


Abstract
The residence times of nicotine and its metabolites in rat brain after acute peripheral nicotine administration were determined. We hypothesize that nicotine metabolites will reach pharmacologically significant concentrations in brain. Cotinine, nornicotine, and norcotinine were structurally identified by dual label radiochemical and gas chromatography-mass spectrometric analysis as biotransformation products of nicotine present in rat brain after s.c. injection ofS(−)-nicotine. Two unidentified minor metabolites were also detected in brain. The half-lives in brain of nicotine metabolites were determined after a single s.c. injection of [2′-14C]-(±)nicotine (0.8 mg/kg) and analysis of radiolabeled metabolites by high pressure-liquid radiochromatography. The brain half-lives of nicotine, cotinine, and nornicotine were 52, 333, and 166 min, respectively. Peak brain concentrations of nicotine metabolites were 300, 70, and 7 nM for cotinine, nornicotine, and norcotinine, respectively. Even with potential accumulation of cotinine in brain after chronic nicotine administration, it is likely that the brain concentration of cotinine will be insufficient to produce neuropharmacological effects resulting from activation of nicotinic receptors to induce dopamine release. Conversely, the concentration of nornicotine in brain after acute nicotine approaches the range found to be neuropharmacologically active. It is likely that nornicotine will accumulate in brain on chronic nicotine administration based on the brain half-life of this metabolite. Importantly, nornicotine is also a major alkaloidal component of tobacco. Thus, as a consequence of tobacco use, alkaloidal and metabolically formed nornicotine may reach concentrations in brain sufficient to produce pharmacological effects.
Although peripheral metabolism of nicotine has been studied extensively (Gorrod and Jenner, 1975;Kyerematen and Vesell, 1991; Crooks, 1993; Gorrod, 1993), nicotine metabolism in brain has been given little attention until recently. The presence of nicotine metabolites in brain is an important factor to consider because nicotine biotransformation products have been shown to be pharmacologically active (Crooks and Dwoskin, 1997), and their presence in brain constitutes a potential contribution to the neuropharmacological effects resulting from nicotine exposure. In earlier studies, only one major metabolite, cotinine, was reported in mouse and cat brain after i.v. injection of [14C-N-1′-methyl]nicotine (Appelgren et al., 1962). More recently, similar results were found in the rat (Deutsch et al., 1992). Other studies have reported the presence of cotinine and four other unidentified minor metabolites in the brain of [14C-N-1′-methyl]nicotine-treated animals (Schmiterlow et al., 1967). The distribution and pharmacokinetics of radiolabeled nicotine in brain after various routes of peripheral administration in several animal species were reported (Saji et al., 1984; Nordberg et al., 1989; Plowchalk et al., 1992;Yamada et al., 1992). Generally, these studies did not determine the concentrations of nicotine biotransformation products. In other studies, both [14C]cotinine and [14C-N-1′-methyl]nicotine-N-oxide have been detected by high pressure liquid radiochromatography (HPLRC)1 in mouse brain after peripheral administration of [14C-N-1′-methyl]nicotine (Stalhandske, 1970; Petersen et al., 1984). The use of [14C]-methyl-labeled nicotine in these studies precludes the detection of nicotine metabolites that have undergone oxidative N-demethylation, e.g., metabolites such as nornicotine and norcotinine.
Previous HPLRC studies from our laboratory have shown that when rats are injected s.c. with [3H-N-methyl]nicotine, a radiolabeled metabolite coeluting with authentic cotinine standard was observed; this had a long residence time in the central nervous system (CNS) (Crooks et al., 1995). Also, a significant amount of tritium eluted in the void volume of the brain supernatants, suggesting either dissociation of the tritium label, which is unlikely, or metabolism of the tritiated methyl group through an oxidative process leading to loss of tritium from the molecule. Thus, the tritium in the void volume was attributed to a low molecular weight oxidized 1-carbon unit, such as tritiated formaldehyde or tritiated formic acid. This led us to suspect that other nicotine metabolites might be present in brain that would be devoid of the 3H label and, therefore, not detectable by HPLRC. Such metabolites would benor-metabolites of which two are known to be formed peripherally from nicotine, viz. nornicotine and norcotinine (Bowman and McKennis, 1962; Zhang et al., 1990; Curvall and Kazemi Valla, 1993;Gabrielsson and Gumbleton, 1993; Liu et al., 1993).
The presence of nornicotine and cotinine in brain after nicotine administration may be of additional relevance to tobacco smoking, because these compounds are also alkaloidal constituents of tobacco. Thus, these compounds in smokers' brain may arise from both nicotine metabolism and from self-administration of these alkaloids. The presence of nornicotine and cotinine in brain may also have significance in understanding the neuropharmacological effects of tobacco use, because both metabolites have been shown to be neuropharmacologically active (Risner et al., 1985; Goldberg et al., 1989; Dwoskin et al., 1995, 1999a,b; Stolerman et al., 1995; Teng et al., 1997). Therefore, we carried out similar metabolic experiments using nicotine with a 14C label at the 2′-carbon to determine whether nor-metabolites of nicotine were indeed formed in the brain after peripheral nicotine administration to rats (Crooks et al., 1997). These HPLRC experiments clearly showed that at 4 h postinjection of peripheral nicotine, two metabolites were detected and quantified in brain by coelution with authentic standards of nornicotine and norcotinine. These data represent the first comprehensive study on the determination and quantitation of nicotine metabolites in brain after acute nicotine administration. However, these brain metabolites have been identified only tentatively by HPLRC. This tentative identification is based on their coelution with authentic UV absorbing metabolic standards, and on the observation that when [3H-N-methyl]nicotine is used instead of [2′-14C]nicotine, none of thenor-metabolites are detected, although cotinine is clearly observed. We now report the definitive structural identity of cotinine, nornicotine, and norcotinine in rat brain after peripheral nicotine administration using dual-label tracers and gas chromatography-mass spectrometry (GC-MS) analysis. Additionally, we have determined the residence time and half-life in rat brain of each of these metabolites after a single peripheral dose of [2′-14C]nicotine.
Materials and Methods
Compounds.
S(−)-[3H-N-Methyl]nicotine (specific activity: 81 Ci/mmol) and (±)[2′-14C]nicotine (specific activity: 55 mCi/mmol) were obtained from New England Nuclear (Boston, MA) and Moravek Biochemicals (Brea, CA). The radiochemical purities of each of the radiolabeled compounds were >98%, as determined by HPLRC (Cundy and Crooks, 1983). Altima AP scintillation cocktail (Packard BioSciences, Groningen, the Netherlands), S(−)-nicotine tartarate (Research Biochemicals International, Natick, MA), andS(−)-cotinine (Sigma, St. Louis, MO) were purchased. HPLC grade triethylamine, acetonitrile, phosphoric acid, and sodium phosphate (dibasic, anhydrous) were purchased from Fisher Scientific (Pittsburgh, PA). S(−)-Nornicotine andS(−)-norcotinine were prepared in our laboratory as described previously (McKennis et al., 1959; Jacob, 1982).
In Vivo Metabolic Experiments.
Male Sprague-Dawley rats (250–350 g) were obtained from Harlan Laboratories (Indianapolis, IN) and were housed two per cage with free access to food and water in the Division of Laboratory Animal Resources at the College of Pharmacy, University of Kentucky. Experimental protocols involving animals were approved by the Institutional Animal Care and Use Committee at the University of Kentucky.
[2′-14C]Nicotine Experiments.
Experiments were performed using groups of four rats (n= 4 per group for each time point analyzed). Rats were injected s.c. with 0.8 mg/kg nicotine free-base equivalent containing 50 μCi of [2′-14C]nicotine in saline (0.2 ml/100 g). This dose of nicotine is nontoxic and is a commonly used dose in behavioral studies using rat as the model, and provides the greatest probability of allowing quantification of nicotine metabolites in brain. In radiolabeled experiments, rats were sacrificed by rapid decapitation at 5, 30, 60, 240, 360, 480, 720, 960, and 1440 min, and the trunk blood and whole brain, including brain stem, were obtained within 1 to 2 min. The presence of radiolabeled nicotine and its metabolites in brain was determined by homogenizing the brain in 3 volumes of ice-cold 1.15% w/v KCl using a polytron homogenizer. The homogenate was then centrifuged at 3000g for 30 min. The supernatant was separated and treated with 2% w/v ZnSO4 and the mixture was maintained in a water bath for 1 h at 34°C. The precipitated protein was then pelleted by centrifugation at 30,000g for 60 min. The resulting supernatants were analyzed directly by HPLRC.
Dual-Labeled Nicotine Experiments.
Experiments were performed using a group of four rats, which were injected with 0.8 mg/kg nicotine free-base equivalent containing 25 μCi of [2′-14C]nicotine and 25 μCi of [3H-N-methyl]nicotine in saline (0.2 ml/100 g). Rats were sacrificed at 240 min (a time point at which significant amounts of all major metabolites are observed) and trunk blood and brain were obtained. Tissue was treated as described above, and supernatants were analyzed directly by HPLRC.
Unlabeled Nicotine Experiments.
A group of five rats were each injected s.c. with 0.8 mg/kg nicotine free-base equivalent, and sacrificed by rapid decapitation 240 min postinjection. Brain supernatants were obtained as described previously, and pooled for analysis by GC-MS. The supernatants were acidified with aqueous HCl to pH 3 to 4 and lyophilized. To the lyophilized powder was added 0.5 ml of a 5 N aqueous NaOH solution, and the mixture was extracted with two-10 ml volumes of methylene chloride. The methylene chloride layers were combined and evaporated to dryness over a stream of nitrogen gas. The resulting residue was reconstituted in 5 μl of methanol, and this solution was injected onto the GC-MS analytical unit. Chromatography was performed on a Hewlett Packard HP 6890 gas chromatograph (Avondale, PA) equipped with an HP 7683 series automatic sampler with a HP 7683 series injector, and interfaced to a HP 5973 mass selective detector operating in scan mode. The GC was equipped with a fused silica capillary column (30 m × 0.25 mm i.d.) containing DB-5 as stationary phase (1.0-μm film thickness) obtained from J and W Scientific (Falson, CA). The total 5-μl brain supernatant extract was injected in the pulsed splitless mode with the injection pulse (20.0 psi) switched on for 0.6 min and the purge valve closed for 0.5 min. The temperatures of the injection port, interface, and electron impact source were 250, 280, and 230°C, respectively. The initial oven temperature was 110°C. After holding for 1 min, the temperature was increased at a rate of 1°C/min to 190°C, and then increased at a rate of 4°C/min to a final temperature of 270°C. The total separation time was 120 min. Helium was used as a carrier gas at a flow rate of 1 ml/min in the constant flow mode. Under these conditions, nicotine, nornicotine, cotinine, and norcotinine eluted with retention times of 26.6, 32.8, 61.8, and 69.2 min, respectively. The electron multiplier voltage was set to 1700 mV. The nature of the peaks were identified and confirmed both by retention time of the compounds and by ion extraction chromatograms atm/z: 162, 133, and 119 for nicotine; 148, 147, and 119 for nornicotine; 176, 145, and 98 for cotinine; and 162, 134, and 80 for norcotinine.
HPLRC System.
The HPLRC system comprised a Packard Series 1100β Radiomatic flow-through detector (Packard Instruments, Meriden, CT) connected to a Spectroflow 783 UV detector (ABI Analytical, Ramsey, NJ) operating at a fixed wavelength of 254 nm. A Beckman System Gold PSM 116 pump (Fullerton, CA) was used. Column effluent (mobile phase) was mixed in a 1:3 ratio volume with Altima flow AP scintillation cocktail before entering the radiomatic detector. Output of the two detectors was recorded simultaneously on the same chromatogram on different channels on an Epson LX 810 printer, and were corrected for lag time. Chromatographic analyses were performed on a 25 × 0.46 cm Partisil 10 C8 column (Clifton, NJ) connected to a Phenomenex 3.0 × 0.46 cm C8 guard column (Torrance, CA). The mobile phase was 0.1 M sodium phosphate/acetonitrile (95:5 v/v%), and 30 μl/liter triethylamine, and the pH was adjusted to 7.0 with phosphoric acid. The system was run at a flow rate of 1.5 ml/min. Each supernatant sample was coinjected with 100 μl of a standard solution containing 1.5 μg of authentic standards of S(−)-nicotine, S(−)-cotinine, andS(−)-norcotinine. Radioactive metabolites eluting from the HPLC column were identified by comparing their retention times with those of the UV-active authentic standards. The retention times for the analytes were 5, 13, 26, 28, 38, and 55 min for metabolite A, nornicotine, norcotinine, metabolite B, nicotine, and cotinine, respectively. For dual label analysis, the energy range settings for3H detection were set at 0 keV for the lower discriminator and 13 keV for the upper discriminator. For14C detection, energy range settings were 14 keV for the lower discriminator and 114 keV for the upper discriminator. Energy spillover of 14C into the3H spectrum was 70% and the quenching efficiencies for 3H and 14C were 32.5 and 48.2%, respectively. The third channel of the analytical unit was used for UV detection at 254 nm.
Results
Dual-labeled nicotine metabolism experiments were performed to confirm the presence of nor-metabolites of nicotine in rat brain. One label was 3H, which was placed on theN-methyl group of nicotine; the second label was14C, which was placed in the pyrrolidine ring skeleton at C-2′. The presence of nor-metabolites can be established using this dual-labeled form of nicotine by dual-channel ratio radioactivity analysis. Nor-metabolites should only retain the 14C label, i.e., the3H label is lost through demethylation, whereasN-methyl metabolites should retain both labels. Thus, rats were injected peripherally with14C-/3H-labeled nicotine, and 4 h later, brains were obtained and supernatants analyzed by HPLRC dual channel analysis. Figure 1clearly shows that the radiolabeled bands tentatively identified as nornicotine and norcotinine have both lost their3H label, demonstrating that they arenor-metabolites. The 14C radiochromatograms afforded radiolabeled bands that were much broader than the bands observed in the 3H channel analysis. This is because the 14C-labeled nicotine is racemic, whereas the [3H]nicotine is the pure S(−)-enantiomer, and the added mass of unlabeled nicotine is in the form of the S(−)-enantiomer. Thus, a typical chirodiastaltic effect is observed with the racemic 14C-labeled nicotine, causing significant peak broadening of both [14C]nicotine and its metabolites (Cundy and Crooks, 1983).

HPLRC Partisil-10 C8chromatogram of supernatant from 4-h brain homogenate from a representative rat administered a single s.c. dose of 0.8 mg/kg nicotine containing 25 μCi each of [2′-14C]nicotine and [3H-methyl]nicotine.
Top: cpm above background for the 14C channel; bottom: cpm above background for the 3H channel. Radiolabeled bands were identified by comparing retention times with those of coinjected UV-absorbing authentic metabolic standards. HPLRC operating conditions are as described in Materials and Methods.
To prove conclusively that the metabolites present in brain after acute peripheral nicotine administration are cotinine, nornicotine, and norcotinine, GC-MS analysis was performed on rat brain supernatant extracts. Because of the extremely small amounts of nicotine metabolites present in brain after acute administration, it was not possible to quantify these metabolites in single animal experiments. Thus, brain supernatants from five animals that had each been administered 0.8 mg/kg of nonradiolabeled S(−)-nicotine 4 h earlier were pooled and processed, and the extract was injected onto the GC-MS. Figure 2illustrates the GC chromatograph of this extract. Nicotine and the three metabolites, nornicotine, cotinine, and norcotinine, were easily detected by GC analysis, displaying retention times of 26.6, 32.8, 61.8, and 69.2 min, respectively. Cotinine was by far the most abundant metabolite in brain. Nornicotine was the second most abundant, whereas norcotinine was only a minor metabolite in brain. The unambiguous structural assignment of each of the above nicotine metabolites was determined by ion extraction GC-MS. The retention times and fragmentation patterns obtained for each metabolite were consistent with their proposed structures. The fragmentation patterns obtained for each of the metabolites and the selected ion chromatograms are shown in Fig. 3. Each metabolite spectrum was peak matched with the spectrum of the authentic compound in the Tobacco and Health Research Institute and Wiley library databases. These data confirm unequivocally the presence of these metabolites in rat brain 4 h after acute peripheral nicotine exposure. The results are entirely consistent with those obtained from radiochemical studies performed at the 4-h time point (Crooks et al., 1997).

GC chromatograph of a sample of an extract of brain supernatant pooled from five rats 4 h after s.c. administration of 0.8 mg/kg of nonradiolabeled S(−)-nicotine (NIC).
Data are expressed as relative abundance in arbitrary units as a function of time (min). Separations were carried out on a fused silica capillary column containing DB-5 stationary phase (see Materials and Methods for operating conditions). Brain metabolites of nicotine were identified and structures confirmed by comparing the retention times of the metabolites with those of authentic metabolic standards and by carrying out the analysis by ion extraction mass spectrometry (see Fig. 3 for details). NNIC, nornicotine; COT, cotinine; NCOT, norcotinine.

Structural identification of nicotine (A), cotinine (B), nornicotine (C), and norcotinine (D) in a pooled sample of an extract of brain supernatant from five rats 4 h after s.c. administration of 0.8 mg/kg of nonradiolabeled S(−)-nicotine by ion extraction chromatography and mass spectrometry.
A to D, ion extraction chromatograms (top) and electron impact mass fragmentation patterns (bottom) for nicotine (m/z 162, 133, 119), cotinine (m/z 176, 145, 98), nornicotine (m/z 148, 147, 119), and norcotinine (m/z 162, 134, 80), respectively.
To determine the time course of appearance and half-life of CNS metabolites, experiments were carried out with [2′-14C]nicotine to reliably quantitate the low levels of these metabolites in brain. Groups of rats were administered 0.8 mg/kg of nicotine containing 50 μCi of [2′-14C]nicotine. At 5, 30, 60, 240, 360, 480, 720, 960, and 1440 min after injection, brain supernatants from individual animals were obtained and analyzed by HPLRC. The half-lives of nicotine and its metabolites were calculated from the terminal slope of log concentration versus time graphs of each component. The results of these time course studies are shown in Figs.4 and 5. Figure 4 illustrates representative radiochromatographs obtained at the different time points analyzed. At the 5-min time point, only nicotine was observed in brain, whereas at 30 min, small amounts of both cotinine and nornicotine began to appear. These two metabolites were relatively greater in amount at the 60-min time point when compared with nicotine, and reached their highest levels between 1 and 4 h. In addition, the minor metabolite norcotinine was clearly apparent at 4 h. At 8 h, nicotine was virtually nonquantifiable. At 8 h postinjection, nicotine was completely cleared from the central compartment, whereas only residual levels of nornicotine were present. The cyclic lactam metabolites, cotinine and norcotinine, which have relatively longer half-lives than the basic amino compounds (nicotine and nornicotine), were still present in brain 12 h postinjection. Two very minor unidentified metabolites (A and B) appeared in brain at the 4-h time point but were barely detectable at 16 h.
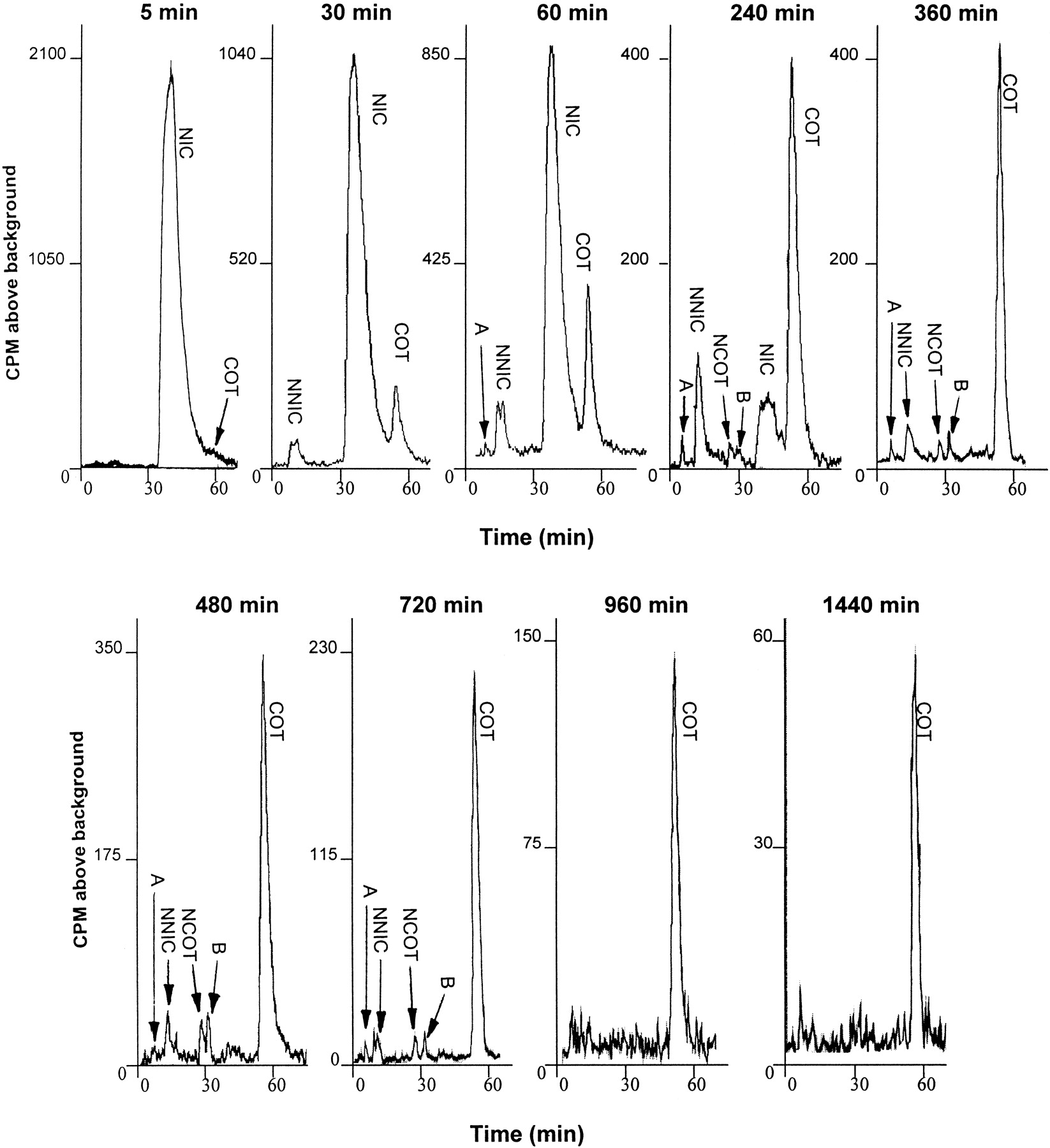
HPLRC chromatograms of brain supernatant from representative rats that have been injected s.c. with 0.8 mg/kg [2′-14C]nicotine (NIC; 50 μCi).
Brains were removed at 5, 30, 60, 240, 360, 480, 720, 960, and 1440 min after injection. Brain supernatants were obtained and either 1-ml (for 5-, 30-, 60-, 240-, and 360-min time points), or 2-ml (for 480-, 720-, 960-, and 1440-min time points) samples were injected onto a Partisil-10 C8 column (for details, see Materials and Methods). A and B, unidentified minor nicotine metabolites. NNIC, nornicotine; COT, cotinine; NCOT, norcotinine.

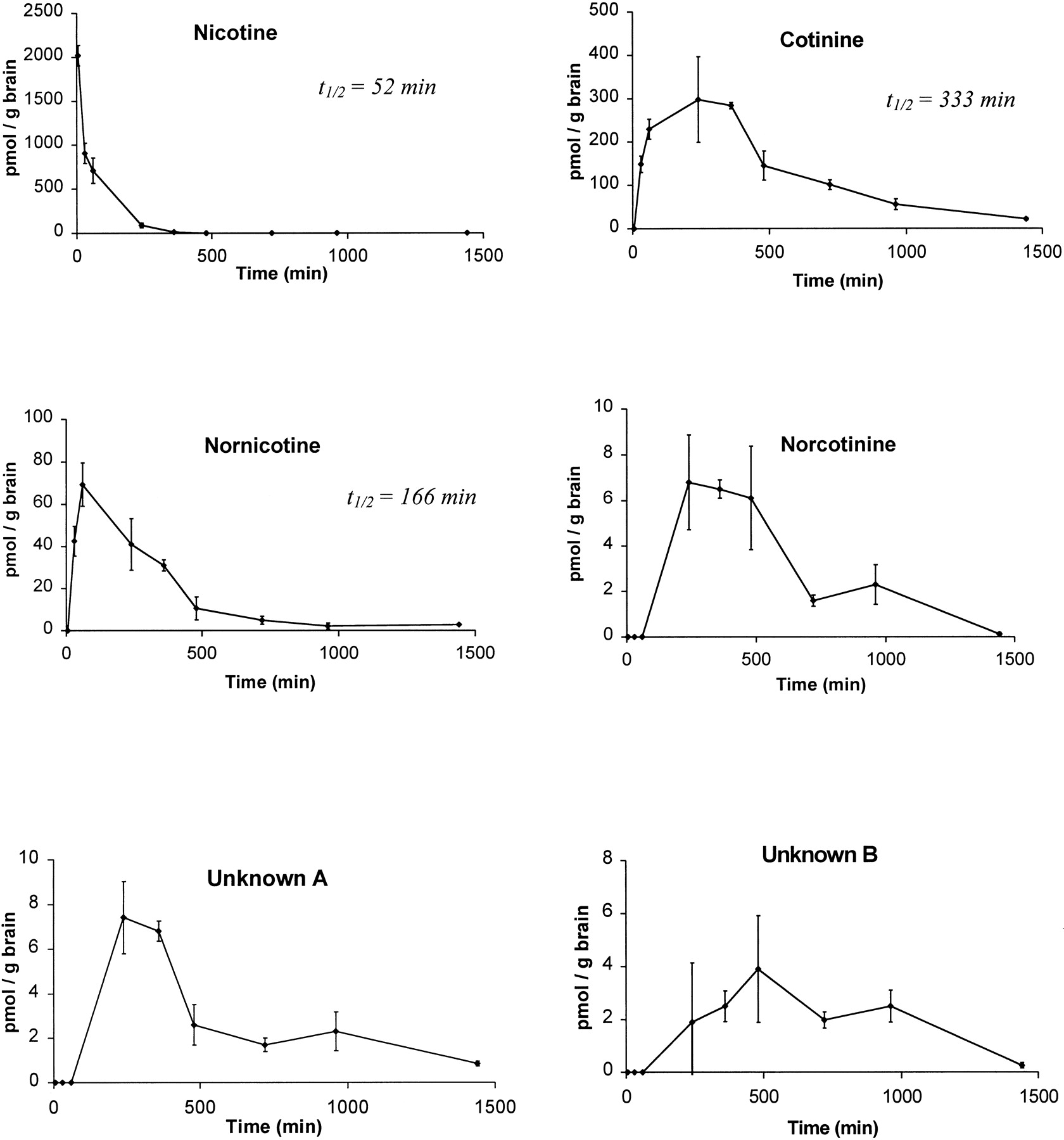
Brain time-concentration profiles for nicotine, cotinine, nornicotine, norcotinine, and two unidentified minor metabolites (A and B) obtained from a group of four rats (mean ± S.E.) that have been injected s.c. with 0.8 mg/kg [2′-14C]nicotine (50 μCi).
The half-lives for nicotine, cotinine, and nornicotine in brain are also shown.
Figure 5 illustrates time-concentration profiles for nicotine, cotinine, nornicotine, norcotinine, and the two unidentified metabolites A and B obtained from a group of four rats in which [2′-14C]nicotine was peripherally (s.c.) administered. Nicotine absorption from the s.c. injection site was rapid, such that at the 5-min time point, the concentration of nicotine in brain had already peaked. The time-concentration profile was clearly biphasic, showing an initial rapid distribution phase followed by a slower elimination phase. By 6 h, nicotine was almost completely cleared from brain. The nicotine T1/2was 58 min. The time-concentration profile for cotinine indicated that it was first detected at 30 min and peaked between 60 and 240 min, and was still present in significant amounts at 8 and 16 h postinjection. Cotinine was the major metabolite in brain over the entire time course studied and reached a peak concentration of 300 pmol/g brain tissue. Thus, cotinine entered the CNS quickly, but was eliminated from the CNS relatively slowly. The cotinineT1/2 was 5.5 h and was still detectable at 24 h postinjection.
The next most abundant metabolite was nornicotine. Nornicotine was not observed at 5 min, but peaked at 60 min with a maximal concentration of 70 pmol/g brain, and was still detectable at 12 h. The nornicotineT1/2 was 2.8 h, and its elimination rate was comparable with that for nicotine.
Norcotinine was a very minor metabolite and was not observed until 4 h, peaked at 7 pmol/g brain between 4 and 6 h, and was almost nonquantifiable by 24 h. The two unidentified metabolites, A and B, were also present in very minor amounts, similar to norcotinine. Thus, several metabolites in brain were detected that had relatively longer CNS residence times and half-lives than those of nicotine.
Discussion
S(−)-Cotinine is a major peripheral metabolite of nicotine and is able to pass the blood-brain barrier from the periphery (Appelgren et al., 1962; Schmiterlow et al., 1967; Deutsch et al., 1992; Crooks et al., 1997; Crooks and Dwoskin, 1997). The origin ofS(−)-cotinine in brain has not been elucidated and could arise from oxidative formation from nicotine locally in the brain, or from oxidative formation in the periphery followed by redistribution to the brain. It is generally accepted that brain is a poor metabolizing organ compared with liver, although the activity of certain metabolic enzymes have been detected in brain, including certain isozymes of cytochrome P-450 that are induced on chronic administration of nicotine (S. Miksys, E. Hoffman, and R. F. Tyndale, submitted). The relatively large amounts of cotinine found in brain would suggest that cotinine is formed in the periphery and then redistributed to the CNS. Nevertheless, one cannot rule out the formation, at least in part, of cotinine from nicotine locally in brain.
Compared with nicotine, the neuropharmacological effects ofS(−)-cotinine have not been widely investigated. In behavioral studies, S(−)-cotinine has been reported to alter responding for food in rats, Beagle dogs, and squirrel monkeys (Risner et al., 1985; Goldberg et al., 1989). However, the rate-increasing effect of S(−)-cotinine during fixed interval responding was not attenuated by the nicotinic receptor antagonist, mecamylamine (Goldberg et al., 1989), suggesting that this effect may not be nicotinic receptor mediated. Drug discrimination studies report generalization of S(−)-nicotine toS(−)-cotinine in rats and squirrel monkeys, but large doses of S(−)-cotinine were required (Goldberg et al., 1989;Tacada et al., 1989). Similar to nicotine, cotinine releases dopamine from its presynaptic terminals in striatum via a nicotinic receptor-mediated mechanism (Dwoskin et al., 1999b). The EC50 for S(−)-cotinine to evoke [3H]dopamine overflow from rat striatal slices was 30 μM. Furthermore, exposure to S(−)-cotinine resulted in nicotinic receptor desensitization (Dwoskin et al., 1999b). Thus, although the concentration of S(−)-cotinine in brain (0.3 μM) after acute nicotine administration in the present study is not within the concentration range found to be effective in the dopamine release assay, cotinine may accumulate in brain during chronic smoking due to its long residence time in brain and due to the fact that cotinine is also an alkaloidal constituent of tobacco (Crooks, 1999). Nevertheless, taking into account potential accumulation with chronic nicotine administration and the expected higher cotinine concentrations in brain from chronic smokers, it remains unlikely that the concentration of cotinine in brain will reach the levels that were found to be effective in the dopamine release assay. Thus, it appears unlikely that cotinine will contribute to the dopamine-mediated neuropharmacological effects of nicotine exposure. In the current studies, cotinine had a half-life of 333 min in rat brain after a single peripheral dose of 0.8 mg/kg nicotine. This is comparable with a plasma cotinine half-life of 294 to 318 min over a range of nicotine doses in the rat (Kyerematen et al., 1988).
The next most abundant metabolite found in brain in the present studies was nornicotine. It is important to note the concentration of nornicotine (0.07 μM) observed in brain after acute peripheral nicotine administration. Nornicotine was found to have a half-life of 166 min in rat brain (present study); this compares to a half-life of 198 min for this metabolite in plasma after i.v. administration of nicotine in the rat (Kyerematen and Vesell, 1991). It has been reported that the plasma half-life of nornicotine in smokers and nonsmokers after a single i.v. dose of nicotine is 7.2 and 8.5 h, respectively; these values are considerably higher than plasma half-lives for nornicotine observed in the rat (Kyerematen and Vesell, 1991). Nornicotine is a minor biotransformation product (0.4% in humans and 8% in rats) of nicotine in the periphery (Cundy and Crooks, 1984; Benowitz et al., 1991; Curvall and Kazemi Valla, 1993). In guinea pigs, only 1.6% of the total nicotine dose was detected as nornicotine in 24-h urine void (Cundy and Crooks, 1984). After administration of a single arterial dose of labeled nicotine to rats, it was found that nornicotine accounted for only 8% of total recovery of administered radioactivity (Curvall and Kazemi Valla, 1993). Therefore, it is intriguing that such significant amounts of this metabolite, relative to nicotine, are found in rat brain after acute peripheral nicotine administration. Thus, redistribution of metabolically formed nornicotine from the periphery to the brain appears to be an unlikely source of brain nornicotine, considering the very small amount of nornicotine detected as a peripheral nicotine metabolite. A more likely origin of nornicotine in brain is via local oxidativeN-demethylation of nicotine.
Nornicotine is pharmacologically active, and similar to nicotine, acts as an agonist at nicotinic receptors, which evoke the release of dopamine from its presynaptic terminal stores (EC50 = 1.0 μM; Teng et al., 1997). Nornicotine also causes nicotinic receptor desensitization (EC50 = 0.095 μM; Dwoskin et al., 1999c). In behavioral studies, nornicotine has been shown to have psychomotor effects that differ qualitatively from those ofS(−)-nicotine, in that behavioral sensitization was not apparent after chronic administration of nornicotine to rats (Dwoskin et al., 1999a). Similar to nicotine, nornicotine has reinforcing qualities, in that it maintains i.v. self-administration (Bardo et al., 1999). The present study demonstrates that concentrations of nornicotine in rat brain after acute administration of nicotine approach those that produce pharmacological effects. It is likely that nornicotine will accumulate in brain on chronic nicotine administration, based on the longer residence time of nornicotine in comparison to nicotine, and that accumulation will reach concentrations necessary for pharmacological effects to be observed. This is particularly relevant in human tobacco smokers, in that not only is nornicotine a biotransformation product of nicotine, but smokers are exposed to significant quantities of nornicotine present in commercial tobacco (15–20% total alkaloid content; Zhang et al., 1990; Curvall and Kazemi Valla, 1993). Thus, a combination of “alkaloidal” nornicotine and “metabolic” nornicotine may be sufficient to produce brain levels of nornicotine that contribute to the neuropharmacological effects of tobacco smoking.
The current study is the first to unequivocally identify and provide detailed information on the residence times and half-lives of several pharmacologically active nicotine metabolites in brain after acute nicotine exposure. Nevertheless, correlation of the current results in the rat with those in other species, including humans, may be limited by factors such as species differences in nicotine metabolism and distribution, as well as dose-related effects (i.e., only one dose of nicotine was used in this study).
An important pharmacokinetic characteristic of nicotine metabolites appearing in brain is that their residence times are all longer than that of nicotine. Therefore, the possibility exists that on repeated peripheral administration of nicotine, these metabolites have the potential to accumulate in brain. This is of particular significance with regard to the chronicity of nicotine exposure during tobacco smoking. Typically, a chronic smoker will smoke a cigarette every 20 min with intermittent inhalation of main-stream smoke. Delivery of nicotine to the CNS constitutes intermittent peaks, superimposed on a constant plasma level (Benowitz et al., 1991; Jacob and Benowitz, 1993). Because the residence time of nicotine in the CNS is short, its ability to accumulate is less likely than that of its metabolites, which have longer half-lives. Thus, it is possible that prolonged intermittent administration of nicotine, as is the case with tobacco smoking, may result in significantly higher levels of the slower effluxing metabolites. We hypothesize that slow effluxing metabolites that possess pharmacological activity may well play a major role in the neuropharmacology of tobacco smoking. It will be important to determine in future studies the ability of such metabolites to accumulate in brain after chronic nicotine administration.
Footnotes
-
Send reprint requests to: Dr. Peter A. Crooks, College of Pharmacy, University of Kentucky, Rose Street, Lexington, Kentucky: 40536-0082; E-mail: pcrooks{at}pop.uky.edu
-
This work was supported in part by National Institute of Drug Abuse Grants DA08656 and DA00399, and by the Tobacco and Health Institute, Lexington, Kentucky.
- Abbreviations used are::
- HPLRC
- high pressure liquid radiochromatography
- GC
- gas chromatography
- GC-MS
- gas chromatography-mass spectrometry
- CNS
- central nervous system
- Received April 16, 1999.
- Accepted September 8, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}