


Abstract
Tamoxifen (TAM), used as the endocrine therapy of choice for breast cancer, undergoes metabolism primarily forming N-desmethyltamoxifen, 4-hydroxytamoxifen, α-hydroxytamoxifen, and tamoxifen-N-oxide (TNO). Our earlier studies demonstrated that flavin-containing monooxygenases (FMOs) catalyze the formation of TNO. The current study demonstrates that human FMO1 and FMO3 catalyze TAM N-oxidation to TNO and that cytochromes P450 (P450s), but not FMOs, reduce TNO to TAM. CYP1A1, CYP1A2, CYP2A6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4 all reduced TNO, with CYP2A6, CYP1A1, and CYP3A4 producing the greatest reduction. A portion of TAM formed by CYP3A4-mediated reduction of TNO was further metabolized, but not TAM formed by the other P450s. TNO reduction by P450s is extremely rapid with considerable TAM formation detected at the earliest time point that products could be measured. TAM formation exhibited a lack of linearity with incubation time but increased linearly as a function of TNO and P450 concentration. TNO was converted into TAM by reduced hemoglobin (Hb) and NADPH-P450 oxidoreductase, suggesting involvement of the same heme-Fe2+ complex in both Hb and P450s. The findings raise the question of whether the reductive activity may be nonenzymatic. Results of this in vitro study demonstrate the potential of TAM and TNO to be interconverted metabolically. FMO seems to be the major enzymatic oxidant, whereas several P450 enzymes and even reduced hemoglobin are capable of reducing TNO back to TAM. The possibility that these processes may comprise a metabolic cycle in vivo is discussed in this article.
Tamoxifen (TAM), an antiestrogenic triphenylethyleneamine derivative, is currently used extensively for breast cancer therapy, and its use has been proposed as a prophylactic agent in healthy women who are considered to be at high risk for breast cancer (Jordan, 1995; Fischer et al., 1998). However, its potential prophylactic benefit is somewhat compromised by an infrequent incidence of TAM-mediated endometrial cancer (Fischer et al., 1998). The mechanism of the endometrial carcinogenic activity of TAM is not fully understood, although it has been studied by several investigators who proposed the formation of TAM-DNA adducts as the potential carcinogenic initiators. Indeed, Shibutani et al. (2000) demonstrated the formation of TAM-DNA adducts in women treated with TAM. DNA adduct formation seems to involve a reaction of “activated” TAM metabolites with DNA (Moorthy et al., 1996; Divi et al., 2001). TAM is metabolized by P450 enzymes, predominantly into N-desmethyltamoxifen, 4-hydroxytamoxifen, and α-hydroxytamoxifen (Reunitz et al., 1984; Mani et al., 1993a; Lim et al., 1994) and into tamoxifen-N-oxide (TNO; Foster et al., 1980; McCague and Seago, 1986) by the flavin-containing monooxygenases (FMOs; Mani et al., 1993b). It has been proposed that the α-hydroxylated TAM metabolites, which became O-sulfated, are the reactive TAM intermediates that form DNA-adducts (Dasaradhi and Shibutani, 1997; Divi et al., 2001). In addition, quinone methide metabolites of TAM have been suggested as the reactive compounds in DNA-adduct formation (Fan et al., 2000; Fan and Bolton, 2001). Despite these potential drawbacks, TAM is still considered to be the most useful triphenylethyleneamine (TPE) agent for the endocrine treatment of breast cancer, and its safety as a therapeutic drug for up to 5 years of treatment has been established. Nevertheless, relatively novel TPEs, such as droloxifene, toremifene, and idoxifene, that apparently do not form DNA adducts as avidly as TAM are being investigated as potential second generation endocrine therapeutic agents (Hasmann et al., 1994; Pace et al., 1997; Williams and Jeffrey, 1997).
Of additional interest is our earlier observation that a metabolite(s) of TNO binds covalently to hepatic microsomal proteins much less effectively than TAM metabolite(s) (Mani and Kupfer, 1991), suggesting that TNO, if used as a drug, may exhibit fewer undesirable side effects than TAM. That finding and the observations that TNO has similar antiestrogenic potency to TAM in MCF7 proliferation assay (Bates et al., 1982) and that TNO can be easily reduced back to TAM (this article) suggest that TNO could possibly be developed as an agent for treatment of breast cancer patients and/or for prophylactic use instead of TAM per se.
Our earlier studies have shown that mouse FMO catalyzes the transformation of TAM into TNO (Mani et al., 1993b). In the course of that study, it was observed that rat and human liver microsomes incubated in the presence of methimazole (an FMO inhibitor) effectively reduced TNO back to TAM. In the current study, we investigated the role of human FMO enzymes in the conversion of TAM into TNO and the function of human P450s and other heme-containing proteins in the reductive metabolism of TNO. Our findings that TNO is easily reduced to TAM by P450s and by hemoglobin (Hb) and possibly by other heme proteins suggest that TNO could serve as a prodrug for the generation of TAM.
Materials and Methods
Chemicals. NADPH, glucose 6-phosphate, glucose-6-phosphate dehydrogenase, methimazole, ketoconazole, troleandomycin, tranylcypromine, 1,2-didodecanoyl-rac-glycero-3-phosphocholine (dl-α-dilauroyl phosphatidylcholine), and tamoxifen were purchased from Sigma-Aldrich (St. Louis, MO). [N-methyl-3H]TAM ([3H]TAM) (85 Ci/mmol) was obtained from Amersham Biosciences UK, Ltd. (Little Chalfont, Buckinghamshire, UK). Sources of other reagents were as follows. EcoLume, the biodegradable scintillation fluid (for scintillation counting), was purchased from MP Biomedicals (Irvine, CA). Individual and pooled human liver microsomes, male or female rat liver microsomes, control supersomes (containing “empty” vectors in insect cell line), recombinant human NADPH-P450 oxidoreductase (OR), cDNA-expressed human P450 reductase and cytochrome b5, human FMO1 and FMO3 supersomes, and cDNA-expressed individual human cytochrome P450s supersomes (provided as microsomal preparations) were obtained from BD Gentest (Woburn, MA). Purified Escherichia coli expressed recombinant human P450s, purified recombinant human NADPH-P450 oxidoreductase, and purified human cytochrome b5 were obtained from PanVera Corp. (Madison, WI), and hemoglobin (Hb) was from Schwarz Mann (Orangeburg, NY). All other chemicals were of reagent grade quality.

N-Oxidation of tamoxifen by FMO1 and FMO3. [3H]Tamoxifen (100,000 dpm, 100 nmol) was incubated for 30 min with 25, 50, or 100 pmol of FMO1 (A) or with 50, 100, or 200 pmol of FMO3 (B) in the presence of NADPH without or with 0.2 and 1 mM MTM. Incubation volume was 1 ml. Values of TNO formed are Mean ± S.D. of triplicate determinations. Levels were considered significant at P ≤ 0.05. * indicates significance of 2a with respect to 1a, 2b with respect to 1b, and 2c with respect to 1c. ** indicates significance of 3a with respect to 2a, 3b with respect to 2b, and 3c with respect to 2c. ▴ indicates significance with respect to its FMO group lacking MTM. ▪ indicates significance compared with its FMO + 0.2 mM MTM group. □, FMO;  , FMO + 0.2 mM MTM; ▪, FMO + 1 mM MTM.
, FMO + 0.2 mM MTM; ▪, FMO + 1 mM MTM.
Incubation of [3H]TAM with cDNA-Expressed Human FMO1 and FMO3 Supersomes. Incubation mixtures (1 ml) contained 0.6 ml of sodium phosphate buffer, pH 8.6 (60 μmol); 1 mM NADPH, FMO1, or FMO3; and H2O with or without methimazole (MTM). After preincubation at 37°C for 2 min, reactions were initiated with [3H]TAM (100,000 dpm, 100 nmol) added in 10 μl of ethanol and incubated at 37°C for 30 or 60 min in a Dubnoff metabolic shaker bath under air. Controls lacked FMO. At the desired time point, 1 ml of ice-cold ethanol was added and tubes were vortexed and centrifuged to eliminate the precipitated proteins at 14,000 rpm for 10 min. The resulting pellet was washed successively with 1 ml of ethanol and 1 ml of methanol. The alcohol supernatants were combined and evaporated at room temperature under nitrogen (N2). The dry residue was dissolved in 1 ml of ethanol and centrifuged, the filtrate evaporated as above, and the residue dissolved in 100 μl of ethanol. Metabolites in 0.02 ml were separated by TLC using normal phase LK6DF Silica Gel 60Å. TLC plates containing a fluorescent indicator (Whatman, Clifton, NJ) were developed in CHCl3/methanol/NH4OH (80:20:0.5) and were quantified by radioscanning as previously described (Mani et al., 1993a,b). TAM and the corresponding TNO (see below) were chromatographed in adjacent lanes as standards. The plates were visualized under UV light (254 nm). The percentage of radioactivity in every UV-absorbing compound in each lane was determined with a System 200 Imaging Scanner (BioScan, Washington, DC), and the amount of each metabolite was quantified.
Synthesis of TNO. The [3H]TAM substrate (70-85 Ci/mmol; Amersham Biosciences Inc., Piscataway, NJ) was found to contain a contaminant on TLC chromatography. To purify the TAM, radiolabeled TAM was subjected to TLC and the silica gel zone containing TAM (detected by radioscanning) was scraped, eluted with 15 ml of methanol, centrifuged at 3000 rpm for 15 min at 4°C, washed again with 15 ml of ethanol, and recentrifuged as above. The combined alcoholic supernatant containing the pure TAM was evaporated. TNO was synthesized as described previously (Foster et al., 1980; Mani and Kupfer, 1991; Mani et al., 1993b). Purified radiolabeled TAM was mixed with nonradiolabeled TAM to a concentration of 200,000 dpm, 100 nmol per 10 μl of HPLC-grade methanol. H2O2 (30%; 0.5 ml/15 mg TAM) was added to 1 ml of this solution. After storage in the dark at room temperature for 24 h, the solution was evaporated to dryness under N2. The residual H2O2 was eliminated by adding small amounts of ethanol and by evaporating the solution to dryness (three times). The residue was dissolved in ethanol to obtain a concentration of 200,000 dpm (100 nmol/10 μl ethanol). After ascertaining by TLC that the purity of the TNO was close to 100%, the TNO was used or stored in ethanol under argon at 4°C until use.
Metabolism of TNO by Liver Microsomes or FMO Enzymes. Incubations were as described for TAM, except that a NADPH-regenerating system (10 μmol of glucose 6-phosphate, 0.5 μmol of NADPH, 2 IU of glucose-6-phosphate dehydrogenase, and 10 μmol of MgCl2) was used for the liver microsomes. Reaction mixtures (1 ml) containing 1 mg of microsomal protein, 50 pmol of FMO1, or 150 pmol of FMO3 were tested to study the metabolism of TNO. The reactions were initiated with [3H]TNO (200,000 dpm, 100 nmol) in 10 μl of ethanol and incubated (in 1 ml of final volume) at 37°C for 60 min, unless stated otherwise. Controls contained the same constituents but without the liver microsomes or FMO enzymes. The incubations were terminated as described for incubations of TAM. When aerobic and anaerobic incubations were compared, both contained glucose (5 mM); however, the anaerobic incubations were rendered hypoxic with a stream of argon before the addition of glucose oxidase (5 U) and catalase (30 U) to catalyze reactions that deplete the residual oxygen (Tschirret-Guth and Wood, 2003).
Incubation of [3H]Tamoxifen N-Oxide with cDNA-Expressed Human Cytochrome P450 Supersomes. Incubation mixtures (1 ml) contained supersomes for cDNA-expressed human cytochromes P450, CYP1A1, CYP1A2, CYP2A6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, or CYP3A4 (each at 100 pmol), and 2 mM NADPH in 60 mM sodium phosphate buffer, pH 7.4. After preincubation at 37°C for 2 min, reactions were initiated with [3H]TNO (200,000 dpm, 100 nmol), added in 10 μl of ethanol, and incubated at 37°C for 20 or 60 min in a Dubnoff metabolic shaker bath. Potential contributions toward this reductive process by NADPH, OR, and cytochrome b5 were determined using controls containing NADPH alone or controls with 100 pmol of cDNA-expressed OR supersomes or cDNA-expressed OR containing coexpressed cytochrome b5 supersomes (BD Gentest). In addition, a reconstituted system (devoid of supersomes) composed of lipid vesicles of 1,2-didodecanoyl-rac-glycero-3-phosphocholine (dl-α-dilauroyl phosphatidylcholine) containing various components was also used to investigate the potential contribution of OR and cytochrome b5 to the reduction of TNO, as previously described for purified P450s (Vatsis et al., 1982). In brief, 150 pmol of purified recombinant OR and 100 pmol of cytochrome b5 individually or in combination were added to 30 μl of dl-α-dilauroyl phosphatidylcholine (1 mg/ml H2O), sonicated to form lipid vesicles, and incubated with [3H]TNO (100,000 dpm, 100 nmol) in 0.8 ml of sodium phosphate buffer (60 mM), pH 7.4. After preincubation at 37°C for 2 min, reactions were started with 2 μmol of NADPH in 0.2 ml of sodium phosphate buffer (20 mM), pH 7.4, containing 10 μM MgCl2 in a final volume of 1 ml and incubated at 37°C. After 60 min, 1 ml of ethanol was added and the metabolites formed were isolated and quantified by TLC chromatography and radioscanning as described above.
Incubation of [3H]TNO with HLM or with cDNA-Expressed Human CYP1A1, CYP2A6, and CYP3A4 and Their Respective Inhibitors. HLM or cDNA-expressed human CYP1A1, CYP2A6, and CYP3A4 were incubated as described above in the presence or absence of their respective inhibitors [100 μM α-naphthoflavone for CYP1A1, 1 μM tranylcypromine for CYP2A6, and 1 μM ketoconazole or 100 μM troleandomycin (TAO) for CYP3A4]. For incubations with TAO, the enzyme preparation (CYP3A4) was preincubated with TAO in the presence of NADPH at 37°C for 10 min (to generate the inhibitory species) before adding the substrate.
Incubation of [3H]TNO with Hemoglobin. Reaction mixtures (0.5 ml) containing [3H]TNO (100,000 dpm, 50 nmol), Hb alone, or 75 pmol of purified recombinant OR and/or 2 mM NADPH were preincubated for 2 min at 37°C. The reaction was initiated with [3H]TNO (100,000 dpm, 50 nmol). One milliliter of ice-cold ethanol was added after the preincubation but before adding [3H]TNO for the zero time point or after incubation at 37°C for 30 min.
HPLC-Electrospray Ionization Mass Spectrometry of the Metabolites of Tamoxifen N-Oxide Reduction. The extract components were separated using a Rheos 2000 solvent delivery system (Flux Instruments AG, Basel, Switzerland). Five microliters of the extract diluted 1:20 with 10% ethanol were injected via a Rheodyne 8125 injector fitted with a 20-μl loop. TNO and its metabolites were separated on a C-18 Hypersil column (3-μm particle size, 100 × 1 mm i.d.) by elution with 1% ammonium acetate, pH 5.7, in a gradient of 50 to 100% methanol (in 7.5 min) and held at 100% methanol (for 12.5 min). The mobile phase was at a flow rate of 50 μl/min. The eluant leaving the HPLC column was directed into a Thermo Electron LCQ quadrupole ion-trap mass spectrometer (Thermo Electron Corporation, Waltham, MA) fitted with an electrospray ion source. Capillary and HV electrode potentials were 3 V and 4.5 kV, respectively. Data were acquired in the positive ion mode over the mass range of m/z 50 to 500.
Data Analysis. All of the data reported represent the mean ± S.D. of triplicate determinations, unless stated otherwise. All experiments were conducted at least twice. Values were compared with Student's unpaired t test, and the difference was considered significant if the p < 0.05.
Results
N-Oxidation of TAM by Human FMOs. cDNA-expressed FMO1 and FMO3 oxidized TAM into TNO (Fig. 1). In our previous study, human FMO5 was found to be inactive toward TAM oxidation (Hodgson et al., 2000) and hence was not further investigated.
TAM N-oxidation was significantly elevated by increasing concentrations of cDNA-expressed FMO1 at an incubation time of 30 min. Controls incubated in the absence of FMO1 did not generate TNO (data not shown). Incubations of TAM with 50 and 100 nM FMO1, along with 0.2 mM MTM, inhibited TNO formation by approximately 22%, and 1 mM MTM inhibited TNO formation by 52 and 53%, respectively (Fig. 1A). Increasing incubation time to 60 min increased TNO formation 1.5-fold over 25 nM FMO1 but did not alter the effect of 50 or 100 nM FMO1. The pattern and extent of inhibition in the presence of MTM remained essentially the same as that seen with 30-min incubation (data not shown).
FMO3 (Fig. 1B) produced less TAM N-oxidation than did FMO1. Nevertheless, as seen with FMO1, N-oxide formation progressively increased at FMO3 concentrations of 50, 100, and 200 nM. Whereas 0.2 mM MTM inhibited the catalysis by 60 to 90%, in 30-min incubations, MTM at 1 mM entirely inhibited the N-oxidation catalyzed by 200 nM FMO3. Incubation for 60 min did not enhance TNO formation further than was observed at 30-min incubation time (data not shown).
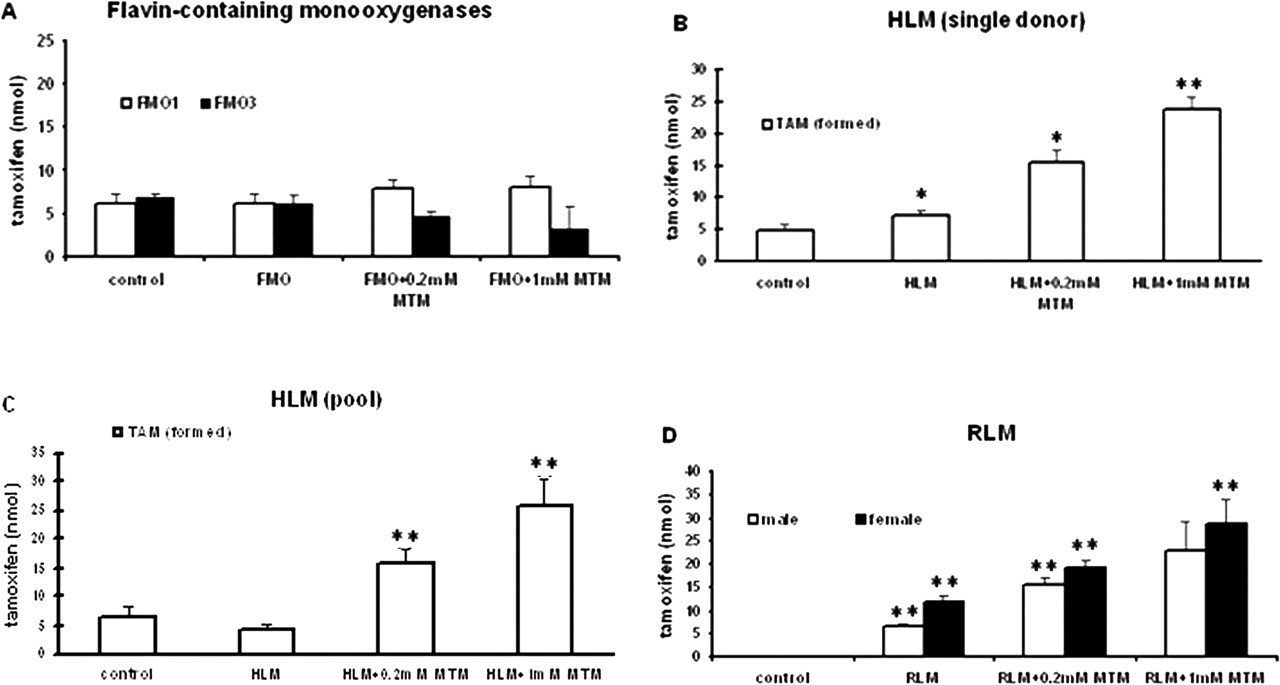
Incubation of TNO with FMO, HLM, and RLM (formation of TAM). [3H]TNO (200,000 dpm, 100 nmol) was incubated for 1 h with 50 pmol of FMO1 or 150 pmol of FMO3 (A), HLM (1-mg protein; single donor) (B), HLM (1-mg protein; pool) (C), male or female RLM (1-mg protein) (D) in the presence of NADPH regenerating system without or with 0.2 or 1 mM MTM for 1 h. Significance level set at P ≤ 0.05. Bars represent the amount of tamoxifen formed. Controls lacked the enzyme preparations (FMO or liver microsomes) but otherwise were similarly treated. * indicates significance with respect to control. ** indicates significance with respect to its preceding group.
Reduction of TNO to TAM. TNO was incubated with liver microsomes or with FMOs or P450 enzymes normally found in the hepatic endoplasmic reticulum. FMO1 and FMO3 did not catalyze the reduction of TNO back to TAM (Fig. 2A). However, HLM (from a single human donor or from pooled human livers) and RLM (from male or female rats) increased TAM formation from TNO compared with controls, indicating that enzymes other than FMOs present in these livers are involved in the reduction of TNO. To prevent the reoxidation of TAM back to TNO by FMOs in the liver microsomes, the FMO inhibitor MTM was included in the incubations. The amount of TAM was substantially increased in the presence of MTM (Fig. 2, B to D). Higher concentrations of MTM further increased TAM levels.
Heat-treated RLM (50°C for 90 s) produced a greater amount of TAM than that produced by RLM in the absence of heat, suggesting that heat inactivated the FMOs and thus prevented the regeneration of TNO from the newly formed TAM. However, MTM increased TAM accumulation even more than that seen with heat-treated RLM, indicating that it was a more effective inhibitor of FMO activities than heat inactivation (Fig. 3A). In addition, more TAM was formed from TNO by HLM in 60-min incubation than in 30-min incubation (Fig. 3B). It is noteworthy that depletion of oxygen from the incubation medium (depletion of oxygen is known to facilitate P450-reductive activity) (Webster et al., 1985) did not significantly alter TAM formation by HLM (Fig. 3C). Control incubations (lacking RLM or HLM) yielded no TAM.
Identification of P450s Reducing TNO in TAM. cDNA-expressed human CYP1A1, CYP1A2, CYP2A6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, and CYP3A4 were examined for their ability to reduce TNO. All of the P450s examined catalyzed the reduction of TNO, although noticeably higher TNO reduction was observed with CYP1A1, CYP2A6, and CYP3A4 (Fig. 4). In other experiments (data not shown), for all of the P450s, the use of TNO could be entirely accounted for by TAM. However, for CYP3A4, TNO plus residual TAM accounted for only 70 to 77% of the products, indicating that other products were produced. Figure 4 also shows that, by 20 min, CYP1A2, CYP2A6, CYP2C8, CYP2C9, CYP2C19, and CYP3A4 reduced TNO maximally, whereas CYP1A1, CYP2D6, and CYP2E1 continued to catalyze the reduction of TNO, even after a 60-min incubation.
NADPH alone (buffer without the P450 enzymes) did not reduce TNO to TAM, demonstrating that reducing equivalents alone were not sufficient to support that reduction (see control in Fig. 4). However, a small but significant amount of TAM was generated from TNO in incubations containing NADPH and supersomes (vector only) (data not shown) or by NADPH plus cDNA-expressed OR supersomes (without P450s) in the presence or absence of cytochrome b5, indicating some contribution to this process by the OR and possibly by cytochrome b5 (Fig. 4). Indeed, our experiments using purified recombinant OR and cytochrome b5 in a reconstituted system of lipid vesicles (described under Materials and Methods) suggest that supersomes probably contained some cytochrome b5, which catalyzed TNO reduction albeit to a small extent.
Reduction of TNO by Individual P450 Enzymes in the Presence of Specific P450 Inhibitors. In a related experiment, selective P450 inhibitors, such as α-naphthoflavone (CYP1A1 inhibitor), tranylcypromine (CYP2A6 inhibitor), and ketoconazole and TAO (CYP3A4 inhibitors), did not inhibit TNO reduction by their respective cDNA-expressed enzymes. By contrast, TAO and ketoconazole inhibited subsequent metabolism of the TAM formed by CYP3A4, indicating that these compounds are effective inhibitors only of the oxidative catalysis by the respective P450s (data not shown). This could also mean that different “active” sites of P450s (which are not amenable to the inhibitors), possibly in the vicinity of the heme or directly involving the heme, are involved in the TNO reduction. In turn, the likelihood of other heme-containing proteins (e.g., Hb) having the ability to reduce TNO was also considered (see below).

Incubation of [3H]TNO (200,000 dpm, 100 nmol) with 1 mg of RLM or HLM under various conditions (formation of TAM). Enzyme activity was studied under conditions of heat treatment of microsomes (50°C for 90 s) (A), incubation time (30 versus 60 min) (B), and anaerobiosis versus aerobiosis (C). Bars represent the amount of tamoxifen formed. Incubations in A and C were performed for 1 h. Respective controls were similarly treated but lacked the liver microsomes. P value of ≤ 0.05 was considered significant. * represents significance with respect to RLM, and ** represents significance with respect to preceding group in A. * represents significance with respect to 30-min incubation in B and with respect to aerobic conditions in C.
Incubation of [3H]TNO with Hb. A small amount of TAM was formed by incubation of Hb with TNO (100,000 dpm, 50 nmol) and NADPH, which was evident at the earliest time point that could be examined (designated here as zero incubation time), and did not increase at 30 min (Table 1, column III). Whereas reductase per se does not induce the reduction of TNO by Hb (Table 1, column II), Hb + NADPH + OR elicited a dramatic increase in the amount of TAM formed in 30 min (Table 1, column IV). As with the P450s, NADPH is absolutely essential for this reaction.
Reduction of TNO by Hb-forming tamoxifen Conditions are as described under Materials and Methods. The incubations contained 2 mM NADPH and [3H]TNO (100,000 dpm, 50 nmol) in a volume of 0.5 ml. Data represent mean values ± S.D. of TAM formed (in nanomolars).
Kinetics of TAM Formation from TNO by cDNA-Expressed CYP1A1, CYP2A6, and CYP3A4. TNO reduction by P450s did not increase as a linear function of incubation time. The finding that there was considerable TAM formation at the ostensible zero time points (no incubation) suggests that ethanol slowed rather than stopped the reaction. Nevertheless, reduction of TNO increased with increasing P450 concentration and increasing concentration of the substrate-TNO (data not shown). The amount of TAM formed was 11.5, 20.2, and 29.1 nmol for CYP2A6; 10.3, 11.7, and 20.8 nmol for CYP1A1; and 15.5, 19.9, and 23.6 nmol for CYP3A4 at enzyme concentrations of 25, 50, and 100 nM, respectively. For CYP3A4, 100 μM TAO was added to the incubations to block further metabolism of the TAM formed.
The observation that there were marked differences in the extent of reduction of TNO by the individual human P450s (i.e., CYP2A6, CYP1A1, and CYP3A4 as the most active) suggested that the P450 apoproteins contribute to the rate of catalysis; the heme seems to be mainly responsible for the reaction.
HPLC-Electrospray Ionization Mass Spectrometry Analysis. The product(s) generated by the P450 enzymes' reduction of TNO were resolved by liquid chromatography (LC) and identified using mass spectrometry. LC-MS analysis of the metabolites identified a compound with the molecular mass of TAM as the major metabolite. The retention times of TNO and TAM were 11 and 12.8 min, respectively. The MH+ ion from the product was observed at m/z 372.2, indicating a molecular weight of 371.2. This ion was also obtained with an authentic sample of TAM, thus confirming that the major metabolite was indeed TAM. The MH+ ion from authentic TNO was observed at 388.2, indicating a molecular weight of 387.2.
Discussion
This study demonstrates that both human FMO1 and FMO3 oxidize TAM into TNO and that FMO1 is more potent than FMO3 in catalyzing that reaction (Fig. 1). Moreover, FMO3 is substantially more sensitive to the inhibition of TAM oxidation by the FMO inhibitor MTM. Other distinguishing characteristics of FMO1 and FMO3 have been reported previously. For example, whereas FMO1 N-oxygenates only tertiary amines efficiently, human FMO3 N-oxygenates the primary, secondary, and tertiary amines (Cashman, 2000).
Earlier we had observed that TNO is metabolized in vitro to a much lesser extent than TAM (C. Mani and D. Kupfer, unpublished data), and we speculated then that TNO may function as a storage form for TAM in the body, yielding TAM as and when required. In the current study, we observed that incubations of TNO with rat or human liver microsomes supplemented with NADPH yield TAM as the major product, with TAM formation increasing further in the presence of the FMO inhibitor MTM (Fig. 2, B to D). Herein, we clearly show that the microsomal reduction of TNO to TAM is catalyzed not by FMOs but by P450 enzymes.
Moreover, TNO can efficiently be reduced back to TAM by a variety of heme-containing proteins. It is of interest in this context that the ability of hemoglobin to facilitate P450-like oxidations has been reported previously (Mieyal and Starke, 1994). Several aspects of the reduction indicate that it differs from a classic P450 monooxygenase reaction in that it 1) occurs extremely rapidly, 2) is catalyzed by numerous P450s without major selectivity, 3) is not significantly different in anaerobic versus aerobic conditions, and 4) is catalyzed by several heme-containing compounds, including hemoglobin and perhaps even cytochrome b5. These characteristics raise the possibility that the reductive reaction may be a chemical reaction involving heme, rather than a classic enzymic reaction. The evidence that reduction reaction rate for P450s CYP1A1, CYP2A6, and CYP3A4 was not inhibited by their respective inhibitors supports that idea. Thus, the mechanism of the reduction reaction will require further study. The main point in this article is that the reduction of TNO represents a novel type of reaction for P450.

Reduction of tamoxifen N-oxide by P450s and NADPH oxidoreductase (formation of TAM). [3H]TNO (200,000 dpm, 100 nmol) was incubated in the presence of only NADPH (control), NADPH + OR without or with coexpressed cytochrome b5, or 100 pmol of cDNA expressed human cytochrome P450 enzymes for 20 min (empty bar) or 60 min (filled bar) in a volume of 1 ml. Each bar represents the average value of TAM formed from two incubations.
This investigation suggests that TAM may undergo oxidation and TNO reduction in the liver in a cyclic fashion. Thus, hepatic FMO1 and FMO3 oxidize TAM to TNO and, in turn, a certain portion of the TNO formed is reduced back to TAM by P450s. In addition, results with Hb in vitro suggest that TNO could possibly be reduced while circulating in body fluids by appropriate heme proteins; e.g., by Hb in red blood cells or even by muscle myoglobin.
TNO has been detected in the plasma of breast cancer patients undergoing TAM therapy (Poon et al., 1993). However, there have not been any studies published to our knowledge that examine whether some of the TNO in vivo can be reduced back to TAM in either animals or humans. Of interest is a related study demonstrating a reduction of another amine N-oxide, the endogenous trimethylamine N-oxide in humans using a pharmacogenetic approach (Al-Waiz et al., 1987). In that study involving individuals with an inherited deficiency of FMO3, approximately 50% of orally administered trimethylamine N-oxide was excreted as trimethylamine, indicating the presence of a functional reductive metabolism of N-oxides in humans; however, the nature of the catalysts and the mechanism involved in the in vivo reduction of trimethylamine N-oxide have not been identified. Nevertheless, based on these observations and our in vitro findings, it can be speculated that the reduction of TNO to TAM in humans in vivo would be a likely occurrence.
There is a tertiary amine N-oxide reduction system in rat liver (mitochondria, microsomes, and cytosol) composed of quinone reductase/diaphorase and heme (Kitamura et al., 1999). That reduction system was studied primarily with imipramine N-oxide as a substrate; however, whether that system could also reduce TNO has not been evaluated.
P450s are involved in the reduction of certain classes of organic compounds. For instance, human CYP3A enzyme has been implicated in the reduction of the anticancer prodrug anthraquinone di-N-oxide, a tertiary amine-N-oxide (Raleigh et al., 1998). Our current study demonstrates the participation of human P450 enzymes, among these CYP3A4 in the reduction of the N-oxide of TAM. These findings and the observations that TNO forms far less DNA and protein adducts than does TAM (Dehal and Kupfer, 1999; Umemoto et al., 2000) suggest that TNO could be an attractive candidate for a prodrug of TAM. Other tamoxifen analogs undergoing clinical trials for antibreast cancer treatment (e.g., iodoxifene, toremifene, and droloxifene) have also been shown to form the corresponding N-oxides (McCague et al., 1990; John et al., 2002; Jones and Lim, 2002). Given the structural similarities between these TPE compounds and TAM, it seems reasonable that the N-oxides of these selective estrogen receptor modulator analogs and of related TPEs could also be reduced to their respective tertiary amines.
The concept of using bioreductively activated drugs in the treatment of solid tumors has been gaining popularity. Tumors that are hypoxic due to malformed vasculature are thought to be indicators of more aggressive disease and enhanced levels of metastasis, because they are resistant to radiotherapy and chemotherapy (Brizel et al., 1996; Hockel et al., 1996). Because TNO is reduced to TAM in the presence or absence of oxygen, it can be expected that TNO will be reduced in both oxygenated and hypoxic tumors. Biotransformation of TNO by P450 forms expressed extrahepatically, such as in breast and endometrium, may be important in determining the tissue-specific effects of TNO. For instance, normal breast tissue and breast tumors were found to contain mRNAs and proteins of certain P450s (Huang et al., 1996; Warner et al., 1997). Because all P450s examined by us as well as Hb were active in TNO reduction, it is highly likely that P450s and Hb in tumor or breast tissue will reduce TNO to TAM. As a result, to enhance transformation of TNO to TAM, vectors coexpressing P450s such as CYP2A6, CYP1A1, or CYP3A4 could be injected directly into breast tumors and thereby trigger these cells to overexpress the respective enzyme. In turn, this procedure would probably increase the efficacy of TNO when administered as a prodrug. Such an approach has been successful in the past with cyclophosphamide treatment of tumors (Waxman et al., 1999). However, further studies are needed to explore the effectiveness of TNO as an antibreast cancer agent and the question of its potential toxicology in vivo needs to be ruled out before TNO can be aimed for clinical investigation.
Acknowledgments
We thank Dr. James Evans (University of Massachusetts Medical School, Worcester, MA) for performing the LC-MS analysis and for help in analyzing the products of TNO reduction, Dr. Eszter Hazai for assistance of with some of our studies, and Elizabeth Euloth for secretarial assistance.
Footnotes
-
This study was supported by National Institute of Environmental Health Sciences (NIEHS), National Institutes of Health Grant ES00834. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIEHS.
-
Preliminary account of this investigation was presented at the Experimental Biology 2004 Meeting in Washington DC, 17-21 April, 2004 (Abstract 3429).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.104.000802.
-
ABBREVIATIONS: TAM, tamoxifen; P450, cytochrome P450; FMO, flavin-containing monooxygenase; Hb, hemoglobin; HLM, human liver microsome(s); MTM, methimazole; HPLC, high-pressure liquid chromatography; OR, NADPH-P450 oxidoreductase (P450 reductase); RLM, rat liver microsome(s); [3H]TAM, [N-methyl-3H]TAM; TAO, troleandomycin; TNO, tamoxifen N-oxide; TPE, triphenylethyleneamine; LC, liquid chromatography; MS, mass spectrometry; TLC, thin layer chromatography.
-
↵† Deceased December 10, 2004.
- Received June 3, 2004.
- Accepted June 28, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}