


Abstract
Nornicotine is an N-demethylated metabolite of nicotine. In the present study, human cytochrome P450 (P450) isoform(s) involved in nicotine N-demethylation were identified. The Eadie-Hofstee plot of nicotine N-demethylation in human liver microsomes was biphasic with high-affinity (apparent Km = 173 ± 70 μM, Vmax = 57 ± 17 pmol/min/mg) and low-affinity (apparent Km = 619 ± 68 μM, Vmax = 137 ± 6 pmol/min/mg) components. Among 13 recombinant human P450s expressed in baculovirus-infected insect cells (Supersomes), CYP2B6 exhibited the highest nicotine N-demethylase activity, followed by CYP2A6. The apparent Km values of CYP2A6 (49 ± 12 μM) and CYP2B6 (550 ± 46 μM) were close to those of high- and low-affinity components in human liver microsomes, respectively. The intrinsic clearances of CYP2A6 and CYP2B6 Supersomes were 5.1 and 12.5 nl/min/pmol P450, respectively. In addition, the intrinsic clearance of CYP2A13 expressed in Escherichia coli (44.9 nl/min/pmol P450) was higher than that of CYP2A6 expressed in E. coli (2.6 nl/min/pmol P450). Since CYP2A13 is hardly expressed in human livers, the contribution of CYP2A13 to the nicotine N-demethylation in human liver microsomes would be negligible. The nicotine N-demethylase activity in microsomes from 15 human livers at 20 μM nicotine was significantly correlated with the CYP2A6 contents (r = 0.578, p < 0.05), coumarin 7-hydroxylase activity (r = 0.802, p < 0.001), and S-mephenytoin N-demethylase activity (r = 0.694, p < 0.005). The nicotine N-demethylase activity at 100 μM nicotine was significantly correlated with the CYP2B6 contents (r = 0.677, p < 0.05) and S-mephenytoin N-demethylase activities (r = 0.740, p < 0.005). These results as well as the inhibition analyses suggested that CYP2A6 and CYP2B6 would significantly contribute to the nicotine N-demethylation at low and high substrate concentrations, respectively. The contributions of CYP2A6 and CYP2B6 would be dependent on the expression levels of these isoforms in any human liver.
Over a billion people worldwide smoke tobacco. Smoking exerts complex central and peripheral nervous system, behavioral, cardiovascular, and endocrine effects in humans (Vial 1986; Benowitz, 1988). The addiction liability and pharmacological effects of smoking are due to nicotine. Pulmonary absorption of nicotine is extremely rapid, occurring at a rate similar to that after intravenous administration (Russell and Feyerabend, 1978). In humans, absorbed nicotine is rapidly and extensively metabolized and eliminated to urine (Benowitz, 1988).
Nicotine is mainly (70–80%) metabolized to cotinine (Fig. 1) by cytochrome P450 (P450) 2A6 in humans (Nakajima et al., 1996). Nicotine is also metabolized to nornicotine, via N-demethylation. In humans, 2 to 3% of nicotine is excreted as nornicotine in 24-h urine (Kyerematen et al., 1990; Yamanaka et al., 2004). Cundy and Crooks (1984) have reported that only small amounts of nornicotine (1.6%) were detected in 24-h urine after [14C]nicotine was injected intraperitoneally into guinea pigs. After a single arterial dose of labeled nicotine in rats, it was found that nornicotine accounted for 8% of the total recovery of the administered dose in urine (Curvall and Kazemi Vala, 1993). Therefore, the metabolism of nicotine to nornicotine would be a relatively minor pathway in the systemic clearance of nicotine. However, experimental animal studies revealed that nornicotine is present in brain at significant levels (approximately 20% of the total amount of nicotine and its metabolites) (Nordberg et al., 1989; Plowchalk et al., 1992; Nyback et al., 1994; Crooks et al., 1997). The nornicotine concentration in brain was nearly equal to that of nicotine (Crooks et al., 1997). These data indicated that nornicotine is a major metabolite of nicotine in brain. The half-lives of nornicotine in human plasma and rat brain are 6-fold and 3-fold longer than those of nicotine, respectively (Kyerematen et al., 1990; Ghosheh et al., 1999). Since nornicotine is pharmacologically active (Zhang et al., 1990; Liu et al., 1993), it may contribute to the neuropharmacological effects of nicotine and tobacco usage.
In our recent study (Yamanaka et al., 2004), the urinary excretion profile of nicotine metabolites in subjects entirely lacking the CYP2A6 gene (poor metabolizers) was determined. We found that the urinary excretion levels of nornicotine were similar between the poor metabolizers and extensive metabolizers who are homozygotes of the wild-type CYP2A6 allele (Yamanaka et al., 2004), indicating the possibility that the enzyme responsible for the nornicotine formation might not be CYP2A6. In contrast, Murphy et al. (2005) recently reported that human CYP2A6 has a catalytic activity for nicotine N-demethylation. However, they did not investigate the other P450 isoforms. In the present study, we sought to identify the P450 isoform(s) involved in the nicotine N-demethylation to nornicotine in human liver microsomes.
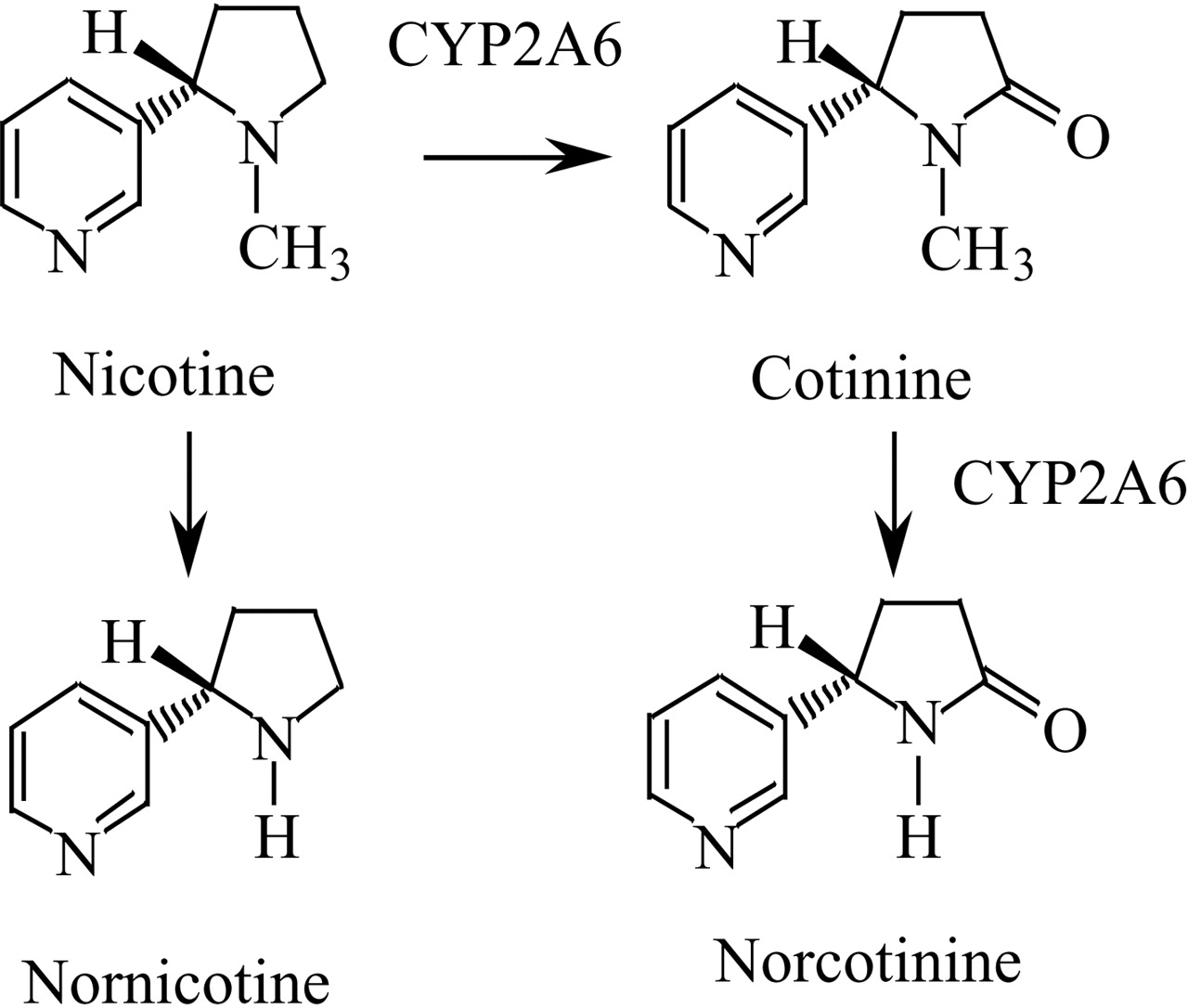
Metabolic pathway of nicotine to cotinine, nornicotine, and norcotinine in humans.
Materials and Methods
Materials. Nicotine, nornicotine, and orphenadrine were purchased from Sigma-Aldrich (St. Louis, MO). Coumarin, quinidine, S-mephenytoin, and phenobarbital were purchased from Wako Pure Chemicals (Osaka, Japan). Nirvanol and ketoconazole were obtained from Ultrafine (Manchester, UK) and BIOMOL Research Laboratories (Plymouth Meeting, PA), respectively. NADP+, glucose 6-phosphate, and glucose-6-phosphate dehydrogenase were purchased from Oriental Yeast Co. (Tokyo, Japan). Pooled human liver microsomes (H161), microsomes from 15 individual human livers (H003, H006, H023, H030, H043, H056, H064, H066, H070, H089, H093, H095, H112, HK23, and HK34), and recombinant human CYP1A1, CYP1A2, CYP1B1, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, CYP3A5, and CYP3A7 expressed in baculovirus-infected insect cells (Supersomes) were purchased from BD Gentest (Woburn, MA). All recombinant P450s were coexpressed with NADPH-P450 oxidoreductase. CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2E1, CYP3A4, and CYP3A7 were also coexpressed with cytochrome b5. CYP1A2, CYP2A6, CYP2B6, CYP2D6, CYP2E1, and CYP3A4 protein contents in the microsomes of each human liver determined by immunoblot analyses were supplied by the manufacturer. The enzymatic activities for human P450 isoforms in each human liver microsome, including phenacetin O-deethylation (CYP1A2), coumarin 7-hydroxylation (CYP2A6), diclofenac 4′-hydroxylation (CYP2C9), S-mephenytoin 4′-hydroxylation (CYP2C19), bufuralol 1′-hydroxylation (CYP2D6), chlorzoxazone 6-hydroxylation (CYP2E1), and testosterone 6β-hydroxylation (CYP3A4), were also supplied by the manufacturer. Anti-human CYP2A6 monoclonal antibody, anti-human CYP2B6 monoclonal antibody, rabbit anti-human CYP2D6 serum, and rabbit anti-human CYP3A4 serum were obtained from Daiichi Pure Chemicals (Tokyo, Japan). It was reported by the manufacturer that these antibodies specifically react with each P450 isoform. All other chemicals and solvents were of the highest grade commercially available.
Construction of Expression Systems of CYP2A6 and CYP2A13 in Escherichia coli. A bicistronic construct consisting of the coding sequence of CYP2A6 followed by that of NADPH-cytochrome P450 reductase (NPR) was previously constructed in the pCW expression vector (Fukami et al., 2004). The full-length human CYP2A13 cDNA was obtained by polymerase chain reaction based on the reference sequence (GenBank accession number NM000766). A bicistronic construct consisting of the coding sequence of CYP2A13 followed by that of NPR was constructed using the CYP2A6/NPR expression vector. The 5′-terminus of CYP2A13 cDNA was modified to achieve a high expression level in a manner similar to that of the CYP2A6/NPR expression vector. Nucleotide sequences were confirmed by DNA sequence analysis (Long-Read Tower DNA sequencer; GE Healthcare, Little Chalfont, Buckinghamshire, UK). These plasmids were transformed to E. coli JM109. E. coli membranes expressing CYP2A6/NPR or CYP2A13/NPR were prepared, and the P450 content, protein concentration, and NADPH-cytochrome c reduction activity were determined according to the methods described previously (Fukami et al., 2004). P450s require a 1- to 3-fold excess amount of NPR to show the full activity in a reconstituted system (Soucek, 1999). The molar ratios of the NPR to the CYP2A6 and CYP2A13 were confirmed to be over 4 in the membrane preparations in the present study.
Preparation of Human Brain Microsomes from Striatum. This study was approved by the Ethics Committees of Iwate Medical University School of Medicine (Iwate, Japan) and Kanazawa University (Kanazawa, Japan). Human striatum samples (n = 3; 23, 79, and 82 years old) were obtained from autopsy materials that were discarded after pathological investigations. The postmortem delay was less than 24 h. After dissection, the samples were immediately frozen in liquid nitrogen and stored at –80°C. Microsomes were prepared as described by Voirol et al. (2000) with slight modifications, and were stored at –80°C until use.
Nicotine N-Demethylation Assays. The nicotine N-demethylase activity was determined by LC-MS/MS. A typical incubation mixture (200-μl total volume) contained 0.5 mg/ml human liver microsomes (or 20 pmol/ml Supersomes, 50 pmol/ml E. coli membrane, or 1 mg/ml human striatal microsomes), 50 mM potassium phosphate buffer (pH 7.4), an NADPH-generating system (0.5 mM NADP+, 2 mM glucose 6-phosphate, 1 U/ml glucose-6-phosphate dehydrogenase, 4 mM MgCl2), and nicotine (20 and 100 μM for human liver microsomes; 200 μM for recombinant P450s and human striatal microsomes). After a 2-min preincubation, the reactions were initiated by the addition of the NADPH-generating system and were incubated at 37°C for 20 min. The reactions were terminated by 100 μl of methanol. After centrifugation at 9000g for 5 min, the supernatant was filtered with a 0.22-μm filter (Ultrafree-MC centrifugal filter unit; Millipore Corporation, Eschborn, Germany). Aliquots of 5 μl were injected into the LC-MS/MS system.
LC-MS/MS Analysis. Nicotine and nornicotine were determined by LC-MS/MS as described previously (Yamanaka et al., 2004), with minor modifications. Liquid chromatography was performed using an HP 1100 system including a binary pump, an automatic sampler, and a column oven (Agilent Technologies, Waldbronn, Germany), which was equipped with a Develosil ODS-UG-3 (2.0 × 150 mm; Nomura Chemical, Aichi, Japan) column. The column temperature was 35°C. The mobile phase was 0.01% ammonia (A) and methanol (B). The conditions for elution were as follows: 20% B (0–0.5 min), 20 to 60% B (0.5–6 min), 60% B (6–10 min), 60 to 20% B (10–15 min). Linear gradients were used for all solvent changes. The flow rate was 0.2 ml/min. The liquid chromatograph was connected to a PE Sciex API 2000 tandem mass spectrometer (Applied Biosystems, Langen, Germany) operated in the positive electrospray ionization mode. The turbo gas was maintained at 550°C. Nitrogen was used as the nebulizing gas, turbo gas, and curtain gas at 50, 70, and 30 psi, respectively. Parent and/or fragment ions were filtered in the first quadrupole and dissociated in the collision cell using nitrogen as the collision gas. The collision energy ranged from 27 to 29 V. Two mass/charge (m/z) ion transitions were recorded in the multiple reaction monitoring mode: m/z 163 and 130 for nicotine, and m/z 149 and 80 for nornicotine. The retention times of nicotine and nornicotine were 13.0 min and 10.9 min, respectively (Fig. 2). Since nornicotine contaminants exist in the commercially available nicotine to the extent of ∼0.25%, the content of nornicotine in the mixture incubated without the NADPH-generating system was subtracted to correct the activity.
Kinetic Analyses of Nicotine N-Demethylase Activities in Human Liver Microsomes or Recombinant P450 Isoforms. The kinetic studies were performed using pooled human liver microsomes or recombinant P450s. When determining the kinetic parameters, the substrate concentration ranged from 10 μM to 2000 μM for human liver microsomes, and from 10 μM to 1000 μM for recombinant P450s. The kinetic parameters were estimated from the fitted curves using a KaleidaGraph computer program (Abelbeck/Synergy, Reading, PA) designed for nonlinear regression analysis.
Inhibition Study. The nicotine N-demethylase activity in pooled human liver microsomes at 100 μM nicotine was determined in the presence of inhibitors for CYP2A6, CYP2B6, CYP2D6, or CYP3A4. Coumarin (100 μM), orphenadrine (500 μM), quinidine (10 μM), and ketoconazole (1 μM) were used as specific inhibitors for CYP2A6 (Yun et al., 1991), CYP2B6 (Reidy et al., 1989), CYP2D6 (Broly et al., 1989), and CYP3A4 (Newton et al., 1995), respectively. All inhibitors were dissolved in methanol so that the final concentration of solvent in the incubation mixture was <1%. For the inhibition study with coumarin, quinidine, and ketoconazole, the assays were performed as described above. For the inhibition study with orphenadrine (mechanism-based inhibitor), the incubation mixture including the inhibitor was preincubated in the presence of the NADPH-generating system at 37°C for 15 min and the reaction was initiated by the addition of nicotine. For the immunoinhibition study, the human liver microsomes (0.5 mg/ml) were incubated with anti-human CYP2A6 antibody (50 μl/mg), anti-human CYP2B6 antibody (50 μl/mg), anti-human CYP2D6 serum (50 μl/mg), or anti-human CYP3A4 serum (50 μl/mg) in 0.1 M potassium phosphate buffer (pH 7.4) on ice for 1 h. The nicotine N-demethylase activities were determined as described above. In addition, two samples of human liver microsomes, H003 (CYP2A6-abundant) and H112 (CYP2B6-abundant), were also used for the inhibition study of CYP2A6 and CYP2B6.

Representative chromatograms of LC-MS/MS analysis of the nicotine N-demethylation in human liver microsomes. A and B, the incubation mixture including human liver microsomes and nicotine in the presence (A) or absence (B) of the NADPH-generating system. C, a mixture of authentic standards of nicotine and norcotinine (each 250 pg). Two mass/charge (m/z) ion transitions were recorded in the multiple reaction monitoring mode: m/z 163 and 130 for nicotine (13.0 min), and m/z 149 and 80 for nornicotine (10.9 min).
Prediction of the Percentage Contributions of CYP2A6 and CYP2B6 to Nicotine N-Demethylase Activity in Human Liver Microsomes. The percentage contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylase activity (20 μM nicotine) were estimated based on the contents of these P450 isoforms in human liver microsomes determined by immunoblotting, based on the following equations (Becquemont et al., 1998): VCYP2A6 = Vrec-CYP2A6 · A and VCYP2B6 = Vrec-CYP2B6 · B. A and B are the immunochemically determined CYP2A6 and CYP2B6 contents, respectively, in human liver microsomes. The Vrec-CYP2A6 and Vrec-CYP2B6 are the nicotine N-demethylase activities (20 μM nicotine) in recombinant CYP2A6 and CYP2B6, respectively. The contributions of CYP2A6 and CYP2B6 to the activity by human liver microsomes (VHL) were calculated as follows: contribution of CYP2A6 (%) = (VCYP2A6/VHL) × 100 and contribution of CYP2B6 (%) = (VCYP2B6/VHL) × 100.
S-Mephenytoin N-Demethylation Assay.S-Mephenytoin N-demethylase activity was determined by HPLC as described elsewhere (Ko et al., 1998) with minor modifications. A typical incubation mixture (200-μl total volume) contained 0.5 mg/ml human liver microsomal protein, 50 mM potassium phosphate buffer (pH 7.4), an NADPH-generating system, and 1 mM S-mephenytoin. After a 2-min preincubation, the reactions were initiated by the addition of the NADPH-generating system and were incubated at 37°C for 60 min. The reactions were terminated by 100 μl of ice-cold CH3CN and added phenobarbital (25 ng) as an internal standard. The reaction mixtures were centrifuged at 9000g for 5 min, and aliquots of 100 μl were injected into the HPLC system.
HPLC analyses were performed using a PC-980 pump (Jasco, Tokyo, Japan), a UV-970 intelligent UV-visible detector (Jasco), an AS-950-10 autosampler (Jasco), a D-2500 integrator (Hitachi, Tokyo, Japan), and a CTO-6A column oven (Shimadzu, Kyoto, Japan) equipped with a Capcell PAK C18 UG120 (4.6 × 250 mm; 5 μm) column (Shiseido, Tokyo, Japan). The eluent was monitored at 204 nm. The mobile phase was 25% CH3CN/50 mM potassium phosphate buffer (pH 3.8). The flow rate was 1.0 ml/min and the column temperature was 35°C. The retention times of nirvanol, phenobarbital, and S-mephenytoin were 9.5 min, 11.4 min, and 16.7 min, respectively. The quantification of nirvanol was performed by comparing the HPLC peak heights to those of an authentic standard with reference to an internal standard.
Statistical Analysis. Data are the mean of duplicate measurements. Correlations between the nicotine N-demethylase activity and immunoreactive P450 contents or enzymatic activities in microsomes from 15 human livers were determined by Pearson's product-moment method.
Results
Nicotine N-Demethylase Activity in Human Liver Microsomes. The formation of nornicotine from nicotine in the pooled human liver microsomes increased in protein concentration- and time-dependent manners. The formation was linear at least at 1 mg/ml microsomal protein and 30-min incubation. Unless specified, the standard incubation mixture containing 0.5 mg/ml microsomal protein was incubated at 37°C for 20 min. The kinetics of nicotine N-demethylation in the pooled human liver microsomes was biphasic (Fig. 3), indicating the involvement of multiple enzymes. The apparent Km and Vmax values for the high-affinity component were 173 ± 70 μM and 57 ± 17 pmol/min/mg, respectively. For the low-affinity component, the Km was 619 ± 68 μM and the Vmax was 137 ± 6 pmol/min/mg.
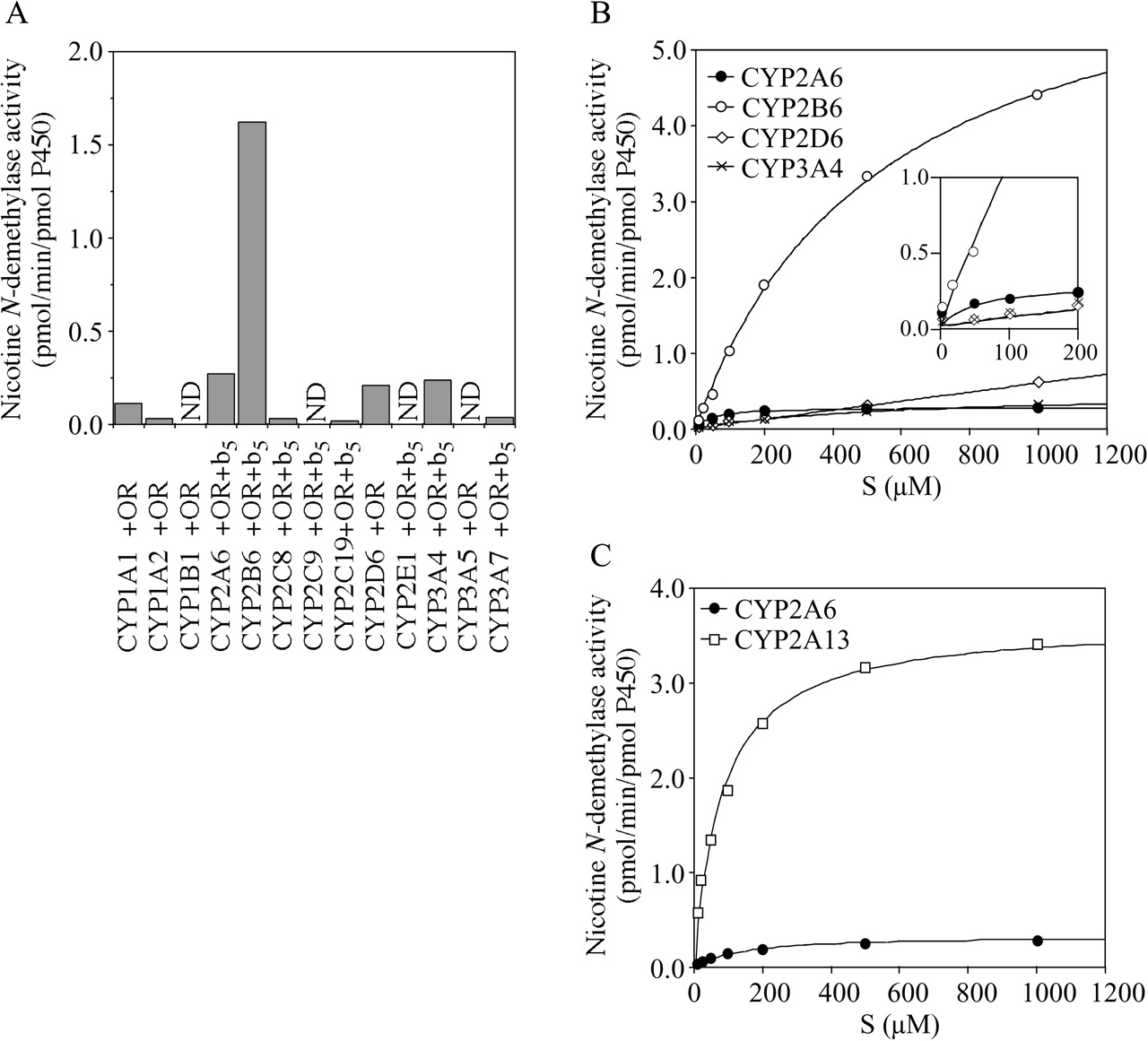
Nicotine N-Demethylase Activities by Recombinant P450 Isoforms. The nicotine N-demethylation was investigated in microsomes of baculovirus-infected insect cells expressing human P450s. As shown in Fig. 4A, CYP2B6 exhibited the highest nicotine N-demethylase activity (1.6 pmol/min/pmol P450), followed by CYP2A6 (0.3 pmol/min/pmol P450), CYP3A4 (0.2 pmol/min/pmol P450), and CYP2D6 (0.2 pmol/min/pmol P450). CYP1A1, CYP1A2, CYP2C8, CYP2C19, and CYP3A7 showed a trivial activity (0.02–0.11 pmol/min/pmol P450). Kinetic analyses were performed for the CYP2A6, CYP2B6, CYP2D6, and CYP3A4 Supersomes (Fig. 4B). The kinetics in recombinant CYP2B6 was fitted to the Michaelis-Menten plot, with apparent Km = 550 ± 46 μM and Vmax = 6.9 ± 0.3 pmol/min/pmol P450, resulting in the intrinsic clearance (CLint) of 12.5 nl/min/pmol P450. The kinetics in recombinant CYP2A6 was also fitted to the Michaelis-Menten plot, with an apparent Km = 49 ± 12 μM and Vmax = 0.3 ± 0.0 pmol/min/pmol P450, resulting in CLint of 5.1 nl/min/pmol P450. The activities in recombinant CYP2D6 and CYP3A4 were increased, with an increase in the concentration up to 1000 μM (0.6 and 0.3 pmol/min/pmol P450, respectively). The CLint values calculated with the initial slope of the plots of V (velocity) versus S (substrate concentration) in recombinant CYP2D6 and CYP3A4 were both 0.7 nl/min/pmol P450, respectively.

Kinetic analyses of the nicotine N-demethylation in human liver microsomes. Michaelis-Menten (A) and Eadie-Hofstee plots (B) are shown. Pooled human liver microsomes were incubated with 10 to 2000 μM nicotine at 37°C for 20 min. Each data point represents the mean of duplicate determinations.

Nicotine N-demethylase activities by recombinant human P450 isoforms. A, Nicotine N-demethylase activities by recombinant P450 isoforms expressed in baculovirus-infected insect cells (Supersomes) at 200 μM nicotine. Each column represents the mean of duplicate determinations. B, kinetic analyses of the nicotine N-demethylation by CYP2A6, CYP2B6, CYP2D6, and CYP3A4 Supersomes. C, kinetic analyses of the nicotine N-demethylation by recombinant CYP2A6 and CYP2A13 expressed in E. coli. The nicotine concentration ranged from 10 μM to 1000 μM. Each data point represents the mean of duplicate determinations. ND, not determined.
Furthermore, kinetic analyses were performed for the recombinant CYP2A6 and CYP2A13 expressed in E. coli (Fig. 4C). The kinetics in recombinant CYP2A6 and CYP2A13 were also fitted to the Michaelis-Menten plot. The apparent Km and Vmax values were 129 ± 26 μM and 0.3 ± 0.0 pmol/min/pmol P450 for the recombinant CYP2A6, resulting in a CLint of 2.6 nl/min/pmol P450. The apparent Km and Vmax values were 80 ± 11 μM and 3.7 ± 0.1 pmol/min/pmol P450 for the recombinant CYP2A13, resulting in a CLint of 44.9 nl/min/pmol P450. Although CYP2A13 exhibited the highest CLint of the nicotine N-demethylation, further analyses were not performed since this P450 isoform is hardly expressed in human liver microsomes.
Interindividual Variability in Nicotine N-Demethylase Activity in Microsomes from 15 Human Livers and Correlation Analyses. The nicotine N-demethylase activities in microsomes from 15 human livers were determined at substrate concentrations of 20 μM and 100 μM. The activities at 20 μM nicotine ranged from 0.6 to 9.7 pmol/min/mg, representing 16-fold variability. The activities at 100 μM nicotine ranged from 2.5 to 14.0 pmol/min/mg (6-fold variability). A significant correlation (r = 0.654, p < 0.005) was observed between these activities at two different substrate concentrations (Fig. 5). As summarized in Table 1, the nicotine N-demethylase activity at 20 μM nicotine was significantly correlated with the CYP2A6 contents (r = 0.578, p < 0.05) and coumarin 7-hydroxylase activity (r = 0.802, p < 0.001) as the specific activity for CYP2A6. In addition, it was significantly correlated with the S-mephenytoin N-demethylase activity (r = 0.694, p < 0.005) as the specific activity for CYP2B6. The nicotine N-demethylase activity at 100 μM nicotine was significantly correlated with the CYP2B6 contents (r = 0.677, p < 0.05) and S-mephenytoin N-demethylase activities (r = 0.740, p < 0.005) as well as testosterone 6β-hydroxylase activity (r = 0.523, p < 0.05) as the specific activity for CYP3A4. No significant correlations were observed between the nicotine N-demethylase activities and the other activities or P450 isoform contents.
Correlation coefficients (r) between the nicotine N-demethylase activity and P450 contents or specific activities in 15 human liver microsomes

Correlation analysis of the nicotine N-demethylase activities at two different substrate concentrations. The nicotine N-demethylase activities in microsomes from 15 human livers at 20 μM and 100 μM nicotine concentration were determined. Each data point represents the mean of duplicate determinations.
Inhibition Analyses. The effects of chemical inhibitors or antibodies against the P450 isoforms on the nicotine N-demethylase activities in pooled human liver microsomes were determined at 100 μM nicotine (Fig. 6). The activity was inhibited by coumarin (67% of the control activity), anti-CYP2A6 antibody (40% of the control activity), orphenadrine (28% of the control activity), and anti-CYP2B6 antibody (30% of the control activity). In contrast, the activity was not affected by quinidine, anti-CYP2D6 serum, ketoconazole, or anti-CYP3A4 serum.
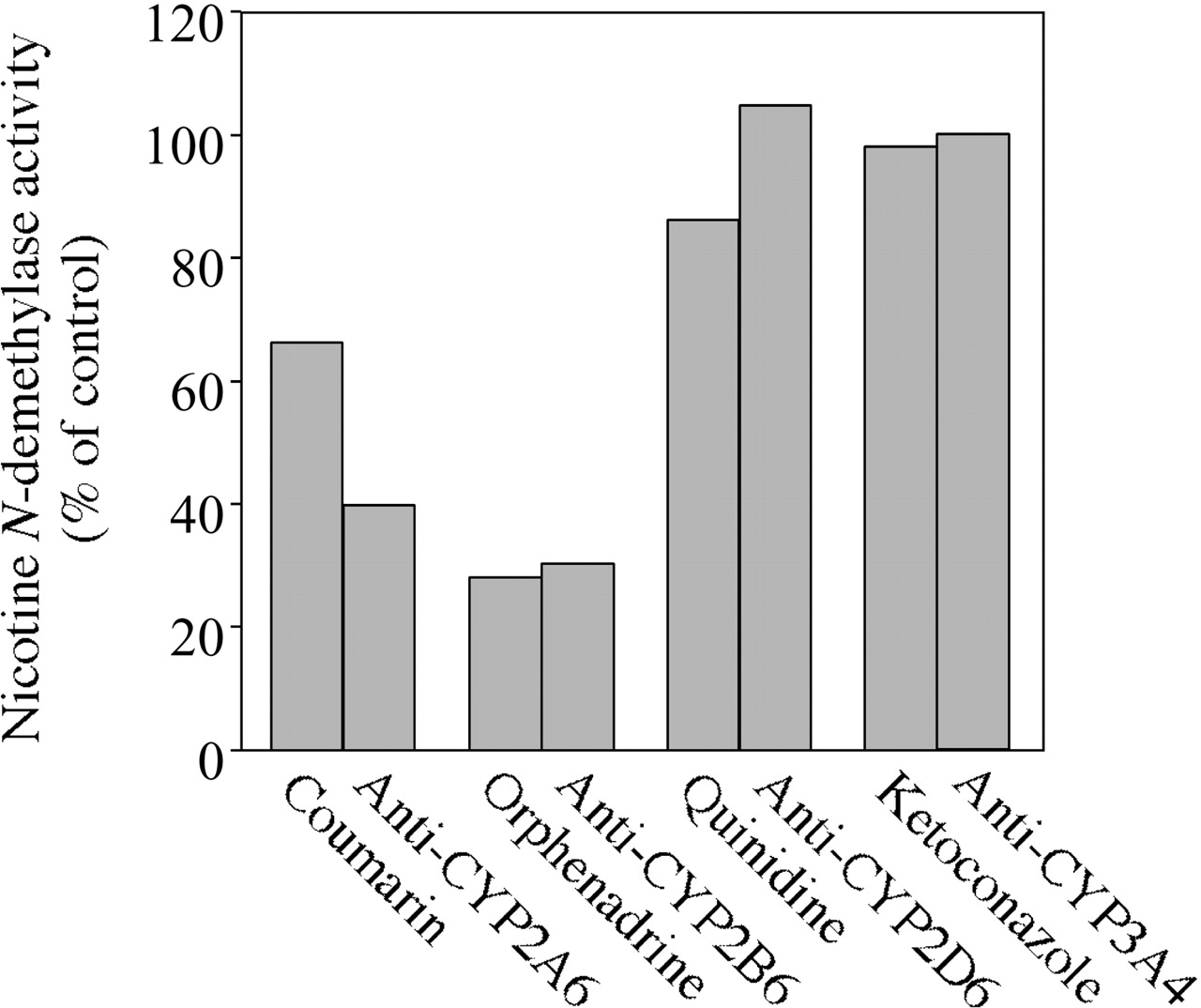
Inhibitory effects of chemical inhibitors and antibodies against CYP2A6, CYP2B6, CYP2D6, or CYP3A4 on the nicotine N-demethylation in pooled human liver microsomes. The substrate concentration was 100 μM. Coumarin (100 μM), orphenadrine (500 μM), quinidine (10 μM), and ketoconazole (1 μM) were used as specific inhibitors for CYP2A6, CYP2B6, CYP2D6, and CYP3A4, respectively. For the immunoinhibition study, the human liver microsomes (0.5 mg/ml) were preincubated with antibodies against each P450 isoform (50 μl/mg) in 0.1 M potassium phosphate buffer (pH 7.4) on ice for 1 h. The nicotine N-demethylase activities were determined as described under Materials and Methods. The control activity in the pooled human liver microsomes was 16.5 pmol/min/mg protein. Each column represents the mean of duplicate determinations.
Next, two samples of human liver microsomes, H003 (CYP2A6-abundant) and H112 (CYP2B6-abundant), were also used for the inhibition study of CYP2A6 and CYP2B6 (Table 2). The nicotine N-demethylase activity in human liver microsomal sample H003 (CYP2A6-abundant) at 20 μM nicotine was markedly inhibited by coumarin to 22% of the control activity and by anti-CYP2A6 antibody to 19% of the control activity. Orphenadrine and anti-CYP2B6 antibody moderately inhibited the activity (35 and 64% of control activity, respectively). The inhibition percentages for the nicotine N-demethylase activity at 100 μM nicotine by coumarin, anti-CYP2A6 antibody, and orphenadrine were almost the same as those at 20 μM nicotine. It is noteworthy that the activity was considerably inhibited by anti-CYP2B6 antibody (28% of control activity). The nicotine N-demethylase activity in human liver microsomal sample H112 (CYP2B6-abundant) at 20 μM nicotine was weakly inhibited by coumarin (81% of control activity) and anti-CYP2A6 antibody (53% of control activity). In contrast, the activity was markedly inhibited by orphenadrine and anti-CYP2B6 antibody (30% and 26% of control activity, respectively). The nicotine N-demethylase activity at 100 μM nicotine was also inhibited to a similar extent.
Inhibitory effects of chemical inhibitors and antibodies against CYP2A6 or CYP2B6 on the nicotine N-demethylase activity in human liver microsomes
Data are means of duplicate determinations. Values in parentheses are percentage of control activity.
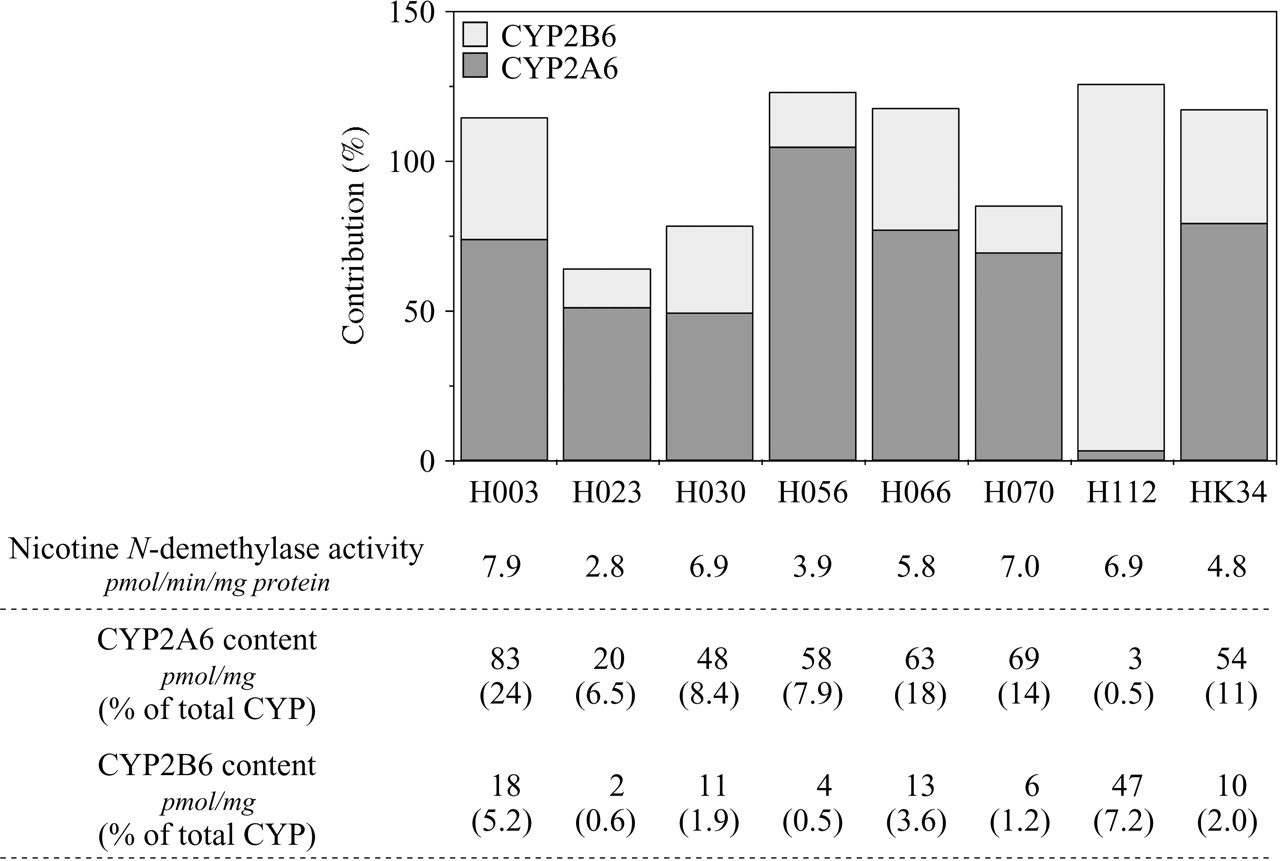
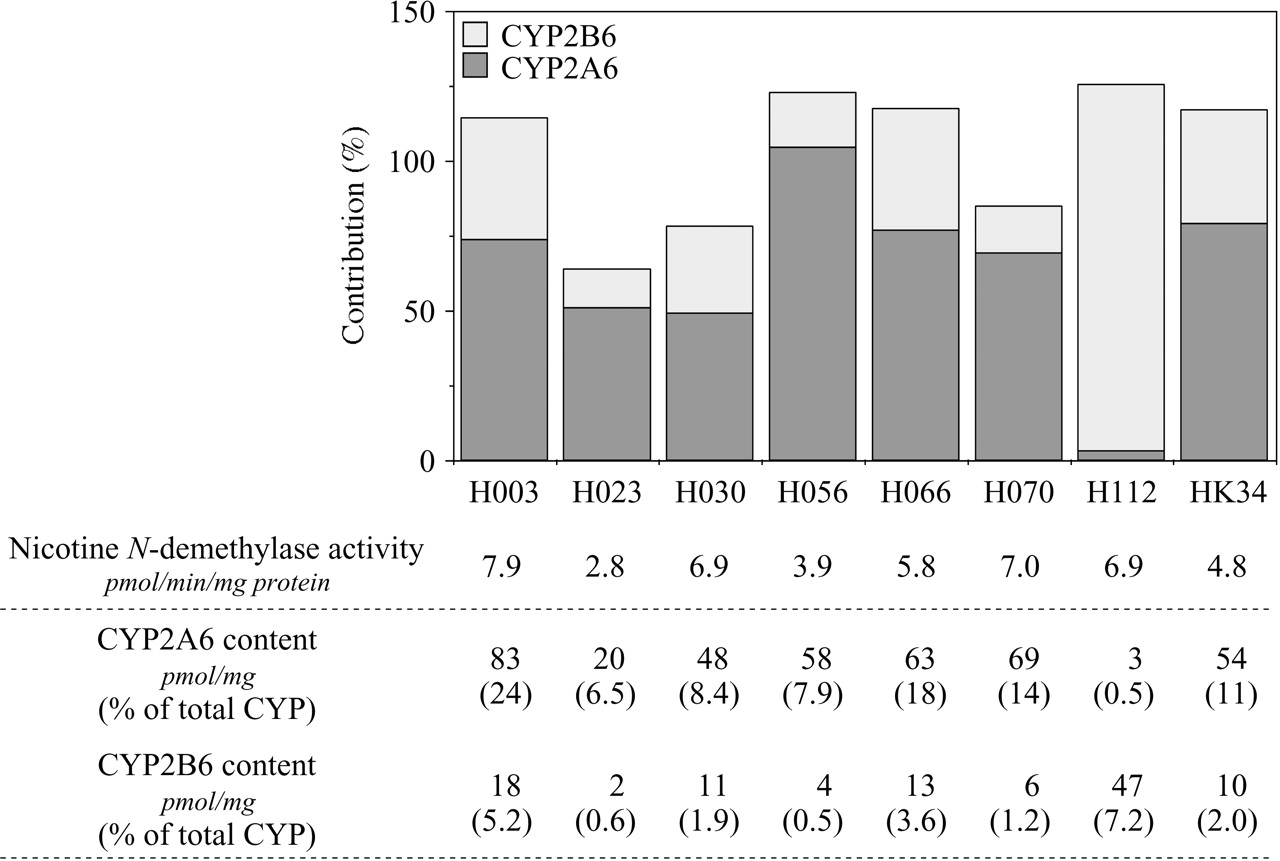
The percentage contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylation in human liver microsomes. The prediction method was based on the activities by recombinant P450s and the contents of each P450 isoform in human liver microsomes. The contributions of CYP2A6 and CYP2B6 in microsomes from eight human livers to nicotine N-demethylase activity at 20 μM nicotine concentration were determined. The total P450 contents determined by CO-difference spectrum and CYP2A6 or CYP2B6 protein contents determined by immunoblot analyses in each human liver microsome were supplied by the manufacturer.
Contributions of CYP2A6 and CYP2B6 to Nicotine N-Demethylase Activity in Human Liver Microsomes. The contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylase activity in microsomes from eight human livers were determined. The nicotine N-demethylase activity in the human liver microsomes (H003, H023, H030, H056, H066, H070, H112 and HK34) at 20 μM nicotine ranged from 2.8 to 7.9 pmol/min/mg protein (Fig. 7). The immunochemically determined contents of CYP2A6 and CYP2B6 (pmol/mg) supplied by the manufacturer are shown in Fig. 7. The Vrec-CYP2A6 and Vrec-CYP2B6 values were 0.07 and 0.18 pmol/min/pmol P450, respectively. Consequently, the contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylase activity in human liver microsomes that are relatively CYP2A6-abundant (those except H112) were estimated as 49 to 104% and 13 to 41%, respectively (Fig. 7). In contrast, the contributions of CYP2A6 and CYP2B6 in the human liver microsomal sample (H112), which is CYP2B6-abundant, were estimated as 3% and 123%, respectively. The sum of the percentages of the contributions of CYP2A6 and CYP2B6 ranged from 64% to 126%.
Nicotine N-Demethylase Activities in Human Brain Microsomes from Striatum. Nicotine N-demethylase activities in human brain microsomes from striatum were determined at 200 μM nicotine. One sample showed a distinct activity (0.9 pmol/min/mg), and the other two samples showed weak activities (0.1 pmol/min/mg). The expression of CYP2A6 and CYP2B6 mRNA in these striatum samples was confirmed with reverse transcription-polymerase chain reaction (data not shown).
Discussion
Recently, Murphy et al. (2005) detected the nornicotine formation from nicotine in recombinant CYP2A6 in baculovirus-infected insect cells with [3H]nicotine. Although they reported that the formed nornicotine was about 5% of the total metabolism of nicotine (Murphy et al., 2005), the absolute activity was unknown. In the present study, we first determined the specific activity for the nicotine N-demethylation in recombinant CYP2A6 (Supersomes), and its CLint was calculated as 5.1 nl/min/pmol P450. It is emphasized that recombinant CYP2B6 revealed a 2.5-fold higher CLint than did CYP2A6. The Km values in the recombinant CYP2A6 and CYP2B6 were close to those for the high- and low-affinity components, respectively, in human liver microsomes. At a relatively low substrate concentration (20 μM nicotine, lower than the Km value for CYP2A6), the nicotine N-demethylase activity was significantly correlated with the content or specific activity for CYP2A6. In addition, it was also correlated with the CYP2B6 specific activity. We confirmed that no significant correlation was observed between CYP2A6 and CYP2B6 in these specific activities and protein contents in the panel of human liver microsomes used in this study (data not shown). At a relatively high substrate concentration (100 μM), the nicotine N-demethylase activity was significantly correlated with the content or specific activity for CYP2B6. These results suggested that the contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylation would be significant at low and high substrate concentrations, respectively.
Murphy et al. (2005) reported that CYP2A13 also could catalyze the nicotine N-demethylation more efficiently than CYP2A6, although the absolute enzymatic activities were not determined. In this study, we found that recombinant CYP2A13 exhibited a 17-fold higher CLint value than recombinant CYP2A6 expressed in E. coli. However, it has been reported that the expression level of CYP2A13 mRNA (7 ± 6 attomoles/mg RNA) was much lower than that of CYP2A6 mRNA (13,000 ± 12,000 attomoles/mg RNA) in human livers (Su et al., 2000). Although we cannot quantify the expression levels of CYP2A13 and CYP2A6 proteins in human livers since antibodies specific for each isoform are not available, it is considered that the expression level of CYP2A13 protein would also be much lower than that of CYP2A6 protein. Thus, the role of CYP2A13 in the nornicotine formation would be mostly insignificant in human livers.
The nicotine N-demethylase activity in human liver microsomes at a nicotine concentration of 100 μM was also significantly correlated with the testosterone 6β-hydroxylase activity (the specific activity for CYP3A4). In the panel of human liver microsomes used in this study, a fortuitous correlation between the testosterone 6β-hydroxylase activity and CYP2B6 protein content (r = 0.809, p < 0.001) was also observed. Furthermore, the nicotine N-demethylase activity in human liver microsomes was not inhibited by ketoconazole and anti-CYP3A4 serum. These results suggested that the involvement of CYP3A4 in the nicotine N-demethylase activity in human liver microsomes would be negligible.
Quantitative analysis of the contributions of each P450 isoform using the activities by recombinant P450s and contents of each P450 isoform in human liver microsomes revealed that the contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylation would be dependent on the individual expression levels of these isoforms. The sum of the percentage contributions of CYP2A6 and CYP2B6 ranged from 64% to 126%, indicating that these P450 isoforms are the major enzymes responsible for the nicotine N-demethylation in humans. One limitation of this prediction method is that the P450 contents were measured by immunoblot analysis. Immunoblot analysis cannot distinguish between active and inactive protein. Another prediction method is using the relative activity factor with specific activities for each P450 (Crespi, 1995; Nakajima et al., 1999). However, the relative activity factor method for predicting the CYP2B6 contribution was not successful (the estimated contributions of CYP2B6 in all human liver microsomes were over 100%), possibly because of the limited specificity of CYP2B6 substrates such as S-mephenytoin or benzyloxyresorufin (data not shown). The inhibition analyses with chemicals and antibodies against CYP2A6 and CYP2B6 in the present study would support the contributions of CYP2A6 and CYP2B6 to the nicotine N-demethylase activity in human liver microsomes.
Previously, we demonstrated that the urinary excretion levels of nornicotine in subjects entirely lacking the CYP2A6 gene were similar to those in normal subjects, suggesting that the involvement of CYP2A6 in the nornicotine formation was slight (Yamanaka et al., 2004). However, the present in vitro study demonstrated that CYP2A6 has a significant catalytic activity for nornicotine formation as a high-affinity component. Therefore, the nornicotine formed in the subjects lacking CYP2A6 would result from the compensatory role of CYP2B6.
In the present study, the CLint for the nornicotine formation in human liver microsomes was calculated as 0.3 μl/min/mg. Previously, we demonstrated that the CLint for the cotinine formation in human liver microsomes was 1.6 μl/min/mg (Nakajima et al., 1996). The difference in the intrinsic clearances between the two metabolic pathways would reflect the metabolic profile of nicotine in human urine, i.e., nornicotine, 2 to 3%, and cotinine, 70 to 80% (Benowitz et al., 1994).
Nornicotine is a major metabolite of nicotine in brain (Crooks et al., 1997). In the human brain, CYP2B6 protein is expressed in various regions including the occipital cortex, hippocampus, striatum, cerebellar vermis, and cerebellar hemisphere (Gervot et al., 1999; Miksys et al., 2003). In addition, Miksys and Tyndale (2004) recently reported that CYP2A6 mRNA is also expressed in various regions of the human brain. In particular, the expression levels of CYP2B6 protein and CYP2A6 mRNA are high in the striatum (Miksys et al., 2003; Miksys and Tyndale 2004). These backgrounds prompted us to investigate the nicotine N-demethylase activities in human brain microsomes from striatum. Consequently, the nicotine N-demethylase activity was detected in human brain microsomes from striatum. The present study demonstrated that nornicotine in brain could have originated from the nicotine N-demethylation locally in the brain, in addition to the nornicotine that is formed in the liver and subsequently passes the blood-brain barrier. It has been reported that nicotine can induce CYP2B in rat brain (Anandatheerthavarada et al., 1993; Miksys et al., 2000). In smokers, a higher level of CYP2B6 protein in the brain was detected than in nonsmokers, suggesting that nicotine can also induce human CYP2B6 in the brain (Miksys et al., 2003). Until now, there has been no information on whether CYP2A6 is inducible by nicotine. Generally, the induction of CYP2B is mediated by constitutive androstane receptor or pregnane X receptor. These receptors usually cross talk with each other toward target genes. Recently, Lamba et al. (2004) reported that nicotine could activate human pregnane X receptor. Collectively, the induction of CYP2B6 by nicotine would enhance the metabolism of nicotine to nornicotine.
In summary, we found that CYP2A6 and CYP2B6 catalyze the nornicotine formation from nicotine in human liver microsomes. These contributions would be dependent on the individual expression levels of CYP2A6 and CYP2B6.
Acknowledgments
We acknowledge Brent Bell for reviewing the manuscript.
Footnotes
-
This study was supported in part by Philip Morris Incorporated and by Research Fellowships of the Japan Society for the Promotion of Science for Young Scientists.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.006254.
-
ABBREVIATIONS: P450, cytochrome P450; NPR, NADPH-cytochrome P450 reductase; HPLC, high performance liquid chromatography; LC-MS/MS, liquid chromatography-tandem mass spectrometry; CLint, intrinsic clearance.
- Received June 24, 2005.
- Accepted August 31, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}