy et al., 1999), exhibited a stereochemical selectivity in favor of S-IFA 4-hydroxylation (S > R, ∼4:1), similar to CYP2B6 (data not shown).

Activation (4-hydroxylation) of R-IFA (1 mM), S-IFA (1 mM), and rac-IFA (2 mM) catalyzed by CYPs 2B6, 3A4, and 3A5.
Each of the cDNA-expressed CYPs (Supersomes) also contained cytochromeb5. Data are expressed as mean ± S.D. (n = 3) and are representative of the results of at least two independent experiments.
N-Dechloroethylation.
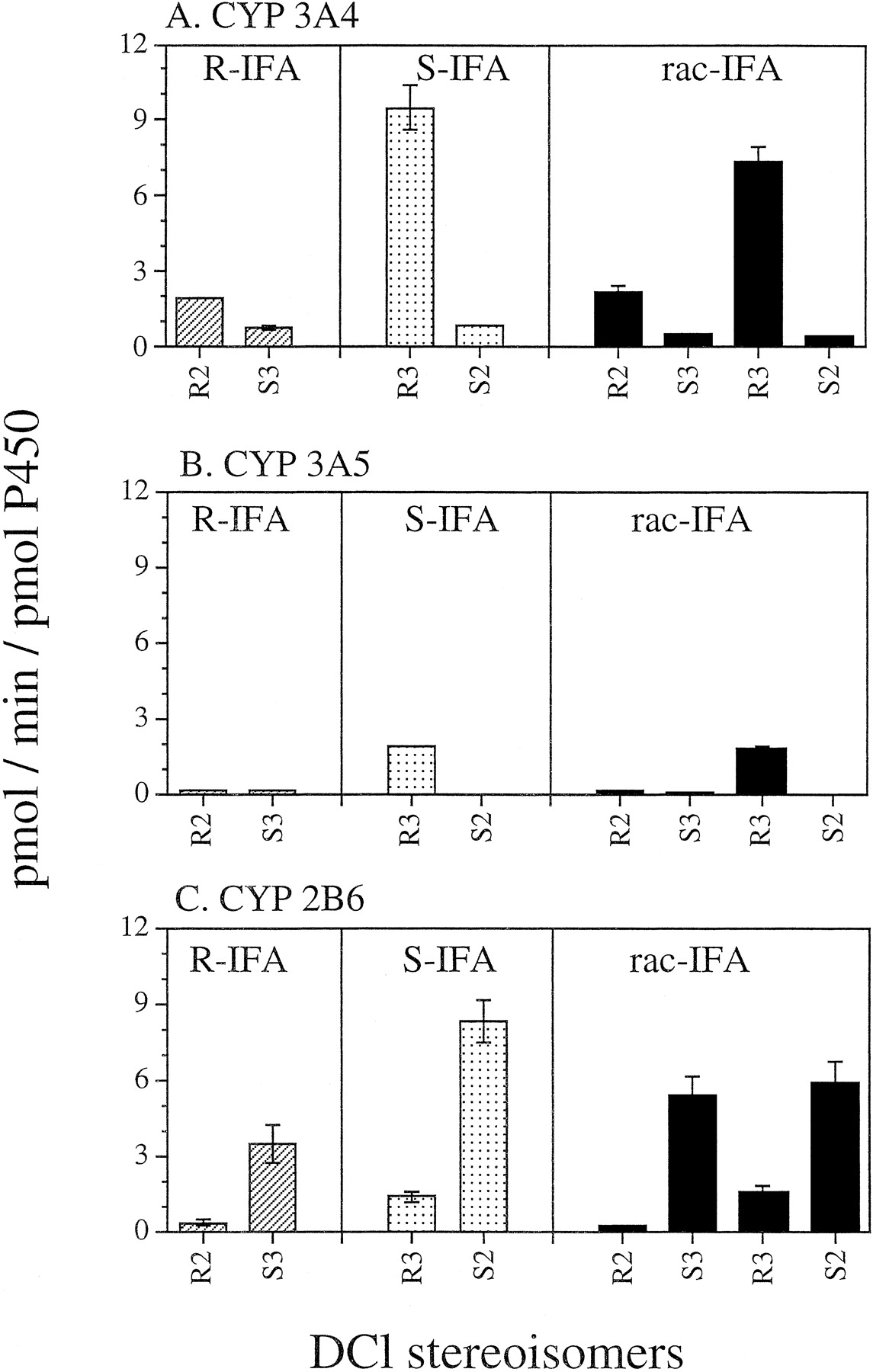
The stereospecificity of IFA N-dechloroethylation was investigated using cDNA-expressed CYPs 3A4, 3A5, and 2B6 (Fig.3). These three CYPs were examined because in aggregate they catalyze >90% of human liver microsomal IFAN-dechloroethylation (Huang et al., in press; Walker et al., 1994). N-dechloroethylation of R-IFA yields R2-DCl-IFA and S3-DCl-IFA, whereas S-IFA can generate R3-DCl-IFA and S2-DCl-IFA. rac-IFA can potentially be metabolized to a mixture of all four DCl-IFA metabolites, shown at the bottom of Fig. 1. These isomeric IFA metabolites were analyzed by enantioselective gas chromatography. As shown in Fig. 3A, S-IFA was N-dechloroethylated by CYP3A4 at a higher overall rate than R-IFA. Moreover, CYP3A4 primarily N-dechloroethylated S-IFA to yield R3-DCl-IFA, whereas S2-DCl-IFA was a minor metabolite (Fig. 3A). A very similar metabolic profile was obtained for CYP3A5, where R3-DCl-IFA (derived from S-IFA) was essentially the sole DCl metabolite (Fig. 3B). In contrast, CYP2B6N-dechloroethylated R-IFA and S-IFA at more similar rates, but exhibited a strong selectivity for formation ofS3-DCl-IFA and S2-DCl-IFA, derived fromR-IFA and S-IFA, respectively (Fig. 3C). Similar patterns were obtained when using rac-IFA as substrate (Fig. 3, right side).
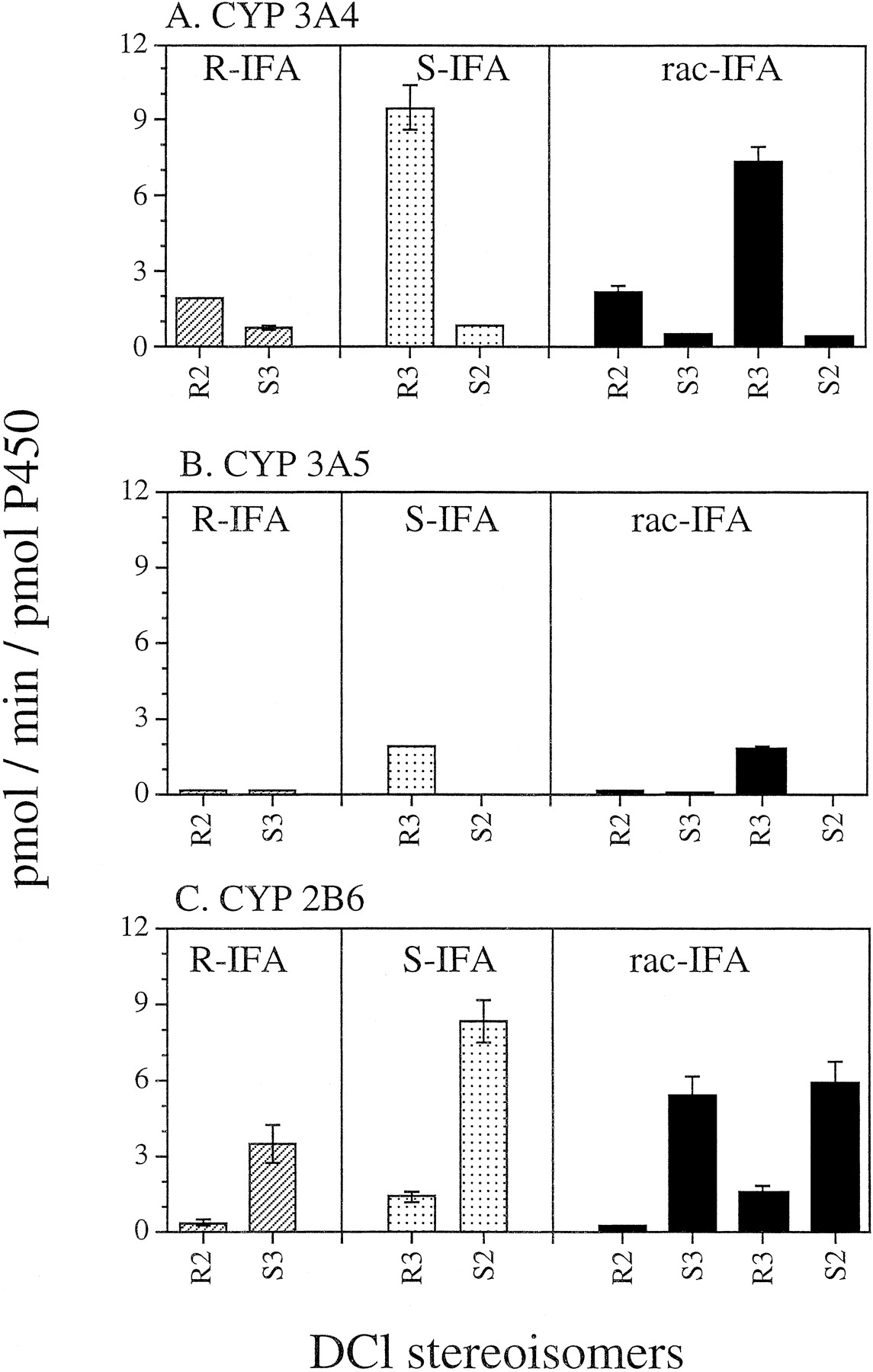
Enantioselective N-dechloroethylation of R-IFA (1 mM), S-IFA (1 mM), and rac-IFA (2 mM) catalyzed by CYPs 3A4 (A), 3A5 (B), and 2B6 (C).
R-IFA only produces R2-DCl-IFA andS3-DCl-IFA, whereas S-IFA yields exclusively R3-DCl-IFA and S2-DCl-IFA (see Fig. 1). Supersomes that express CYPs 3A4 and 2B6 also contained cytochrome b5. Data are expressed as mean ± S.D. (n = 3).
Apparent Km andVmax values for theseN-dechloroethylation reactions were determined separately for each enantiomer, following whichVmax/Km(intrinsic clearance) values were calculated from Michaelis-Menten double reciprocal plots (Table 1). Analysis of the data using either Eadie-Hofstee or Hanes plots gave very similar results, with no evidence for multiphasic kinetics (data not shown).Km values ranged from 0.5 to 2.3 mM in the various N-dechloroethylation reactions, and are similar to those measured for the corresponding 4-hydroxylation reactions catalyzed by these same two CYPs (Roy et al., 1999). These low millimolar Km values are typical of many P-450 drug substrates and are in the range of peak plasma IFA concentrations in patients treated with IFA as bolus injection at 4 g/m2 (Lokiec et al., 1996). For CYP3A4 and withR-IFA as substrate, the rank order of enzyme efficiency, expressed in the form of theVmax/Km ratio, was R2-DCl-IFA > S3-DCl-IFA; whereas withS-IFA as substrate, the order was: R3-DCl-IFA ≫S2-DCl-IFA (∼15-fold). By contrast, for CYP2B6 and withR-IFA as substrate, the rank order ofVmax/Km wasS3-DCl-IFA > R2-DCl-IFA; whereas with S-IFA as substrate, the order was S2-DCl-IFA ≫ R3-DCl-IFA (∼5-fold). With both enzymes, greater positional selectivity in theN-dechloroethylation reaction was seen withS-IFA, where CYP3A4 preferentially metabolized the ring chloroethyl group to give the N3 metabolite, whereas CYP2B6 metabolized the exocyclic chloroethyl group to give the N2 metabolite.
Stereoselectivity of IFA Metabolism by Human Liver Microsomes.
4-Hydroxylation
Analysis of the R-IFA and S-IFA 4-hydroxylase activities of a panel of eight human liver microsomes revealed significant interindividual differences both in the activity and the stereochemical specificity of the reactions (Fig.4). For two of the liver samples, HLS2 and HLS9, the sum of the 4-hydroxylase activities measured individually at 1 mM R-IFA and 1 mM S-IFA was similar to the activity obtained at 2 mM rac-IFA. By contrast, with the other liver samples, the 4-hydroxylase activity with rac-IFA was significantly lower than would be predicted based on the sum of the activities obtained with R-IFA and S-IFA individually, a finding that is characteristic of substrate saturation. Thus, when present as a racemic mixture, R-IFA and S-IFA do not behave as independent substrates, but have the potential for inhibitory interactions at the liver microsomal P-450’s active site in a manner similar to that seen with the cDNA-expressed enzymes (cf., Fig. 2).
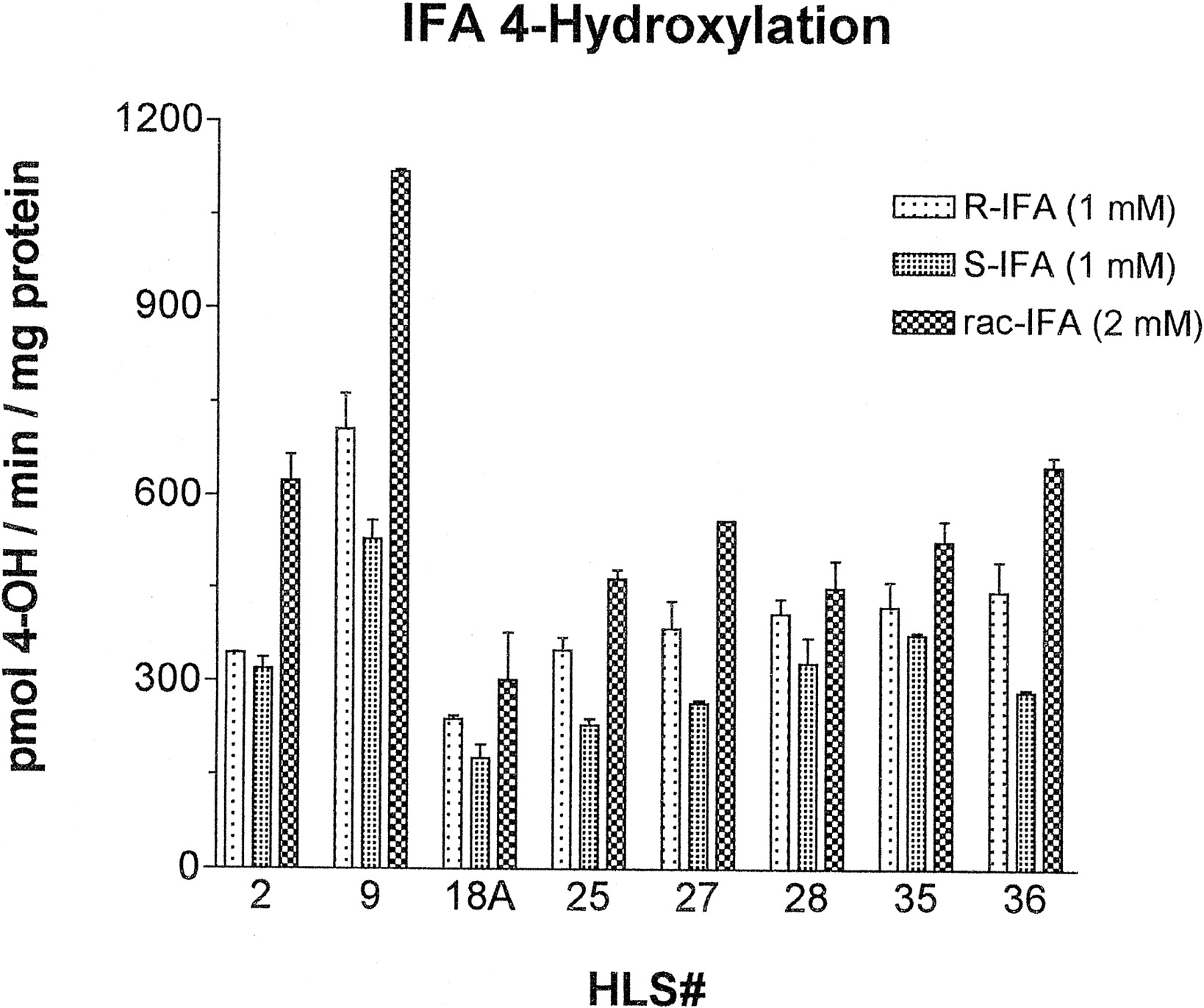
Activation (4-hydroxylation) of R-IFA (1 mM), S-IFA (1 mM), and rac-IFA (2 mM) catalyzed by eight individual human liver microsomes.
Data obtained for each liver sample are expressed as mean ± half the range of duplicate determinations.
N-Dechloroethylation.
A panel of 17 human liver microsomes was assayed forN-dechloroethylation of R-IFA andS-IFA. Data for eight of the individual human liver microsomes, which encompass the range of CYP2B6 and CYP3A4 marker activities seen in the full panel (Table2 and data not shown), are presented in Fig. 5. Large interindividual differences were noted in the formation of specific positional- and stereoisomers of DCl-IFA. R2-DCl-IFA was the exception, because its levels were relatively constant across the liver panel. The overall pattern was characterized by a high level of R3 metabolite, a medium level of S2 and S3 metabolites, and a low level of R2 metabolite (Fig. 5). This pattern mirrors the relative abundance of these four IFA metabolites in cancer patients (Granvil et al., 1996a). Among the livers, the CYP2B6-rich liver HLS2 demonstrated the highest rate of formation of S2-DCl-IFA andS3-DCl-IFA, whereas liver HLS9, which expresses a particularly high level of CYP3A4 activity (Table 2) displayed the highest rate of R3-DCl-IFA formation (Fig. 5). High amounts of R3-DCl-IFA were also formed by HLS36, despite its low level of CYP3A-dependent (Waxman et al., 1988) testosterone 6β-hydroxylase activity, indicating the possible participation of other P-450s in this reaction in some livers.
CYP2B6 and CYP3A diagnostic catalytic activities in human liver microsomes

Enantioselective N-dechloroethylation (N-deCl) of R-IFA (1 mM) and S-IFA (1 mM) catalyzed by eight human liver microsomes.
Data obtained for each liver sample are expressed as mean ± half the range of duplicate determinations.
Examination of the relationship between the rates of formation of each of the DCl metabolites of IFA and the CYP3A-dependent testosterone 6β-hydroxylase activity of each individual liver (n = 17) revealed that liver microsomal CYP3A activity correlated with the formation of R3-DCl-IFA (Fig.6A). A much weaker correlation between CYP3A activity and formation of R2-DCl-IFA, the minor metabolite of CYP3A4 (cf., Fig. 3 and Table 1) was observed (r = 0.39, p < .2; data not shown). In contrast, the rates of formation of S3-DCl-IFA (Fig. 6C) andS2-DCl-IFA (r = 0.81, p < .001; data not shown) correlated with the CYP2B6-dependent 7-ethoxy-4-trifluoromethyl coumarin (7-EFC) O-deethylase activity (Code et al., 1997) of the human liver microsome panel. There was no correlation between rates of formation of the two CYP3A-associated metabolites, R2 and R3 DCl-IFA, with the rates of formation of the two CYP2B6-characteristic metabolites, S2 and S3 DCl-IFA (data not shown). In contrast, R2 and R3 DCl-IFA correlated with each other (Fig. 6B), as did the rates of formation of S2 and S3 DCl-IFA (Fig. 6D). These latter findings mirror the correlations reported by Wainer and associates (Wainer et al., 1996) for the corresponding urinary metabolites in cancer patients, supporting the relevance of the in vitro liver microsomal enzyme system for prediction of metabolic profiles in patients. Of note, the single outlier point on the S2-DCl versus S3-DCl correlation curve (Fig. 6D; circled data point; liver HLS9), was not an outlier on the CYP2B6 activity versus S3-DCl curve (Fig.6C), indicating that this liver exhibits a S2-DCl activity that is lower than expected based on its CYP2B6 content. This may reflect the presence in liver HLS9 of a very high level of CYP3A4 (Table 2), which may compete with CYP2B6 for the rate-limiting microsomal NADPH P-450 reductase by actively metabolizingS-IFA to yield R3-DCl-IFA at the expense ofS2-DCl-IFA.
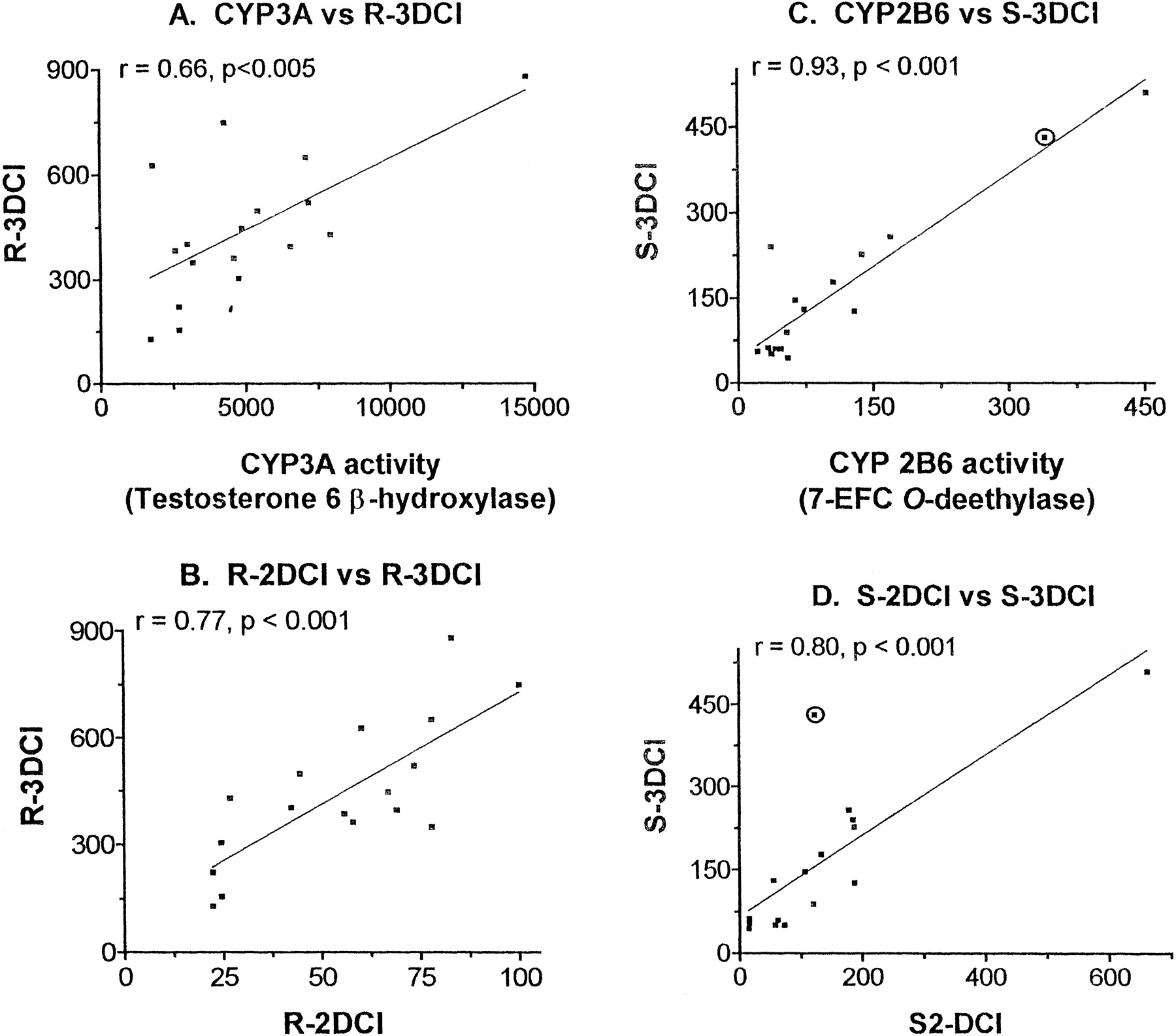
Correlation studies with CYP3A and CYP2B6 marker activities and the production of IFA’s DCl enantiomers in a panel of 17 human liver microsomes.
CYP3A4-specific testosterone 6β-hydroxylase activity (A) andR2-DCl-IFA formation (B) are correlated with the formation of R3-DCl-IFA, whereas the CYP2B6 component of human liver microsomal 7-EFC O-deethylase activity andS2-DCl-IFA formation (D), are correlated with the formation of S3-DCl-IFA (C). Data point circled in C and D corresponds to liver HLS9.
Inhibition of Stereospecific IFAN-Dechloroethylation in Human Liver Microsomes.
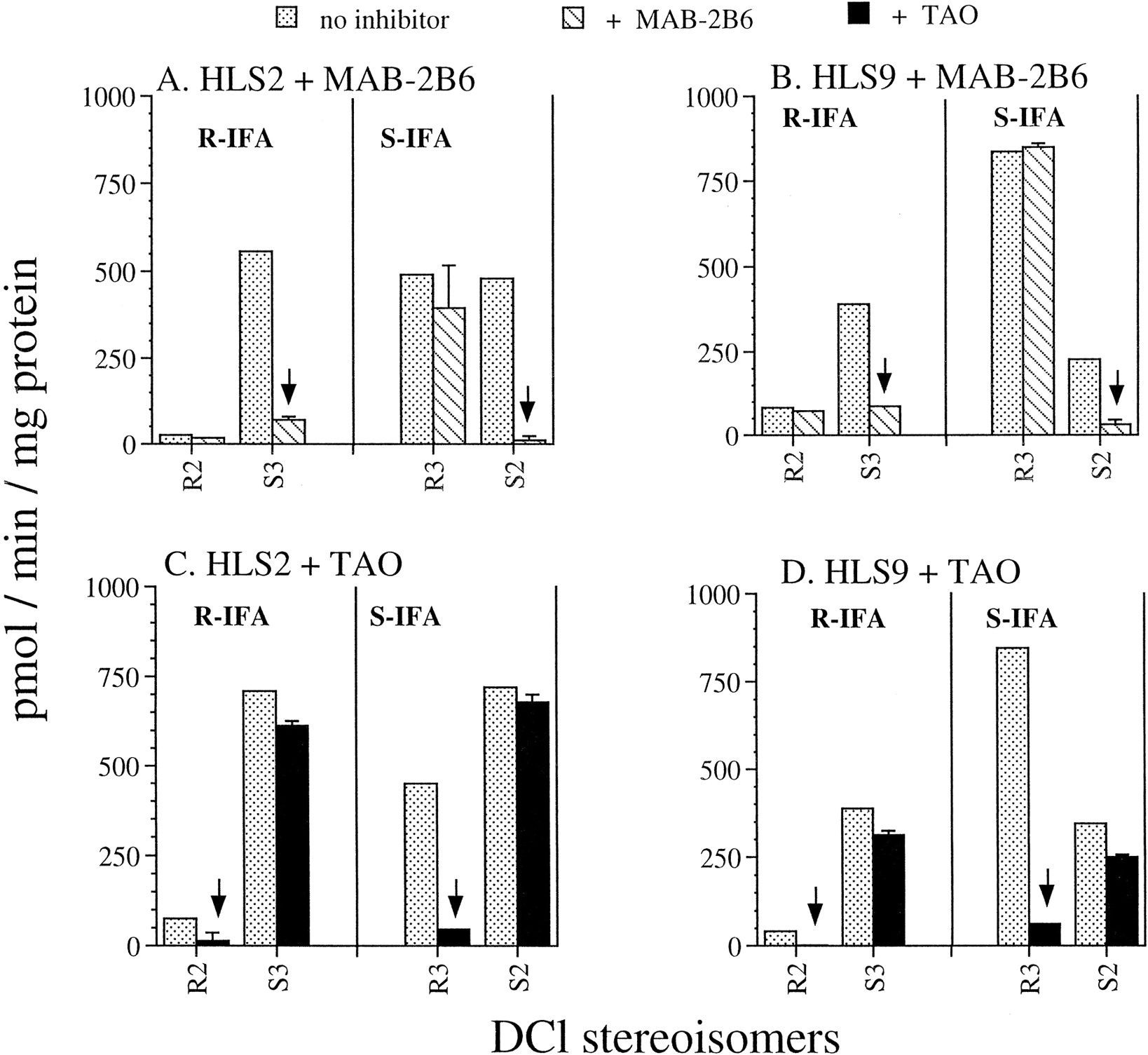
P-450 form-specific chemical inhibitors and inhibitory antibodies can be used to establish the contribution of individual P-450 enzymes to the metabolism of specific drugs in human liver microsomes. Monoclonal antibody immunoinhibitory to CYP2B6 (mAb-2B6) (Yang et al., 1998) and the CYP3A4-specific inhibitor TAO (Chang et al., 1994; Hickman et al., 1998) were employed to further establish the roles played by these two enzymes in the generation of DCl-IFA enantiomers in human liver microsomes. In liver HLS2, saturating levels of mAb-2B6 (50 μg/100 μg microsomal protein) inhibited by ∼85% the formation ofS3-DCl-IFA from R-IFA and completely blocked formation of S2-DCl-IFA from S-IFA (>95% inhibition; Fig. 7A, arrows). A similar inhibitory pattern was seen in a second liver sample, HLS9 (Fig. 7B). A high selectivity of the antibody was apparent, insofar as formation of the R3-DCl metabolite from S-IFA was not inhibited in either liver. In contrast, the CYP3A4 inhibitor TAO (50 μM) substantially blocked formation of R2-DCl-IFA andR3-DCl-IFA from R-IFA and S-IFA, respectively (≥80–90% inhibition) in both human liver microsomes (Fig. 7, C and D, arrows). Formation of the S3 andS2 DCl-IFA metabolites was inhibited by TAO to a much lesser extent (≤10–20%), consistent with the dominant role of CYP2B6 in these latter microsomal reactions indicated by the antibody inhibitions.

Inhibition of human liver microsomal IFA N-dechloroethylation of R-IFA and S-IFA by P-450 form-selective inhibitors.
Contribution by CYPs 3A and 2B6 to liver microsomalN-dechloroethylation was determined by applying inhibitory probes such as CYP3A-specific chemical inhibitor TAO (50 μM) and a CYP2B6-specific inhibitory mAb, mAb-2B6 (50 μg/100 μg microsomal protein) at concentrations shown to be maximally inhibitory. Data are shown for studies carried out in two individual human liver microsomes, HLS2 (A and C) and HLS9 (B and D). IFA enantiomers were assayed at a concentration of 1 mM. Data obtained for each liver sample are expressed as mean ± half the range of duplicate determinations. The arrows indicate the amount of 2- and 3-DCl-IFA enantiomers formed following inhibition by P-450 form-specific inhibitors.
Evaluation of Metabolic Properties of R-IFA versusS-IFA.
In view of the proposal, based on pharmacokinetic studies, that the metabolic properties of R-IFA may be superior to those ofS-IFA (Wainer et al., 1995), we compared R-IFA and S-IFA with respect to their total 4-hydroxylase andN-dechloroethylase activities (sum of all four DCl-IFA metabolites) catalyzed by CYPs 2B6, 3A4, and 3A5 and eight individual human liver microsomal samples (Table 3). With all three expressed enzymes and with five of the eight livers,S-IFA was more actively metabolized than R-IFA [Table 3; sum (4OH + DCl)]. With cDNA-expressed CYP2B6, bothR-IFA and S-IFA were metabolized predominantly byN-dechloroethylation, with little 4-hydroxylation (4OH/DCl ratio, ≪1). By contrast, with CYP3A4 and R-IFA as substrate, N-dechloroethylation corresponded to only 32% of total IFA metabolism (4-hydroxy-IFA + N-DCl-IFA) compared with 84% of the total metabolism with S-IFA as substrate (Table 3, last column). An even more striking difference was seen with CYP3A5 (17% of total metabolism via N-dechloroethylation for R-IFA versus 92% for S-IFA). Moreover, in the panel of liver microsomes, the proportion of substrate channeled through the N-dechloroethylation pathway was significantly lower with R-IFA (31.8 ± 12.6% metabolism viaN-dechloroethylation) than with S-IFA (61.8 ± 16.7% N-dechloroethylation) (mean ± S.D.,n = 8 livers; p < .001). This reducedN-dechloroethylation of R-IFA was achieved without a decrease in 4-hydroxylation activity compared withS-IFA as substrate (Table 3; total DCl-IFA versus 4-OH-IFA columns). This difference is also reflected in the 4-hydroxylation toN-dechloroethylation product ratio, which was ≥3- to 4-fold higher for R-IFA than S-IFA in almost all the liver samples (4-OH/DCl = 2.66 ± 1.49 versus 0.76 ± 0.56 for R and S IFA, respectively (mean ± S.D., n = 8 livers, p < .001).
Comparison of the rates of liver microsomalN-dechloroethylation of S-IFA versusR-IFA revealed a decrease in the extent to whichS-IFA is N-dechloroethylated in incubations of rac-IFA compared with incubations containing S-IFA alone (data not shown). This resulted in a consistently higherS/R N-dechloroethylation ratio (S/R DCl = 3.1 ± 1.3, mean ± S.D., n = 17) when the data was obtained from separate incubations of the two enantiomeric substrates (1 mM each) compared with incubations with rac-IFA (2 mM) (S/RDCl = 1.8 ± 0.6). Thus, R-IFA (1 mM), or a microsomal metabolite of R-IFA, can apparently inhibitS-IFA N-dechloroethylation.
Discussion
IFA metabolism is increasingly recognized as a key determinant of this drug’s antitumor activity, adverse side effects, and potential for emergence of chemoresistance. Large differences in the activity and level of expression of IFA metabolizing enzymes, specifically CYPs (Chang et al., 1993, 1997a; Walker et al., 1994), glutathione and glutathione S-transferase (Dirven et al., 1995), and aldehyde dehydrogenase (Bunting and Townsend, 1996), can in large part account for the significant interpatient differences in the clinical pharmacokinetics and biotransformation of IFA. In spite of the superior antitumor activity of IFA compared with its isomer CPA with some human cancers (Wheeler et al., 1986; de Kraker and Voute, 1989), the clinical efficacy and safety of IFA can be severely compromised due to extensiveN-dechloroethylation yielding chloroacetaldehyde, believed to be responsible for several adverse side effects observed with IFA chemotherapy, including cerebral neuropathy (Boddy et al., 1995a). IFAN-dechloroethylation is both regioselective and stereospecific and gives rise to the inactive metabolites 2-DCl-IFA and 3-DCl-IFA, respectively derived from dechloroethylation of the exocyclic (N2) and ring nitrogen atoms (N3), each of which exists as a pair of enantiomers (Fig. 1). Substantial stereospecificity in IFA efficacy, toxicity, disposition, and metabolism has been described (Farmer, 1988; Wainer et al., 1994a,b; Prasad et al., 1994; Granvil et al., 1996a). The present study identifies the human liver P-450 enzymes responsible for the stereospecific formation of IFA’s DCl metabolites and provides a metabolic basis for the wide variation in patient toxicities and responses to IFA. Moreover, the comparisons of the metabolic activity of R-IFA versus S-IFA presented here provide strong support for the proposed clinical use ofR-IFA in place of the racemic drug currently in use.
Experiments using cDNA-expressed enzymes revealed higher IFA 4-hydroxylase activity associated with CYP3A4 as compared with CYP2B6 (Fig. 2), in agreement with an earlier report (Chang et al., 1993) and a more recent comprehensive study (Roy et al., 1999) showing that CYP3A4 is a major IFA 4-hydroxylase in human liver microsomes. Moreover, R-IFA was found to be activated by CYP3A4 at a much higher rate than by CYP2B6 (Fig. 2). CYP3A5, which is expressed in human liver at a severalfold lower level than CYP3A4, also exhibited a strong preference for 4-hydroxylation of R-IFA compared withS-IFA. In contrast, CYP2B6 activated S-IFA at a much higher rate than R-IFA, although with the cDNA-expressed enzyme preparations used in this study S-IFA was still activated by CYP3A4 at a higher absolute rate than by CYP2B6. These findings indicate that the P-450 enzyme contributions to IFA 4-hydroxylation in human liver tissue may differ for R-IFA and S-IFA. Moreover, given the greater abundance of CYP3A4 compared with CYP2B6 in human liver (Shimada et al., 1994), the rate ofR-IFA activation is expected to exceed that ofS-IFA, a finding seen in six of the eight livers examined in the present study (Fig. 4). More extensive 4-hydroxylation ofR-IFA than S-IFA was also seen in a pharmacokinetic study using a rat mammary carcinoma model (Wainer et al., 1994b). The stereoselectivity for R-IFA compared withS-IFA in the 4-hydroxylation reaction in human liver microsomes was modest, however (Fig. 4), suggesting that CYP2B6 may be more active in human liver microsomes than would be predicted based on its abundance and cDNA-expressed activity. Alternatively, P-450 enzymes other than CYP2B6, such as CYP2C19 (as reported above), may contribute to human hepatic S-IFA 4-hydroxylation. Such a lack of stereoselectivity with respect to IFA 4-hydroxylation has been seen earlier in mice (Masurel et al., 1990) and in cancer patients (Farmer, 1988).
In the N-dechloroethylation reaction, cDNA-expressed CYP3A4 preferentially formed R2-DCl-IFA and R3-DCl-IFA from R-IFA and S-IFA, respectively. The much lower rate of formation of the R2 metabolite, both with expressed CYP3A4 (Fig. 3) and in human liver microsomes (Fig. 5) most likely reflects the fact that CYP3A4 actively metabolizesR-IFA by the alternative 4-hydroxylation pathway (Fig. 2). In contrast, cDNA-expressed CYP2B6 predominantlyN-dechloroethylated IFA to yield S3-DCl-IFA andS2-DCl-IFA, from R-IFA and S-IFA, respectively, in agreement with findings reported by Wainer and colleagues in abstract form (Granvil et al., 1996b). Thus, one P-450 enzyme (3A4) preferentially metabolizes the exocyclicN-chloroethyl group (N2) in the case of R-IFA, as contrasted with the ring N-chloroethyl group (N3) in the case of S-IFA, whereas a second P-450 enzyme (2B6) exhibits the opposite specificity: ring dechloroethylation of R-IFA and exocyclic dechloroethylation of S-IFA (cf., Fig. 1). The specificities seen for the cDNA-expressed CYPs also characterize these enzymes in human liver tissue, as is evident from correlation studies (Fig. 6) and from liver microsomal studies using CYP3A4- and CYP2B6-selective inhibitors (Fig. 7). These findings are consistent with an earlier report, where the urinary levels ofR2-DCl-IFA and R3-DCl-IFA were closely correlated in a group of 11 adult cancer patients treated with IFA, as were those of S2-DCl-IFA and S3-DCl-IFA (Wainer et al., 1996). The observation that the CYP2B6-derived DCl metabolites of IFA,S3-DCl-IFA, and S2-DCl-IFA, comprise a significant fraction of the total DCl metabolites of IFA, both in human liver microsomes (Fig. 5) and in cancer patients (Granvil et al., 1996a) indicates that CYP2B6 makes a significant contribution to IFAN-dechloroethylation (up to 50% in several livers). This finding is inconsistent with an earlier report suggesting that CYP3A enzymes catalyze >80% of this human liver microsomal activity (Walker et al., 1994).
Kinetic analysis of the stereoselectivity of IFAN-dechloroethylation by CYPs 3A4 and 2B6 (Table 1) revealed the highest Vmax andVmax/Km values for formation of R3-DCl-IFA by CYP3A4, followed byS2- and S3-DCl-IFA by CYP2B6. A similar pattern of DCl metabolites excreted into urine was observed in IFA-treated cancer patients (Wainer et al., 1996) and in human liver microsomal incubations (Fig. 5), where R3-DCl-IFA was the major metabolite, followed by the S2 and S3 metabolites, with R2-DCl-IFA generally being a minor product. Moreover, the total amount of DCl metabolites formed fromS-IFA in a panel of 17 human liver microsomes was found to be ∼3-fold greater than that from R-IFA. This observation is in agreement with the more extensive clearance byN-dechloroethylation that is seen for S-IFA in clinical studies (Boos et al., 1991; Wainer et al., 1996).
The present in vitro studies support the proposed use ofR-IFA in place of rac-IFA currently in clinical use, insofar as R-IFA undergoes less extensive deactivation/neurotoxification via N-dechloroethylation than the S-IFA enantiomer in all of the liver samples examined (Table 3), and in some cases exhibits a higher rate of 4-hydroxylation (Fig. 4). Moreover, because CYP3A4 is the most abundant P-450 associated with drug metabolism in human liver (Shimada et al., 1994) and can be induced severalfold by a number of drugs commonly prescribed together with IFA (Maurel, 1996), the use of R-IFA in place of rac-IFA may provide for an even greater therapeutic advantage. cDNA expression studies demonstrated that CYP2B6 exhibits proportionally higher R-IFA N-dechloroethylation compared with CYP3A4 (≥96% versus 32%) as well as minimal R-IFA 4-hydroxylase activity (Table 3). Hence, patients with a low liver CYP2B6 level would be expected to exhibit less extensiveR-IFA N-dechloroethylation (cf., livers HLS 25, 27, 28; Table 3). In contrast, patients with a high level of hepatic CYP2B6 (e.g., HLS2; Table 1) and/or a comparatively low level of hepatic CYP3A4 are likely to exhibit more extensive R-IFAN-dechloroethylation. This was clearly evident for the CYP2B6-enriched liver HLS2, which exhibited the highest extent ofR-IFA metabolized by the N-dechloroethylation pathway (53%) compared with the other livers (Table 3) and might not benefit from using R-IFA to the same extent as would a CYP2B6-deficient liver (e.g., HLS25). Nevertheless, the proportion ofR-IFA subject to N-dechloroethylation was found to be significantly lower than that of S-IFA in all livers examined, highlighting the benefit of using R-IFA. These findings further suggest that the therapeutic efficacy of IFA may be enhanced by administration of a CYP2B6-specific chemical inhibitor in combination with R-IFA with the goal of further suppressingN-dechloroethylation. Alternatively, individuals with a high CYP2B6/CYP3A4 ratio could benefit from using CPA in place of IFA, in view of the major role played by CYP2B6 in CPA activation and that of CYP3A4 in CPA N-dechloroethylation in human liver (Chang et al., 1993; Roy et al., 1999). It should be noted, however, that hepatic P-450 profiles are not fixed for a given patient, and can be induced or otherwise modulated by exposure to drugs routinely administered to cancer patients (Brain et al., 1998; Yu et al., 1999), including IFA itself (Chang et al., 1997b). Methods to phenotype individual patients for their levels of expression of individual P-450s using noninvasive methods are presently available or under development (Kivisto and Kroemer, 1997) and should find increasing use in efforts to improve drug responses by tailoring the choice of chemotherapeutic drugs to individual patients on the basis of their hepatic P-450 profiles.
After submission of this manuscript, Granvil and coworkers (Granvil et al., 1999) reported a major role for CYP3A4 in human liver microsomal metabolism of R-IFA and S-IFA to yieldR2-DCl-IFA and R3-DCl-IFA, respectively, and of CYP2B6 to yield S3-DCl-IFA and S2-DCl-IFA, respectively, in agreement with the present report. Those investigators also presented multiphasic Eadie-Hofstee plots suggestive of the presence of high-affinity (Km, ≤ 10 μM), low-capacity IFA N-dechloroethylase enzyme(s) in human liver microsomes; however, the relationship of that low-capacity microsomal activity to CYP2B6 and CYP3A4, characterized in the present study as high Km, high-capacity IFAN-dechloroethylases (Table 1), was not established in that report.
In conclusion, substantial interindividual differences in the human hepatic levels and expression of the IFA-metabolizing P-450 enzymes CYP3A4 and CYP2B6 are likely to make important contributions to the interpatient variability in the enantioselective metabolism and clinical response to IFA. Metabolic studies carried out with cDNA-expressed human P-450 enzymes and human liver microsomes strongly support the proposal that R-IFA can be used to reduceN-dechloroethylation and the associated formation of the neurotoxic metabolite chloroacetaldehyde commonly seen with rac-IFA. IFA induces several human hepatic P-450 enzymes active in IFA metabolism (Chang et al., 1997b) via a process that is likely to contribute to the autoinduction pharmacokinetics that characterizes the use of IFA in the clinic (Kurowski and Wagner, 1993; Prasad et al., 1994). It will thus be of interest to investigate whetherR-IFA and S-IFA exhibit differential capacities to induce human hepatic P-450s capable of metabolizing cancer chemotherapeutic drugs, such as CYP3A4, CYP2B6, and CYP2C subfamily enzymes. Finally, hepatic P-450 profiling in cancer patients may be useful for tailoring the choice of chemotherapeutic drugs before the initiation of treatment in an effort to better design drug regimens on an individualized basis.
Footnotes
-
Send reprint requests to: Dr. David J. Waxman, Department of Biology, Boston University, 5 Cummington St., Boston, MA. E-mail: djw{at}bio.bu.edu
-
This work was supported in part by National Institutes of Health Grant CA49248 (to D.J.W.).
- Abbreviations used are::
- CYP
- cytochrome P-450
- IFA
- ifosfamide
- rac-IFA
- racemic IFA
- CPA
- cyclophosphamide
- DCl
- dechloroethyl
- R2-DCl-IFA and S2-DCl-IFA
- N2-dechloroethylated metabolites of R-IFA andS-IFA, respectively
- S3-DCl-IFA and R3-DCl-IFA
- N3-dechloroethylated metabolites ofR-IFA and S-IFA, respectively
- HLS
- human liver microsomal sample
- OR
- NADPH P-450 oxidoreductase
- mAb-2B6
- monoclonal antibody to CYP2B6
- TAO
- troleandomycin
- 7-EFC
- 7-ethoxy-4-trifluoromethyl coumarin
- Received May 4, 1999.
- Accepted August 2, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}