


Abstract
Milk thistle extract is one of the most commonly used nontraditional therapies, particularly in Germany. Milk thistle is known to contain a number of flavonolignans. We evaluated the effect of silymarin, on the activity of various hepatic drug-metabolizing enzymes in human hepatocyte cultures. Treatment with silymarin (0.1 and 0.25 mM) significantly reduced the activity of CYP3A4 enzyme (by 50 and 100%, respectively) as determined by the formation of 6-β-hydroxy testosterone and the activity of uridine diphosphoglucuronosyl transferase (UGT1A6/9) (by 65 and 100%, respectively) as measured by the formation of 4-methylumbelliferone glucuronide. Silymarin (0.5 mM) also significantly decreased mitochondrial respiration as determined by MTT reduction in human hepatocytes. These observations point to the potential of silymarin to impair hepatic metabolism of certain coadministered drugs in humans. Indiscriminate use of herbal products may lead to altered pharmacokinetics of certain drugs and may result in increased toxicity of certain drugs.
Milk thistle extract is one of the most commonly used nontraditional therapies, particularly in Germany. The annual sale of this product is about $180 million in Germany alone (Cowley, 1995). Milk thistle [Silybum marianum (L.) Gaertn. (Fam. Asteraceae)] is known to contain a number of flavonolignans, compounds that are produced in plants by radical coupling of a flavonoid and a phenylpropanoid (Dewick, 1997). A mixture of these flavonolignans, termed silymarin, was first isolated from an extract of milk thistle fruit [from which the pappus (feathery tuft) had been removed] some three decades ago. Silymarin is known to be composed of mainly silybin (about 50–70%), but also contains silychristin, silydianin, and other closely related flavonolignans (Wagner et al., 1985). A standardized extract of milk thistle contains at least 70% silymarin (Schulz et al., 1998; Foster and Tyler, 1999).
Silymarin/silybin is reported to protect the liver against CCl4, acetaminophen-, amanitin-, thioacetamide-, and d-galactosamine-mediated hepatotoxicity in rats (Schriewer et al., 1973; Vogel et al., 1984; Mourelle et al., 1989;Muriel et al., 1992; Chrungoo et al., 1997a,b). Silymarin has been reported to inhibit certain hepatic enzymes such as aminopyrine demethylase, benzopyrene hydroxylase, hexobarbital hydroxylase, and ethoxy coumarin O-deethylase in rats (Letteron et al., 1990). Silybin is primarily conjugated and excreted in the bile and urine in rats (Lorenz, 1982). Silymarin is known to deplete the pool of uridine diphosphoglucuronic acid (UDPGA)1 in hepatocytes and decrease glucuronidation of bilirubin in rats (Chrungoo et al., 1997b). The above observations would suggest that there would be a competition between silymarin and other drugs that are metabolized by various cytochrome P450 (CYP) enzymes or conjugated to a glucuronide in the liver. Our objective was to evaluate the effect of acute and chronic exposure of silymarin on the activity of CYP3A4 and uridinediphosphoglucuronosyl transferase (UGT1A6/9) in primary cultures of human hepatocytes.
Materials and Methods
Chemicals.
Williams E medium, medium supplements, dexamethasone, and insulin were obtained from BioWhittaker (Walkersville, MD). Gentamycin was obtained from Life Technologies Inc. (Grand Island, NY). Silymarin, 4-methylumbelliferone (4-MU), and testosterone were purchased from Sigma (St. Louis, MI). 6-β-Hydroxy (OH) testosterone was obtained from Steraloids (Wilton, NH). MTT was obtained from Bio-Rad (Richmond, CA).
Hepatocyte Culture and Treatment.
Hepatocytes were isolated from human donor livers by a three-step collagenase perfusion technique as described previously (Strom et al., 1996, 1998). The viability of the cells obtained was measured by Trypan blue exclusion test. Only cells that are >80% viable were used for further studies. The hepatocytes (1.5 × 106) were plated on six-well culture plates previously coated with rat-tail collagen. The hepatocytes were plated in Williams E medium supplemented with 0.1 μM dexamethasone, 0.1 μM insulin, 0.05% gentamicin, and 10% bovine calf serum. Cells were allowed to attach for 4 to 6 h, at which time the medium was replaced by a serum-free medium containing all the supplements listed above. The medium was changed every 24 h. The cells were maintained in culture at 37°C in a humid atmosphere containing 5% CO2 and 95% air throughout the experiment. For the acute study, varying concentrations of silymarin [0.1, 0.25, or 0.5 mM in dimethyl sulfoxide (DMSO); final concentration of DMSO in the medium being 0.1%] was added to the hepatocytes at 95 h of culture. For chronic study, varying concentrations of silymarin (0.1, 0.25, or 0.5 mM in DMSO) was added to the culture at 48 and 72 h after plating.
Enzyme Activity.
At 96 h, the hepatocytes were washed with fresh medium, incubated with fresh medium for an hour, and then exposed to a medium containing 350 μM testosterone or 100 μM 4-MU in DMSO (final concentration in the media was 0.1%) for 30 min. At this time point, the medium was harvested and stored at −20°C. 6-β(OH) testosterone (CYP3A activity) (Kostrubsky et al., 1999; Ramachandran et al., 1999) and 4-methylumbelliferone glucuronide (4-MUG) were measured by HPLC. The cells were harvested for protein determination.
MTT Reduction.
In a separate set of experiments, after chronic exposure (48 h) of the hepatocytes to silymarin at various (0.1, 0.25, and 0.5 mM) concentrations, 10% v/v of 5 mg/ml MTT was added to Williams E medium at 96 h of culture and incubated for 30 min. At 30 min, the medium was aspirated and washed with phosphate-buffered saline. Isopropanol (same volume as the medium) was added and shaken gently for 5 min. Two hundred microliters of this solution was transferred to a 96-well plate, and the absorbance was measured at 490 nM.
Statistical Analysis.
The significance of the effect of silymarin on the activity of various enzymes was determined by ANOVA and Duncan groupwise comparison at aP < .05.
Results and Discussion
Milk thistle is commonly used as a hepatoprotectant. Silymarin, the major component of milk thistle, is reported to inhibit nitric oxide production (Dehmlow et al., 1996), is a potent free radical scavenger and prevents lipid peroxidation (Valenzuela et al., 1985; Muriel and Mourelle, 1990), enhances the activity of hepatocyte RNA polymerase (Sonnenbichler et al., 1976), and complexes toxic free iron (Pietrangelo et al., 1995). Silymarin/silybin is reported to protect the liver against several hepatotoxins in rats. In randomized clinical trials in patients with acute viral hepatitis, silymarin either exerted no benefit (Kiesewetter et al., 1977) or accelerated clinical recovery (Magliulo et al., 1978). In a recent randomized placebo-controlled trial in patients with alcoholic cirrhosis, the earlier observation of increased survival with silymarin could not be confirmed (Pares et al., 1998).
Silymarin has been hypothesized to inhibit the activity of several CYP enzymes in rats. Silymarin has been reported to decrease bilirubin conjugation in patients with liver cirrhosis (Salmi and Sarna, 1982). Based on studies in rats, it has been hypothesized to impair glucuronidation (Chrungoo et al., 1997b). Silymarin is primarily conjugated and excreted in the bile and urine. We anticipated potential interactions between silymarin and several coadministered drugs. We evaluated the potential of silymarin to alter the metabolism of other drugs by measuring the activity of CYP3A4, the major phase 1 enzyme, and UGT, the major phase 2 enzyme in primary cultures of human hepatocytes.
Primary cultures of human hepatocytes are valuable models to evaluate the regulation of chemicals on hepatic drug-metabolizing enzymes. We have used this system to evaluate the effects of rifampin, taxol, and troglitazone previously in our laboratory (Kostrubsky et al., 1997,1999; Ramachandran et al., 1999). Use of human hepatocytes obviates the need for extrapolation of in vivo animal studies to humans.
Figure 1 illustrates the activity of CYP3A4 in the hepatocytes as measured by the formation of 6-β(OH) testosterone in untreated or chronic silymarin treated (0.1 or 0.25 mM) cultures. The formation rate of 6-β(OH) testosterone in the untreated cells was 0.04 nmol/min/mg of protein. Silymarin at both concentrations significantly decreased CYP3A4 activity. Figure2A illustrates the activity of UGT as measured by the formation of 4-MUG after acute treatment (1 h) of silymarin. The rate of formation of 4-MUG was 0.35 nmol/min/mg of protein in untreated cells. Phenobarbital marginally (less than 2-fold) increased the activity of UGT1A6/9, while silymarin drastically reduced the activity of UGT in these hepatocytes. Figure 2B illustrates the activity of UGT after chronic treatment (for 48 h) of the hepatocytes with silymarin. Chronic treatment with silymarin significantly reduced the activity of UGT1A6/9 at 0.1 mM and completely inhibited the formation of 4-MUG at 0.25 and 0.5 nM.
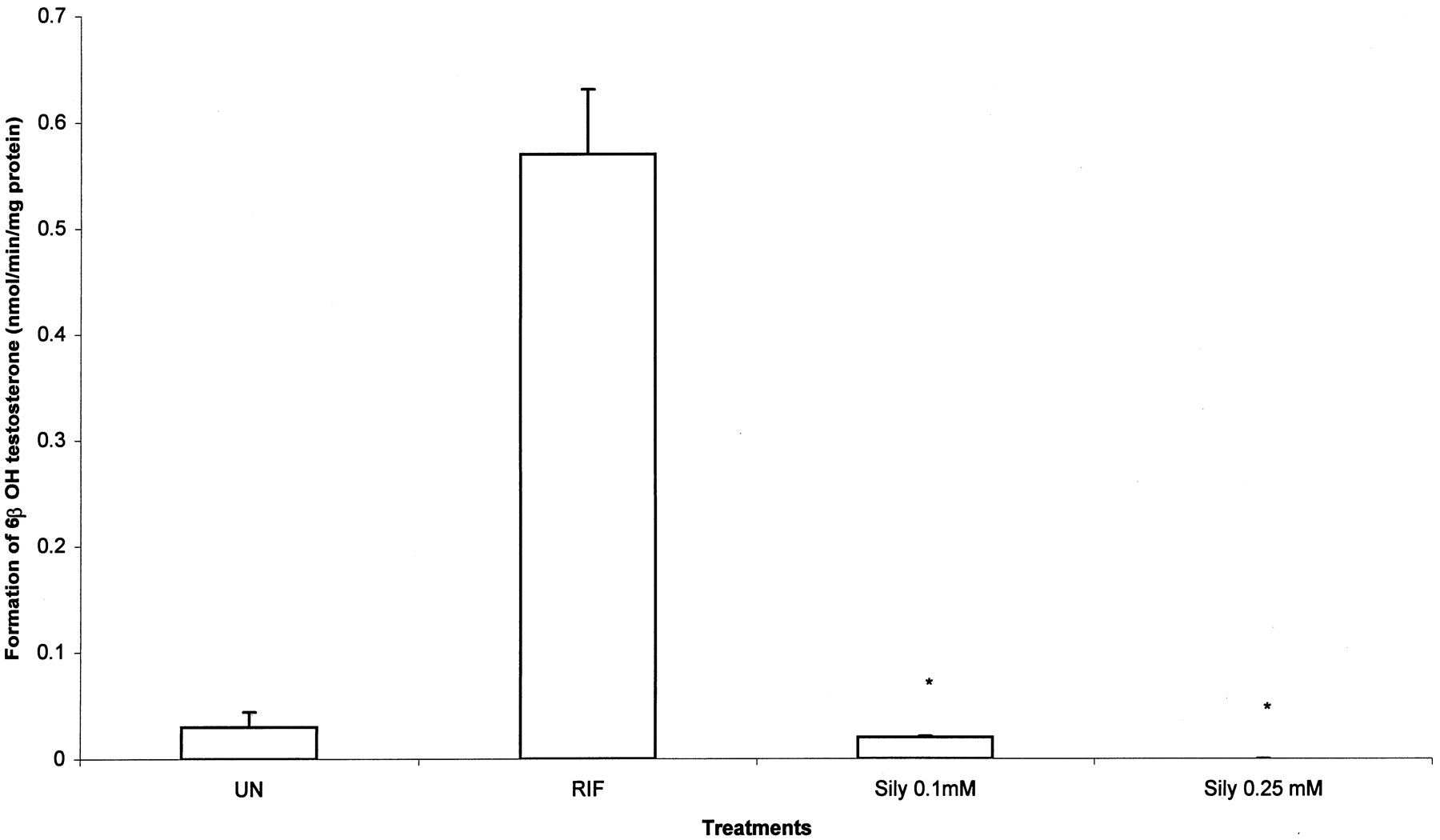
Effect of silymarin on CYP3A4 activity in primary cultures of human hepatocytes.
The formation rate of 6β(OH) testosterone was measured in primary cultures of human hepatocytes that were not treated or treated for 48 h with rifampin, or treated for 48 h with silymarin (0.1 and 0.25 mM). ∗, indicates significantly different from untreated cells.

Effect of acute silymarin (A) and chronic silymarin (B) on glucuronidation in primary cultures of human hepatocytes.
The formation rate of 4-MUG was measured in untreated, phenobarbital-treated (2 mM for 48 h), acute silymarin-treated (0.1 or 0.25 mM for 1 h), or chronic silymarin-treated (0.1, 0.25, or 0.5 mM for 48 h). ∗, indicates significantly different from untreated cells; ∗∗, indicates significantly different from phenobarbital-treated cells.
Our studies clearly document the potential of silymarin to inhibit the metabolism of substrates of CYP3A4 and UGT1A6/9 in humans. CYP3A enzymes are responsible for the metabolism of several drugs. UGT is the primary phase 2 enzyme responsible for the metabolism of several drugs. The exact reason(s) for the reduced activity of CYP3A4 in silymarin-treated cells is not clear at this time. Depletion of UDPGA and competition for the UGT may be the mechanisms involved in the reduction in UGT activity in presence of silymarin as measured in this study. The observation that UGT1A6/9 activity is lower after acute and chronic silymarin treatment would suggest that down-regulation of UGT1A6/9 is not likely a major mechanism involved. Additional ongoing studies will address the mechanistic basis for the observations presented here. Our study indicates that the use of silymarin with other drugs that are conjugated by UGT1A6/9 will lead to a reduction in the clearance of certain drugs. In addition, there is also a potential for increased toxicity due to increased formation of toxic metabolite(s) of coadministered drugs in the presence of silymarin. The toxicity of acetaminophen is increased in the presence of drugs that are also conjugated (Kostrubsky 1997).
Figure 3 illustrates MTT reduction by cells exposed to phenobarbital or silymarin at various concentrations. While phenobarbital (2 mM) or 0.1 and 0.25 mM silymarin did not have any effect on MTT reduction, silymarin at 0.5 mM significantly decreased the reduction of MTT.

Toxicity of silymarin in primary cultures of human hepatocytes.
Toxicity was measured by MTT reduction and expressed as percentage of control in untreated, phenobarbital, or 0.1, 0.25, or 0.5 mM silymarin treated for 48 h.
Despite the recent widespread use of herbal drugs, limited information is available on the safety and efficacy of herbal products used alone or in combination with prescription drugs. Further studies are needed immediately to characterize the safety and efficacy of herbal products in humans. In the meantime, caution must be exercised in prescribing drugs to patients who are on herbal supplement. Patients and health care professionals must be encouraged to discuss the use of herbs and be educated about the potential interactions between herbs and drugs.
Footnotes
-
Send reprint requests to: Raman Venkataramanan, Ph.D., F.C.P., Professor of Pharmaceutical Sciences and Pathology, 718 Salk Hall, University of Pittsburgh, Pittsburgh, PA 15261. E-mail:rv+{at}pitt.edu
-
This work was supported in part by University of Pittsburgh Pharmacy Associates grant from the School of Pharmacy (R.V., B.J.K.), by Pfizer undergraduate summer Research Fellowship (B.J.K.), and by National Institutes of Health N01-DK-9-2310 (S.C.S.).
- Abbreviations used are::
- UDPGA
- uridine diphosphoglucuronic acid
- UGT
- uridine diphosphoglucuronosyl transferase
- CYP
- cytochrome P450
- 4-MU
- 4-methylumbelliferone
- 4-MUG
- 4-methylumbelliferone glucuronide
- MTT
- (3-(4,5-dimethylthiazol-2yl) 2,5-diphenyltetrazolium bromide, Thiazolyl blue
- DMSO
- dimethyl sulfoxide
- OH
- hydroxy
- Received June 20, 2000.
- Accepted July 31, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}