


Abstract
The antipsychotic agent risperidone, is metabolized by different cytochrome P-450 (CYP) enzymes, including CYP2D6, to the active 9-hydroxyrisperidone, which is the major metabolite in plasma. Two enantiomers, (+)- and (−)-9-hydroxyrisperidone might be formed, and the aim of this study was to evaluate the importance of CYP2D6 and CYP3A4/CYP3A5 in the formation of these two enantiomers in human liver microsomes and in recombinantly expressed enzymes. The enantiomers of 9-hydroxyrisperidone were analyzed with high pressure liquid chromatography using a chiral α-1 acid glycoprotein column. A much higher formation rate was observed for (+)-9-hydroxyrisperidone than for (−)-9-hydroxyrisperidone in microsomes prepared from six individual livers. The formation of (+)-9-hydroxyrisperidone was strongly inhibited by quinidine, a potent CYP2D6 inhibitor, whereas ketoconazole, a CYP3A4 inhibitor, strongly inhibited the formation of (−)-9-hydroxyrisperidone. Recombinant human CYP2D6 produced only (+)-9-hydroxyrisperidone, whereas a lower formation rate of both enantiomers was detected with expressed CYP3A4 and CYP3A5. In vivo data from 18 patients during treatment with risperidone indicate that the plasma concentration of the (+)-enantiomer is higher than that of the (−)-enantiomer in extensive metabolizers of CYP2D6. These findings clearly suggest that CYP2D6 plays a predominant role in (+)-9-hydroxylation of risperidone, the major metabolic pathway in clinical conditions, whereas CYP3A catalyzes the formation of the (−)-9-hydroxymetabolite. Further studies are required to evaluate the pharmacological/toxic activity of both enantiomers.
Risperidone is a relatively new atypical antipsychotic with a potent serotonin 5-HT2 and a moderate dopamine D2 antagonistic activity (Leysen et al., 1988). It is effective in the treatment of both positive and negative symptoms of schizophrenia and has a lower potential to cause extrapyramidal symptoms compared with classical antipsychotics (Chouinard and Arnott, 1993).
Risperidone is metabolized mainly by 9-hydroxylation and to a lesser extent by N-dealkylation and 7-hydroxylation (Mannens et al., 1993). 9-Hydroxyrisperidone is the major metabolite in plasma and is claimed to be equipotent with the parent compound in terms of dopamine receptor affinity and hence contribute to the overall therapeutic effect of risperidone. The plasma concentrations of the parent compound plus the 9-hydroxymetabolite has therefore been reported as the “active moiety” in several studies (Megens et al., 1994; Van Beijsterveldt et al., 1994; Schotte et al., 1996).
A single-dose kinetic study in healthy subjects showed that the formation of 9-hydroxyrisperidone is predominantly catalyzed by the polymorphic CYP2D6 (Huang et al., 1993). A correlation betweenCYP2D6 genotype and the plasma concentration ratio of risperidone and 9-hydroxyrisperidone was recently reported in patients on chronic treatment with risperidone (Scordo et al., 1999).
Furthermore, in vivo (Bork et al., 1999; Spina et al., 2000) and in vitro experiments in human and rat liver microsomes and recombinantly expressed enzymes (Fang et al., 1999) revealed that not only CYP2D6 but also CYP3A are involved in the metabolism of risperidone. The in vitro finding of the CYP3A4-mediated metabolic pathway is not necessarily indicative that this enzyme is important since a risperidone concentration of 100 μM was used. This concentration is unlikely to be clinically relevant when considering that plasma risperidone levels observed in patients treated with therapeutic doses of 4 to 8 mg/day range approximately between 5 and 100 nM (Olesen et al., 1998;Balant-Gorgia et al., 1999; Scordo et al., 1999; Spina et al., 2001a).
The 9-hydroxylation results in the formation of a chiral carbon atom yielding two enantiomers, (+)- and (−)-9-hydroxyrisperidone (Fig.1). To date, there is no information about differences in the pharmacological activity of the two enantiomers, and no information about which CYP enzymes are involved in their formation. Since CYP2D6 seems to be an important enzyme in the metabolism of risperidone, it is possible that the formation of 9-hydroxyrisperidone is highly stereoselective as shown for other CYP2D6 substrates like nortriptyline (Dahl et al., 1991) and mianserin (Dahl et al., 1994). The aim of the present study was to investigate the stereoselective formation of 9-hydroxyrisperidone using human liver microsomes and recombinantly expressed enzymes. For this purpose, a chiral HPLC1 method was developed and used also to measure the enantiomers in plasma samples from patients of known CYP2D6 genotype stabilized on risperidone treatment.

Chemical structures of risperidone and the formed 9-hydroxyrisperidone.
Asterisk indicates the chiral center.
Experimental Procedures
Materials.
Risperidone, 9-hydroxyrisperidone (racemate), (+)- and (−)-9-hydroxyrisperidone, and ketoconazole were kindly provided by Janssen Biotech NV (Olen, Belgium). Quinidine was provided by Apoteksbolaget (Stockholm, Sweden). β-Nicotinamide adenine dinucleotide phosphate, reduced form (β-NADPH) was purchased from Sigma (St. Louis, MO). All other chemicals were of analytical grade and purchased from Merck (Darmstadt, Germany).
Human Liver Microsomes.
Microsomes were prepared as described by von Bahr et al. (1980) from six healthy organ donors (HL-47, -49, -51, -54, -55, and -67) from a liver bank established at the Department of Clinical Pharmacology, Huddinge University Hospital (approved by the Ethical Committee at Huddinge University Hospital). The protein content was estimated according to Lowry et al. (1951). The microsomes were stored in 50 mM potassium phosphate buffer (pH = 7.4) at −80°C until use. DNA was extracted from all livers except HL-47, using QIAamp Tissue DNA kit (Qiagen, Hilden, Germany) and genotyped for the CYP2D6*3 and*4 alleles according to Heim and Meyer (1990).
Enzyme Kinetics in Human Liver Microsomes.
Human liver microsomes were incubated in 100 mM potassium phosphate buffer (pH = 7.4) using 0.5 mg of microsomal protein in a final volume of 500 μl. Incubations were performed at 37°C for 15 min in the presence of 1 mM NADPH. The reaction was stopped by adding 500 μl of 0.5 M sodium hydroxide and stored frozen at −20°C until analysis. The linearity with respect to microsomal protein concentration and incubation time was first evaluated and the formation kinetics of both enantiomers was determined by incubating risperidone at ten different concentrations (0.1, 0.5, 1, 5, 10, 25, 50, 100, 200, and 300 μM) in microsomes from six livers. The experiments were performed in duplicate, and the reaction velocities were calculated in the unit of picomoles of product formed per minute and milligram of microsomal protein.
Inhibition Study in Human Liver Microsomes.
Quinidine and ketoconazole were used as selective inhibitors of CYP2D6 and CYP3A4, respectively (Rodrigues, 1999). Quinidine was dissolved in water and ketoconazole in methanol, which was evaporated before incubation. The experiments were performed in microsomal preparations from livers HL-47, HL-51, HL-55, and HL-67 at three different concentrations of risperidone (0.25, 5, and 100 μM) in the presence of the inhibitors at four different concentrations (0.01, 0.1, 1, and 10 μM). Quinidine and ketoconazole were also incubated without risperidone under the same condition to ensure that the inhibitors or their metabolites would not interfere with quantification of the 9-hydroxymetabolites. One peak, obtained at quinidine concentrations of 10 μM, interfered with the (−)-enantiomer of 9-hydroxyrisperidone and data are therefore not reported.
Assay with cDNA-Expressed Human P-450s.
Microsomes from cDNA-transformed yeast cells overexpressing yeast reductase as well as human CYP2D6 or CYP3A4 were produced in our laboratory as described by Bylund et al. (2000). Cytochromeb5 was kindly provided by AstraZeneca R&D, Mölndal, Sweden. Expression levels and catalytic activity of cDNA-expressed human CYP2D6 in yeast have been confirmed and reported by Oscarson et al. (1997). The yeast cDNA-expressed human CYP3A4 with additional human b5 have aVmax of 1 pmol of formed product per minute and per picomole of CYP and a Km of around 500 μM with regards to testosterone 6-β-hydroxylase activity. Expression levels and catalytic activities for CYP3A4 with and without cytochrome b5 have been confirmed in our laboratory and reported by Andersson et al. (2001). Since the yeast-expressed human CYP3A4 with additional cytochromeb5 were incubated simultaneously and had no detectable risperidone 9-hydroxylase activity, SUPERSOMES containing human reductase as well as CYP3A4 and CYP3A5 were purchased from GENTEST Corporation (Woburn, MA). This product is a mixture of microsomes prepared from insect cells (BTI-TN-5B1–4) expressed from human CYP3A4 and CYP3A5 cDNA using a baculovirus expression system. Microsomal protein concentrations and CYP content were provided by the manufacturer.
Enzyme Kinetics in Yeast Microsomes.
The optimal protein and time dependence were evaluated also for recombinantly CYP2D6, CYP3A4, or CYP3A5 expressed microsomes and incubation conditions used were the same as those used for human liver microsomes, except for a preincubation of 3 min. The formation kinetics of both enantiomers of 9-hydroxyrisperidone were determined by incubating risperidone at eight different concentrations (0.25, 0.5, 1, 5, 10, 25, 50, and 100 μM) in yeast-expressed human CYP2D6, whereas the activity of CYP3A4 and CYP3A5 were determined by incubating risperidone at 100 μM. Reaction velocities were calculated in the unit of picomoles of product formed per minute and per picomoles of P-450.
HPLC Analysis of (+)- and (−)-9-Hydroxyrisperidone.
After thawing, the samples were extracted with 3 ml of diisopropyl ether/isoamyl alcohol 97:3 (v/v) for 10 min followed by centrifugation at 3000g for 5 min. The organic phase was transferred to a new test tube and extracted with 100 μl of potassium phosphate buffer (25 mM pH = 3.5) for 5 min. After centrifugation, the organic phase was discarded, and the residue was washed with 500 μl ofn-heptane. The organic phase was discarded and 30 μl of the sample was injected onto a chiral α-1 acid glycoprotein column (100- × 4.0-mm i.d., 5 μm, ChromTech AB, Stockholm Sweden). The mobile phase consisted of potassium phosphate buffer, 50 mM, pH = 6.5, and methanol 85:15 (v/v). The flow rate was 0.9 ml/min and the detection wavelength was 278 nm. The retention times for the enantiomers of 9-hydroxyrisperidone were 5.2 min for the (−)-form and 6.5 min for the (+)-form, respectively (Fig.2). Standard curves using the racemate were analyzed without internal standard in the concentration range of 50 to 2500 nM (r > 0.99 for both enantiomers). The low available amount of the separate enantiomers could only be used to check the accuracy in the stereospecific area determinations and to identify the retention times in the HPLC assay. No evidence for interconversion of the enantiomers was seen during the analytical procedure. The extraction recovery was 70%. The reproducibility and accuracy of the assay were determined by analyzing a set of quality control samples at the concentrations of 50 and 1000 nM for both enantiomers. The limit of quantification was 25 nM for each enantiomer. The intra- and interday variations at 25 nM were 5.7 and 6.8%, respectively. The corresponding values at 1000 nM were less than 5.7%.

HPLC chromatograms.
A, the chiral separation of racemic (−)-9-hydroxyrisperidone (1) and (+)-9-hydroxyrisperidone (2) and risperidone (3); B, the in vitro formation of the two enantiomers in human liver microsomes after incubation with risperidone (HL-47); C, plasma sample from a patient (number 15) during treatment with risperidone.
Plasma Analysis.
Chiral separation of 9-hydroxyrisperidone was performed in plasma samples from patients participating in previously published studies (Scordo et al., 1999; Spina et al., 2001b). The subjects consisted of 18 schizophrenic patients (12 males and 6 females, aged 27–60 years) treated with risperidone (4–9 mg/day) for at least 4 weeks. Plasma samplings were performed 12 h after the bedtime dose, just before the morning dose. No other medications were administered except benzodiazepines in some subjects. Risperidone and total (achiral) 9-hydroxyrisperidone concentrations in plasma were analyzed by HPLC (Avenoso et al., 2000) in all patients. Genotyping for the main detrimental (CYP2D6*3, *4, *5, *6) and for the duplicated (*2x2) CYP2D6 alleles was performed in 12 of the 18 patients as described by Scordo et al. (1999). The extraction procedure for plasma was the same as described for human liver microsomes after adding 500 μl of 0.5 M NaOH to 1.0 ml of plasma. Plasma concentrations of the separate enantiomers were calculated from the total concentration of 9-hydroxyrisperidone earlier determined and the peak area ratio of the (+)- and (−)-enantiomers.
Data Analysis and Statistics.
The formation kinetics of (+)- and (−)-9-hydroxyrisperidone in human liver microsomes and cDNA-expressed CYPs was described by one of the following models.
A one-enzyme Michaelis-Menten model:
For the in vivo data, Student's t test was used for statistical calculations of the plasma levels of the separate enantiomers of 9-hydroxyrisperidone between different CYP2D6genotype groups. A p value of 0.05 or less was regarded as statistically significant.
Results
Enantioselective Formation of 9-Hydroxyrisperidone in Human Liver Microsomes.
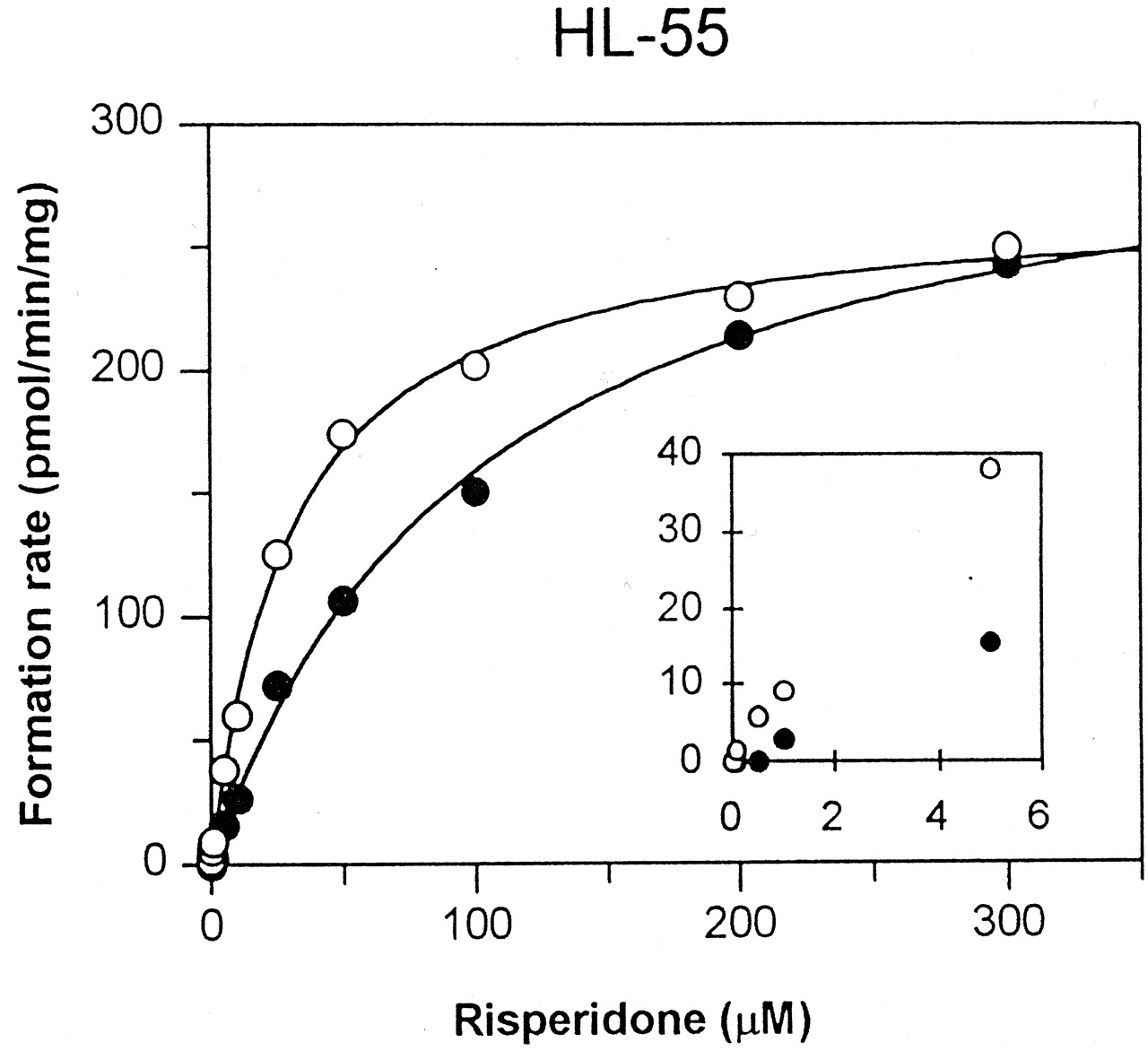
The rate of formation of (+)-9-hydroxyrisperidone was higher than that of (−)-9-hydroxyrisperidone at risperidone concentrations lower than 100 μM in all six livers. A more pronounced difference was observed at substrate concentrations below 5 μM (Fig.3). The formation of (+)- and (−)-9-hydroxyrisperidone followed Michaelis-Menten kinetics (Fig. 3). Individual Eadie-Hofstee plots for the (−)-9-hydroxylation showed a monophasic profile in all six livers. The (+)-9-hydroxylation showed a biphasic profile in four livers and monophasic profile in two livers due to undetectable formation of metabolites at lower substrate concentrations. This indicates that several enzymes, high- and low-affinity enzymes, could be involved in the (+)-hydroxylation. The computer-derived Km1 values calculated with the two-enzyme model are however very low (0.26 μM ± 0.18; mean ± S.D) in relation to the substrate concentration range (0.1–300 μM) and might therefore not be considered as reliable (Table 1). The limit of the chiral assay did not allow substrate concentrations lower than 100 nM. No relation between CYP2D6 genotype and the rate of formation of the enantiomers was observed.

A representative Michaelis-Menten curve for the formation of 9-hydroxyenantiomers in one microsomal preparation (HL-55).
Open circles indicate the formation rate of (+)-9-hydroxyrisperidone, and closed circles indicate those for (−)-9-hydroxyrisperidone. The formation rate for the enantiomers at a substrate concentration of 0.25 μM is enlarged and inserted in Michaelis-Menten curve.
Apparent Michaelis-Menten kinetic parameters of the formation of (+)- and (−)-9-hydroxyrisperidone in human liver microsomes
Experiments with Chemical Inhibitors.
Figure 4 shows the results after inhibition by quinidine and ketoconazole at three different concentrations, 0.25, 5, and 100 μM risperidone. Data were excluded in one liver (HL-51) at a concentration level of 0.25 μM due to a too low formation of the enantiomers. Quinidine strongly inhibited the formations of (+)-9-hydroxyrisperidone at risperidone concentration levels of 0.25 and 5 μM, whereas no inhibitory effect was found in the formation of the (−)-enantiomer. The (+)-9-hydroxylation at risperidone concentration of 100 μM was only slightly inhibited by quinidine. On the other hand, ketoconazole strongly inhibited the formation of (−)-9-hydroxyrisperidone in the range 0.25 to 100 μM whereas inhibition of (+)-9-hydroxylation by ketoconazole was found only at 100 μM. This clearly shows that (+)- and (−)-9-hydroxylation are specifically catalyzed by CYP2D6 and CYP3A4, respectively. At a higher substrate concentration like 100 μM, the stereospecificity is lost.

The stereoselective inhibition by quinidine and ketoconazole on the 9-hydroxylation at a risperidone concentration of 0.25, 5, and 100 μM in human liver microsomes.
Open circles indicate the velocities of (+)-9-hydroxyrisperidone and closed circles indicate those for (−)-9-hydroxyrisperidone. Data are mean ± S.D. of three livers at a concentration of 0.25 μM and of four livers at 5 and 100 μM.
Experiments with Recombinant cDNA Human Microsomes.
The (+)-9-hydroxyrisperidone, but not the (−)-form was detectable after incubation with risperidone using yeast-expressed human CYP2D6 (Table 2). The formation of (+)-9-hydroxyrisperidone followed the one-enzyme Michaelis-Menten kinetics and Eadie-Hofstee plot showed a monophasic profile. The estimated apparent Km andVmax were 1.4 μM and 2.21 pmol/min/pmol of P-450, respectively. Both (+)- and (−)-9-hydroxyrisperidone were detected when risperidone was incubated with SUPERSOME, recombinant human microsomes of CYP3A4 and CYP3A5. The CYP3A4 mediated formation of (+)- was almost 4 times lower than that for (−)-9-hydroxyrisperidone, 0.17 and 0.64 pmol/min/pmol P-450, respectively (Table 2). Very similar results were obtained with the CYP3A5 microsomes.
Formation of (+)- and (−)-9-hydroxyrisperidone in cDNA-expressed microsomes at different concentrations of risperisone
Analysis of the 9-Hydroxyenantiomers in Patient Plasma Samples.
Plasma concentrations of (+)- and (−)-9-hydroxyrisperidone were detected in 17 of 18 patients as shown in Table3. Chiral separation showed that (+)-9-hydroxyrisperidone was the predominant enantiomer present in plasma from all patients except one (number 11) with two mutantCYP2D6 alleles (CYP2D6*4/*5), who had no detectable (+)-hydroxyenantiomer. One patient (number 16) had a concentration of total 9-hydroxyrisperidone of 33 nM, which is too low for an accurate chiral quantification, and the ratio is therefore not calculated. The mean concentration ratio of (+)- to (−)-9-hydroxyrisperidone in patients heterozygous for mutatedCYP2D6 alleles (*1/*4, n = 6 and1*/5*, n = 1) was significantly lower than in patients with no mutated alleles (*1/*1,n = 3) (2.50 ± 0.65 versus 3.86 ± 0.48,p = 0.013). No significant difference in plasma concentration of (+)- or (−)-9-hydroxyrisperidone was found between these two groups.
Steady-state plasma concentrations of risperidone and enantiomers of 9-hydroxyrisperidone (9-OH) in 18 patients treated with risperidone and CYP2D6 genotype (n = 11)
Discussion
This is the first study showing that the formation of 9-hydroxyrisperidone is highly stereoselective with respect to the activity of CYP2D6 and CYP3A4 both in human liver microsomes and in patients. Fang et al. (1999) have shown that CYP2D6, CYP3A4, and/or CYP3A5 catalyze the risperidone 9-hydroxylation in human liver microsomes, recombinant-expressed enzymes, and rat liver microsomes at a relatively high substrate concentration (100 μM). The plasma levels of risperidone observed in patients treated with therapeutic doses range approximately between 5 and 100 nM. In the present study, we examined the kinetics at substrate concentrations slightly higher than therapeutic levels. The importance of CYP2D6 and CYP3A4 was evaluated with quinidine and ketoconazole as specific in vitro inhibitors.
A higher rate of formation was observed for (+)-9-hydroxyrisperidone than for (−)-9-hydroxyrisperidone in microsomes from six individual livers. The difference was even more pronounced in the lower substrate concentration range, 0.1 to 1 μM (Fig. 3). This suggests that (+)-9-hydroxylation is the predominant reaction in clinical situation. The (−)-9-hydroxylation fitted the one-enzyme model suggesting that this reaction is catalyzed by one or two similar enzymes. The (+)-9-hydroxylation has good fit for the two-enzyme model in four of six livers indicating that at least two enzymes (high-affinity and low-affinity enzymes) with overlapping activity could be involved. Kinetic data obtained from two livers fitted the one-enzyme model due to undetectable concentrations of the formed metabolites in the low concentration range. Although we have no clear explanation for this discrepancy, it seems that the enzymatic activity was lower in the microsomes prepared from these two livers. Immunoblotting of individual variability of the CYP2D6 and CYP3A4 protein levels might explain this difference, but this was unfortunately not possible in this study.
The results from the inhibition study using quinidine and ketoconazole revealed the involvement of only CYP2D6 in the (+)-9-hydroxylation at low-substrate concentrations, whereas both CYP2D6 and CYP3A4 seem to be involved at higher substrate concentrations. These findings support the kinetic study, showing a two-enzyme model for (+)-9-hydroxyrisperidone. Because CYP3A4 is regarded as a low-affinity and high-capacity enzyme whereas CYP2D6 as a high-affinity and low-capacity enzyme (Venkatakrishnan et al., 1999), it is reasonable to conclude that the first enzyme in the two-enzyme model is CYP2D6 and the second enzyme is CYP3A4. In addition, it seems likely that inhibition of (+)-hydroxylation by ketoconazole is ascribable to alter the effect of CYP3A4 after saturation of CYP2D6. On the other hand, since the (−)-9-hydroxylation was inhibited only by ketoconazole, but not quinidine, it seems likely that CYP3A4 is quantitatively the most important enzyme in the (−)-9-hydroxylation. This finding is also in agreement with the kinetic study fitting the one-enzyme model for the (−)-form.
The clear stereoselective inhibition by ketoconazole and quinidine was demonstrated after incubations with risperidone at 0.25 and 5 μM, whereas no such difference was seen at a substrate concentration of 100 μM. When the formation rate of 9-hydroxyrisperidone was calculated from the sum of both enantiomers, our results for inhibition study using 100 μM risperidone was quite similar to a previous in vitro study showing that the degree of inhibition by ketoconazole is greater than that by quinidine (Fang et al., 1999). The Km value is recommended as a substrate concentration for inhibition studies (Rodrigues, 1999), and 100 μM corresponds to theKm value for the (−)-9-hydroxylation in this study. Notwithstanding, it should be noted that 100 μM is much higher than the risperidone plasma levels in patients (0.01–0.1 μM).
To confirm the findings from the inhibition study, we examined recombinant cDNA-expressed human CYP2D6, CYP3A4, and CYP3A5. Total turnover values for the formation of both enantiomers with CYP2D6, CYP3A4, and CYP3A5 are similar to a previous study at the same substrate concentration (100 μM) (Fang et al., 1999), although we cannot simply compare these values since different kinds of expression systems were used. Only (+)-9-hydroxylation and not (−)-9-hydroxylation was detected with yeast-expressed CYP2D6, suggesting the involvement of CYP2D6 only in the (+)-9-hydroxylation. Meanwhile, both (+)- and (−)-9-hydroxylation were detected with recombinant cDNA-expressed CYP3A4 and CYP3A5. These findings confirm the results from the inhibition study with human liver microsomes and support our hypothesis that the high- and low-affinity enzymes in the two-enzyme model of (+)-9-hydroxylation are CYP2D6 and CYP3A4, respectively.
Both (+)- and (−)-9-hydroxyrisperidone were found in plasma of patients treated with risperidone. It was also observed that the (+)-enantiomer was predominant in plasma of patients with normal CYP2D6 activity, whereas it was not detected in one patient homozygous for two mutated CYP2D6 alleles (*4/*5). The ratio of (+)-9- to (−)-9-hydroxyrisperidone in the patients with no mutated alleles (*1/*1) was significantly higher (p < 0.02) than the ratio in patients carrying one mutated allele (*1/*4 or 1*/5*). This is in agreement with the in vitro findings that the rate of (+)-9-hydroxylation is higher than the rate of (−)-hydroxylation, and that risperidone (+)-9-hydroxylation is catalyzed predominantly by CYP2D6. Thus, in vitro techniques are useful to predict CYP enzymes responsible for the drug metabolism in vivo.
It has been suggested that 9-hydroxyrisperidone has a potency similar to that of the parent compound in receptor binding assays (Megens et al., 1994; Van Beijsterveldt et al., 1994; Schotte et al., 1996). Therefore the sum of the risperidone and 9-hydroxyrisperidone levels in plasma (described as the active moiety), has been used to examine the clinical effect-plasma concentration relationship (Olesen et al., 1998;Spina et al., 2001a), but no information about pharmacological activity of the two 9-hydroxyrisperidone enantiomers is available today. Therefore, further studies on the pharmacological properties and metabolism of the two enantiomers are needed.
In conclusion, this study shows that CYP2D6 plays a predominant role in the (+)-9-hydroxylation of risperidone, the major metabolic pathway both in vivo and in vitro, whereas CYP3A catalyzes the formation of the minor (−)-9-hydroxymetabolite. Potential drug-drug interaction should be kept in mind when drugs influencing CYP2D6 and CYP3A4 activities are co-administered with risperidone. With regard to this, in a recent study in patients with schizophrenia or schizoaffective disorder, concomitant administration of paroxetine, a potent inhibitor of CYP2D6, was found to increase plasma levels of risperidone and the active moiety, leading to extrapyramidal side effects in one subject (Spina et al., 2001b). On the other hand, addition of carbamazepine, an inducer of CYP3A4, markedly decreased the plasma levels of risperidone and its 9-hydroxymetabolite, possibly resulting in clinical consequence (Spina et al., 2000, 2001c).
Acknowledgments
We thank Dr. Bo Eriksson (Jenssen-Cilag, Mölndal Sweden) for providing us with the 9-hydroxyrisperidone enantiomers and Prof. Leif Bertilsson for valuable critic.
Footnotes
-
This study has been supported by grants from Björn Lindström Memorial Found, the Swedish Medical Research Council (3902), and Karolinska Institutet.
- Abbreviations used are::
- HPLC
- high pressure liquid chromatography
- P-450 or CYP
- cytochrome P-450
- CLint
- intrinsic clearance
- Received April 12, 2001.
- Accepted June 4, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}