


Abstract
We have cloned and functionally characterized the rat ortholog of multidrug and toxin extrusion type transporter 1 (rMATE1). The mRNA of rMATE1 was strongly expressed in kidney and detectable in the various tissues such as brain, stomach, colon, lung, liver, spleen, skeletal muscle, and prostate. When stably expressed in HEK293 cells, rMATE1 could mediate the transport of tetraethylammonium (TEA) and cimetidine under the condition where the membrane potential was disrupted by a high concentration of potassium ion and intracellular pH was reduced by NH4Cl pretreatment. When extracellular pH was changed from 5.5 to 8.5, the transport of TEA by rMATE1 was greatest at pH 7.5. Kinetic analyses showed that the transports of TEA and cimetidine mediated by rMATE1 were both saturable with a Km of 260 ± 10 and 3.01 ± 0.21 μM, respectively. It was found that cimetidine is the most potent inhibitor of rMATE1, and many other organic cations, such as 1-methyl-4-phenylpyridinium, amiloride, imipramine, and quinidine, are also effective as inhibitors. Pretreatment of the cells expressing rMATE1 with p-chloromercuribenzene sulfonate significantly reduced TEA transport, but this effect was totally reversed by subsequent treatment with dithiothreitol. These results indicate that the functional nature of rMATE1 is consistent with that of the hypothetical organic cation/H+ antiporter system in the brush-border membrane of the renal tubular epithelial cells. Accordingly, these results suggest that rMATE1 is an electroneutral and multispecific organic cation transporter energized by the trans-proton gradient, and plays a physiological role in renal secretion of organic cations, including clinically used cationic drugs.
Urinary excretion is a major detoxication pathway for xenobiotics and endogenous metabolites in the body. It is well recognized that transporters that mediate secretion of such compounds in renal tubular epithelium play critical roles in this detoxication pathway (Pritchard and Miller, 1996; Wright and Dantzler, 2004). In particular, organic cations such as tetraethylammonium (TEA) are known to be actively secreted from blood to urine across renal tubular epithelial cells by a set (or multiple sets) of specific transporters at two membranes, basolateral membrane and brush-border membrane (Rennick, 1981).
The basolateral transporters are named organic cation transporters (OCTs) and are known to be driven by inside-negative membrane potential. OCT1 was identified as the first member of these transporters from the rat kidney (Grundemann et al., 1994) and OCT2, which is homologous to OCT1, was also isolated from the rat kidney (Okuda et al., 1996). Human orthologs of OCT1 and OCT2 have also been cloned (Gorboulev et al., 1997; Zhang et al., 1997). However, it was later shown that OCT1 is far more abundant in the liver than in the kidney, whereas OCT2 is mainly expressed in the kidney. It is also proved that OCT2 is localized at the basolateral membrane of renal proximal tubules and mediates the transport of a broad range of diverse organic cations (Urakami et al., 1998; Karbach et al., 2000). Thus, it is now generally accepted that OCT2 is the major molecular entity of the organic cation transport system at the basolateral membrane.
The brush-border transporters, which mediate the subsequent transport of organic cations from the cell interior into urine, have been characterized less clearly. Brush border organic cation transport is known to depend on a pH gradient across the luminal membrane, and has been suggested to be mediated by an electroneutral organic cation/H+ antiporter(s) that can exchange intracellular organic cations with extracellular proton (Koepsell et al., 2003). Tamai et al. (1997) cloned OCTN1 (novel organic cation transporter 1), which can mediate pH-dependent TEA transport, from the human fetal liver. OCTN1 is also expressed in the human kidney, being localized at the brush-border membrane of renal proximal tubules (Tamai et al., 1997). Although the substrate specificity of OCTN1 is similar to that of the putative organic cation/H+ antiporter suggested by studies using isolated brush-border membrane vesicles, functional identity with the organic cation/H+ antiporter is in question because the expression of OCTN1 mRNA is relatively low in the human kidney, and TEA transport by OCTN1 is electrogenic, depending on inside-negative membrane potential (Motohashi et al., 2002; Tamai et al., 2004).
More recently, Otsuka el al. (2005) have identified novel transporters of the multidrug and toxin extrusion (MATE) type, human MATE1 (hMATE1) and mouse MATE1 (mMATE1). They are significantly homologous to MATE family transporters conferring multidrug resistance on bacteria, and are primarily expressed in the kidney and liver, where they are localized at the luminal membranes of the renal tubules and bile canaliculi (Otsuka et al., 2005). Since the MATE1s of both animals can mediate pH-dependent TEA transport and are inhibited by various cationic compounds known to inhibit the putative organic cation/H+ antiporter, it is suggested that MATE1 could be the molecular entity of the long sought antiporter. However, it is not clear yet whether the hMATE1-mediated transport of TEA and other organic cations requires a proton gradient as driving force, because it has so far been demonstrated only that extracellular alkalinization can enhance its transport activity.
In the present study, we report on the cloning and detailed functional characterization of rat MATE1 (rMATE1). We also provide evidence for the electroneutral organic cation/H+ antiport nature of rMATE1-mediated transport.
Materials and Methods
Materials. [14C]Tetraethylammonium bromide (2.4 mCi/mmol), [3H]cimetidine (25.0 Ci/mmol), and [α-32P]dCTP (3000 Ci/mmol) were obtained from GE Healthcare Bio-Sciences Co. (Piscataway, NJ). [3H]Clonidine (66.0 Ci/mmol), [3H]estrone sulfate (57.3 Ci/mmol), and [3H]taurocholate (1.2 Ci/mmol) were obtained from PerkinElmer Life Sciences, Inc. (Boston, MA). [3H]l-Carnitine (81.0 Ci/mmol) was obtained from Moravek Biochemicals, Inc. (Brea, CA). Tetraethylammonium, cimetidine, and geneticin were obtained from Sigma-Aldrich Co. (St. Louis, MO). Restriction enzymes were obtained from TOYOBO (Tokyo, Japan). All other reagents were of analytical grade and commercially obtained.
Isolation of rMATE1. Total RNA was prepared from the rat kidney by a guanidine isothiocyanate extraction method (Chomczynski and Sacchi, 1987). A reverse transcription reaction was carried out using 3 μg of total RNA, an oligo(dT) primer, and ReverTra Ace (TOYOBO) as a reverse transcriptase. The cDNA of rMATE1 was isolated from the rat kidney cDNA by PCR using KOD polymerase (TOYOBO) and the following primers: rMATE1-F1 (forward primer), 5′-GGCCTGTGGGTCCCCGTTCC-3′; rMATE1-R1 (reverse primer), 5′-TGGATTTGTCAGGCCTTAACTT-3′. PCR was performed using the following conditions: 94°C for 1 min; 35 cycles of (i) 94°C for 40 s, (ii) 60 °C for 40 sec, and (iii) 72 °C for 1.5 min. The PCR product was subcloned into pME18 vector, and transferred into a mammalian expression vector, pCI-neo (Promega, Madison, WI). The sequence was determined with an automated sequencer ABI PRISM 3100 (Applied Biosystems, Foster City, CA). The sequence was analyzed using the National Center for Biotechnology Information server (www.ncbi.nlm.nih.gov/).
Northern Blot Analysis. Tissue distribution of rMATE1 transcripts in the rat was studied by Northern blots of total RNA samples. In brief, 25 μg of total RNA isolated by guanidine isothiocyanate extraction from various rat tissues was separated on 1% formaldehyde-agarose gels and transferred to a Hy-bond-N+ nylon membrane (GE Healthcare Bio-Sciences Co.). The UV cross-linked membrane was then hybridized at high stringency using rMATE1-specific cDNA probe labeled with [α-32P]dCTP using a random primer DNA labeling kit (Amersham Corp).
RT-PCR Analysis. Total RNA isolated from various rat tissues was used for cDNA synthesis. PCR reactions were performed using TaqDNA polymerase (Invitrogen, CA) and the following primers: rMATE1-F2, 5′-TTC AGT GGG ACA TGG CCT ATC T-3′; rMATE1-R2, 5′-GAC AGG CGA GTT TCC AAT TAA GC -3′. PCR conditions were as follows: 94 °C for 2 min; 35 cycles of 1) 94°C for 30 s, 2) 58°C for 30 s, and 3) 72°C for 1 min.
Cell Culture and Stable Transfection. HEK293 cells were maintained at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium with 10% fetal calf serum, 100 units/ml penicillin, and 100 μg/ml streptomycin. Cells were transfected with the plasmid carrying rMATE1 cDNA by the calcium phosphate coprecipitation method (Wigler et al., 1977) and cultured in Dulbecco's modified Eagle's medium containing 10% fetal calf serum and 800 μg/ml geneticin for 2 to 3 weeks. Antibiotic-resistant clones were selected and tested for the transport of cimetidine.
Transport Study in HEK293 Cells. For transport assays, HEK293 cells stably expressing rMATE1 and mock cells (1.5 × 105 cells/well initially) were grown on 24-well plates coated with poly-l-lysine and cultured for 24 to 36 h. To assess the function of rMATE1, which was expected to operate as an organic cation/H+ antiporter driven by trans-proton gradient, by the uptake of probe substrates into the cells, we used a method to generate an outward H+ gradient by reducing the intracellular pH (Boron and De Weer, 1976). Before transport assays, cells were washed with uptake buffer (140 mM KCl, 0.4 mM KH2PO4, 0.8 mM MgSO4, 1.0 mM CaCl2, 25 mM glucose, and 10 mM HEPES, pH 7.4), incubated in uptake buffer containing 20 mM NH4Cl (1.5 ml) for 10 min, and thereafter incubated in uptake buffer (NH4Cl-free, 1.5 ml) for 5 min. Transport assays were started by replacing the substrate-free uptake buffer with one containing a 3H-labeled substrate. All the procedures were conducted at 37°C. Assays were stopped by addition of ice-cold substrate-free uptake buffer (2 ml), and the cells were washed two times with 2 ml of the same buffer. The cells were solubilized in 0.5 ml of 0.2 N NaOH solution containing 0.5% SDS, and associated radioactivity was measured by liquid scintillation counting. Cellular protein content was determined by the method of Lowry et al. (1951) using bovine serum albumin as the standard. When the influence of extracellular pH on uptake was examined, uptake buffers of different pH values were prepared by appropriately altering the concentrations of Tris, Mes, and Hepes.
Determination of pHi using BCECF. The intracellular pH (pHi) was evaluated by using 2′,7′-bis-(carboxyethyl)-carboxyfluorescein (BCECF), a pH-sensitive fluorescent dye. For the measurements of pHi, rMATE1 cells were cultured on coverslips for 1 day. The cells were then incubated at room temperature in the dark in Na+-based uptake buffer (135 mM NaCl, 5 mM KCl, 0.4 mM KH2PO4, 0.8 mM MgSO4, 1.0 mM CaCl2, 25 mM glucose, and 10 mM HEPES, pH 7.4), containing 5 μM membrane-permeable BCECF acetoxymethyl ester (BCECF/AM) for 30 min, and washed with BCECF/AM-free Na+-based uptake buffer and stored in the dark for an additional 30-min period to ensure the release of membrane-impermeable and fluorescent BCECF by the hydrolysis of BCECF/AM. For the ratiometric measurement of pHi, BCECF was excited at the wavelengths of 450 and 490 nm, and fluorescent emission was measured at 540 ± 25 nm. The pHi was calibrated using standard solutions of pH 6.0, 6.5, 7.0, 7.5, 8.0, 8.5 and 9.0, which were prepared using the K+-based standard uptake buffer containing nigericin (10 μM).
Data Analysis. To estimate kinetic parameters for the saturable transport of TEA and cimetidine, the following equation was fitted to the uptake rate versus concentration profiles by means of nonlinear least-squares regression analysis using WinNonlin (Pharsight Co., Mountain View, CA): v = Vmax × s/(Km + s), where v and s are the uptake rate and concentration, respectively, of TEA or cimetidine, and Km and Vmax are the half-saturation concentration (Michaelis constant) and the maximum transport rate, respectively. Results are presented as means ± S.E., and statistical analysis was performed using two-tailed, unpaired Student's t test or, when multiple comparisons were needed, analysis of variance followed by Dunnett's test, with p < 0.05 considered significant.
Results
Structural Features of Rat MATE1. BLAST (Basic Local Alignment Search Tool) searches of the National Center for Biotechnology Information databases were performed using the amino acid sequence of ALF5, which encodes a member of the MATE family that has been shown to be involved in the resistance of Arabidopsis to toxic tetramethylammonium (Diener et al., 2001). By combining PCR cloning and bioinformatics analysis, we isolated an orthologous cDNA sequence in the rat (GenBank accession number AAH88413). During this study, human and mouse orthologs of the cDNA sequence have been identified as MATE 1. Therefore, the rat cDNA was designated as rMATE1. The cDNA encodes a putative protein of 566 amino acids, which have 30, 93, and 78% similarities with ALF5, mMATE1, and hMATE1, respectively. Hydropathy analyses using the programs SOSUI (http://sosui.proteome.bio.tuat.ac.jp/~sosui/proteome/sosuiframe0.html), TopPred (http://www.sbc.su.se/~erikw/toppred2/), TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/), and HMMTOP (http://www.enzim.hu/hmmtop/html/document.html) consistently predicted 13 potential membrane-spanning domains with a large extracellular loop between transmembrane domains XII and XIII. Putative extracellular N-glycosylation sites were not found.

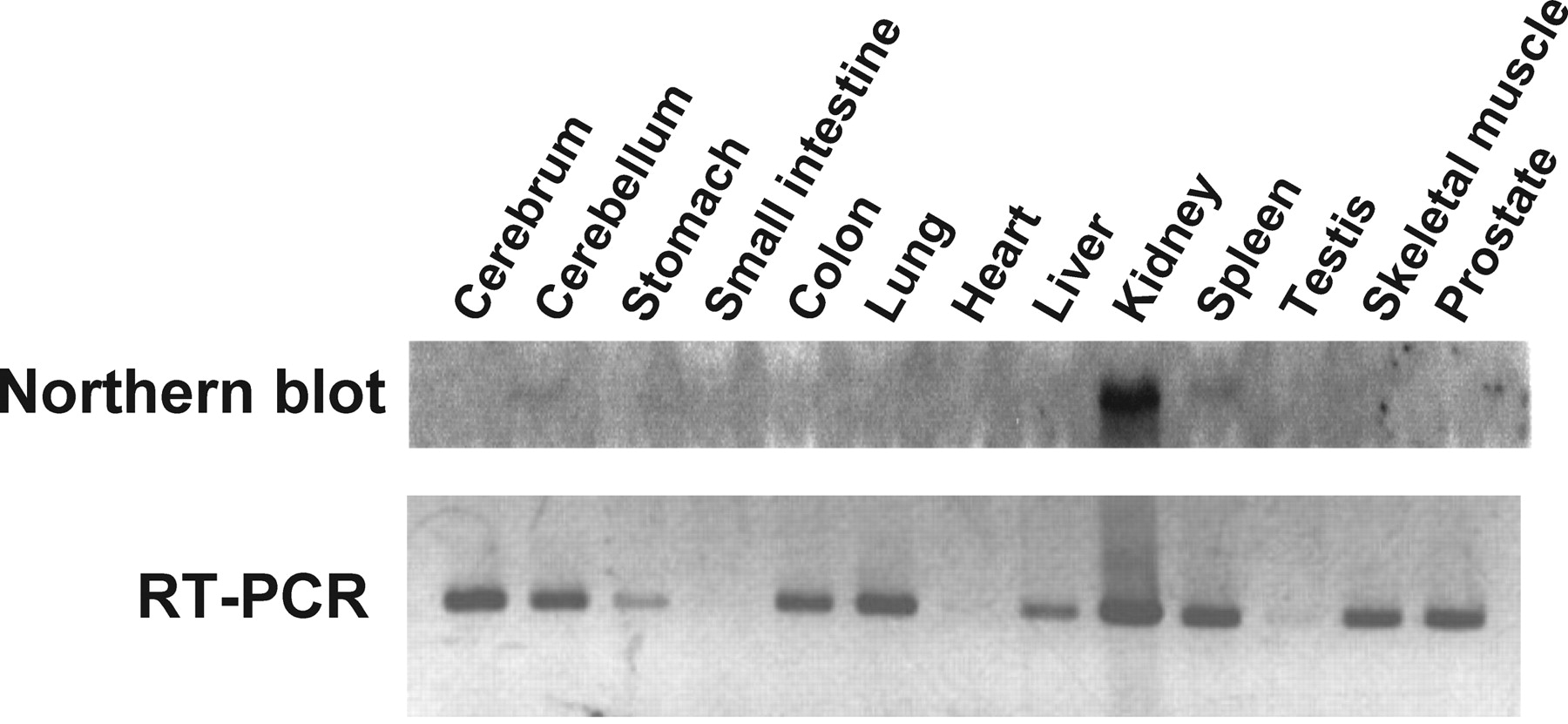
Northern blot and RT-PCR analyses for rMATE1 expression. Total RNA from various rat tissues was probed with 32P-labeled rMATE1 cDNA in Northern blot analysis. Aliquots (3 μg) of total RNA from various rat tissues were reverse transcribed and then amplified by PCR using a set of specific primers for rMATE1 in RT-PCR analysis.
Tissue Expression Pattern of MATE1 mRNA in the Rat. To analyze the tissue distribution of rMATE1, we performed Northern blot analysis and RT-PCR in rat tissues (Fig. 1). Northern blot analysis shows that MATE1 mRNA (∼3.0 kilobase pairs) is strongly expressed only in kidney with a single transcript, and any signal is not detected in other tissues. This result indicates that kidney is the primary site of function for rMATE1. But RT-PCR revealed that rMATE1 is also expressed in various tissues, such as brain, stomach, colon, lung, liver, spleen, skeletal muscle, and prostate, as well as kidney, but not in small intestine, heart, and testis. Since it has been reported that the expression levels of mMATE1 are relatively high in liver and brain as well as kidney, as detected by Northern blot analysis (Otsuka et al., 2005), there may be some species differences in the tissue distribution of MATE1 among rodents.
Functional Characteristics of rMATE1. We first determined the transport activity of various compounds in HEK293 cells stably expressing rMATE1 (rMATE1 cells) and also for comparison in mock cells (Fig. 2). Under the condition where pHi was reduced by NH4Cl pretreatment, it was found that TEA and cimetidine are transported by rMATE1 very efficiently, exhibiting 10- to 17-fold greater uptake in rMATE1 cells compared with that in mock cells after 20 s of uptake. Appreciable rMATE1-mediated uptake was not observed for clonidine as a hydrophobic cation, l-carnitine as a zwitterion, and organic anions such as taurocholate and estrone sulfate. These results suggest that rMATE1 is a multispecific transporter involved in the transport of hydrophilic organic cations.
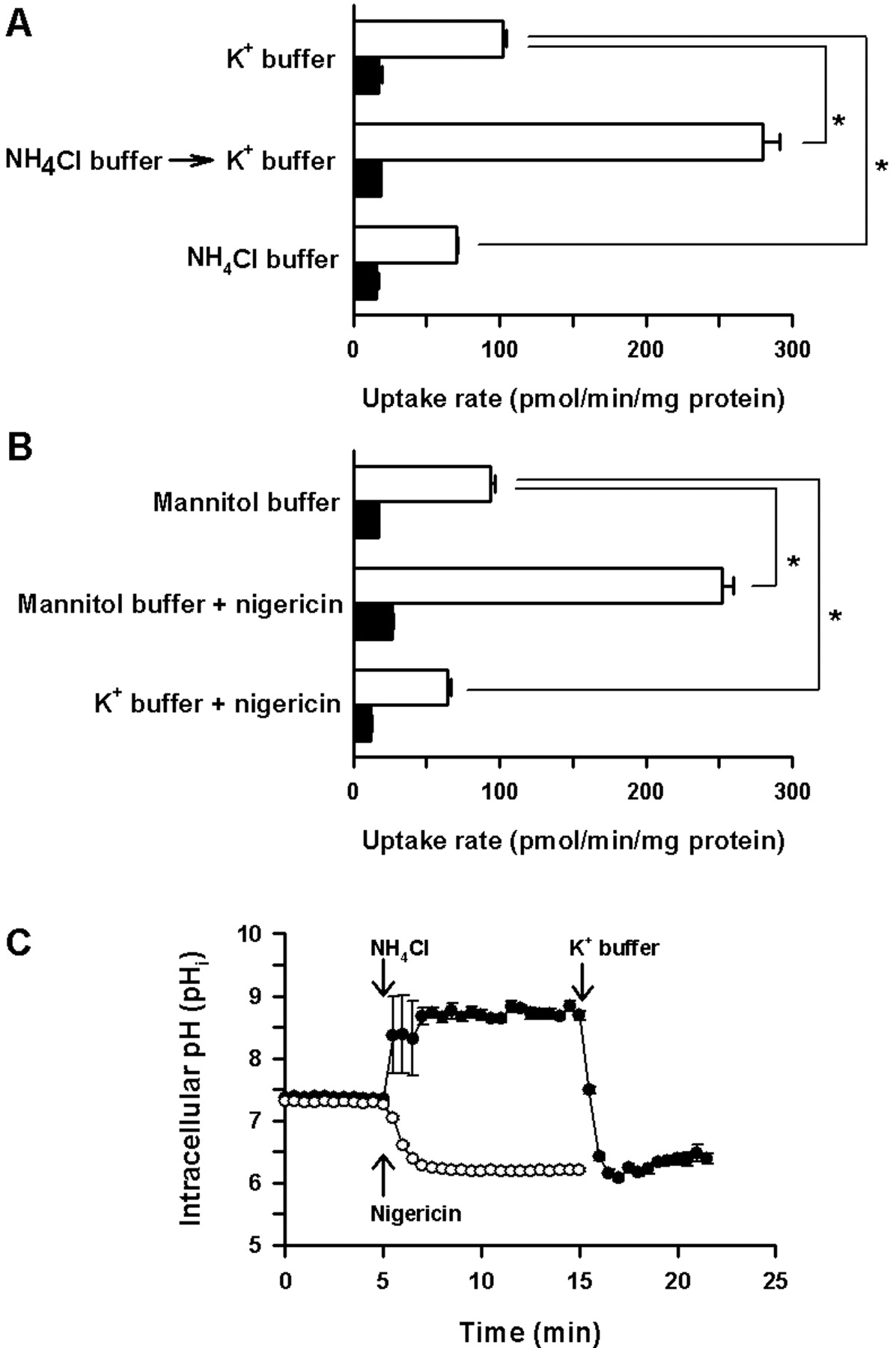
To examine whether an outwardly directed proton gradient is required for rMATE1 to function, a series of TEA uptake experiments were conducted using cells with different pHi conditions accomplished by various treatments using NH4Cl (Fig. 3A) or nigericin (Fig. 3B). Significant rMATE1-mediated uptake of TEA was observed when the K+-based standard uptake buffer (K+ buffer) was used during the uptake period for cells not pretreated with NH4Cl (Fig. 3A). When cells were pretreated in the K+ buffer containing NH4Cl (NH4Cl buffer) and subsequently with the K+ buffer before the uptake period with the K+ buffer, TEA uptake was enhanced in rMATE1 cells, but not in mock cells. The NH4Cl buffer contains NH +4 and NH3 in equilibrium, but only NH3 can diffuse into the cell interior easily. Therefore, when cells are in NH4Cl buffer, intracellular pH rises because of the deprivation of protons by protonation of NH3 in the cells. When the extracellular medium is changed to the K+ buffer, which is free of NH4Cl (free of NH3 as well as NH +4), intracellular NH3 rapidly diffuses out of the cells, resulting in the accumulation of protons released from NH +4 in a process to produce NH3 and compensate for its loss in the cells. Therefore, considering these intracellular events, the increase in TEA uptake by the NH4Cl pretreatment observed only in rMATE1 cells indicates that an outwardly directed proton gradient accelerates rMATE1-mediated TEA uptake. When the cells were pretreated in the NH4Cl buffer for 15 min (10-min and subsequent 5-min period) continuously until the transport assay was initiated, TEA uptake was reduced to the lowest level, as expected, as a result of intracellular alkalinization, which imposes a reversed proton gradient higher outside. Similar results were obtained when nigericin was used to alter pH conditions (Fig. 3B). When the mannitol-based uptake buffer (mannitol buffer), in which KCl was replaced with mannitol, was used during the uptake period in the absence of nigericin, a K+/H+ exchanger (ionophore), some appreciable rMATE1-mediated TEA uptake was observed. When the mannitol buffer was used in the presence of nigericin for intracellular acidification, TEA uptake in rMATE1 cells was increased to a level comparable to that observed for the NH4Cl pretreatment, whereas TEA uptake in mock cells was unchanged. In the presence of nigericin in the K+ buffer for the pH-clamped condition, on the other hand, TEA uptake in rMATE1 cells was decreased to a level lower than that in the mannitol buffer without nigericin. However, interestingly, the uptake was still higher than that in mock cells, even though the pHi should be clamped to the same value as the extracellular pH. This result indicates that rMATE1 can also mediate TEA transport in the absence of proton gradient.

Substrate specificity of the rMATE1 expressed in HEK293 cells. rMATE1-specific uptake was evaluated by dividing the uptake in rMATE1-transfected cells by that in mock cells. Uptake of [14C]TEA (7.3 μM), [3H]cimetidine (16 nM), [3H]clonidine (6.1 nM), [3H]l-carnitine (4.6 nM), [3H]estrone-3-sulfate (7.0 nM), and [3H]taurocholate (340 nM) was evaluated at 37°C and pH 7.4 for 20 s in mock and rMATE1 cells. Data are represented as the means ± S.E. (n = 3 or 4).
As shown in Fig. 3C, pHi of rMATE1 cells rose above pH 8.5 when the cells were incubated in the K+ buffer containing NH4Cl. When the medium was changed to one without NH4Cl, pHi rapidly declined below pH 6.5. The treatment with nigericin in the mannitol buffer also reduced pHi to a similar level. Such acidified conditions were maintained throughout the period of pHi monitoring. These results indicate that our uptake experiments, which were performed 5 min after the initiation of acidification, were suitably designed to evaluate uptake at the stably acidified intracellular condition.

Effect of intracellular pH on [14C]TEA uptake in rMATE1-transfected HEK293 cells. Uptake of [14C]TEA (10 μM) was evaluated at 37°C for 20 s in rMATE1 cells (open bar) and mock cells (closed bar). In A, cells were incubated at 37°C in the K+-based standard uptake buffer (K+ buffer, pH 7.4) for 15 min (10-min and subsequent 5-min period), in the K+ buffer containing 20 mM NH4Cl (NH4Cl buffer) for 10 min, and subsequently in the K+ buffer (NH4Cl-free) for 5 min, or in the NH4Cl buffer for 15 min (10-min and subsequent 5-min period), before TEA uptake was evaluated using the K+ buffer containing [14C]TEA. In B, cells were incubated in the K+ buffer or mannitol-based buffer (pH 7.4), in which KCl was replaced with mannitol, in the absence or presence of nigericin (10 μg/ml) at 37°C for 5 min, as indicated, before TEA uptake was evaluated using each buffer added with [14C]TEA. Shown in C are profiles of pHi monitored in rMATE1 cells loaded with BCECF, a pH-sensitive fluorescent dye, and acidified by sequential treatment using NH4Cl buffer and K+ buffer (closed circles) or nigericin in mannitol buffer (open circles). Data are represented as the means ± S.E. (n = 3 for A and B, and n = 10 for C). *, significantly different from the value for control at p < 0.05.
When extracellular pH was changed from 5.5 to 8.5, the transport of TEA by rMATE1 was greatest at pH 7.5 in cells pretreated with NH4Cl for intracellular acidification and also in those untreated (Fig. 4). The uptake rate decreased significantly when pH became more alkaline or more acidic than the pH optimal for transport.
Kinetic Features of rMATE1-Mediated Transport. The time courses of TEA uptake were examined in rMATE1 cells and mock cell pretreated with NH4Cl for intracellular acidification. The uptake of TEA increased with time rapidly in rMATE1 cells, whereas it was far slower in mock cells, being only minimal even at 5 min. Since the rate of rapid TEA uptake in rMATE1 cells began declining after 20 s (Fig. 5), we decided to evaluate the initial rate of uptake at 20 s. Based on similar observation (data not shown), initial cimetidine uptake was evaluated also at 20 s. As shown in Fig. 6, kinetic analyses indicated that the transports of TEA and cimetidine mediated by rMATE1 were both saturable with a Km of 260 ± 10 and 3.01 ± 0.21 μM, respectively, and a Vmax of 5.4 ± 0.2 nmol/min/mg protein and 87.9 ± 3.7 pmol/min/mg protein, respectively, as the computer-fitted parameters with S.E.

Effect of extracellular pH on [14C]TEA uptake in rMATE1-transfected HEK293 cells. Uptake of [14C]TEA (10 μM) was evaluated at 37°C for 20 s in mock cells (▴) with NH4Cl pretreatment for intracellular acidification and in rMATE1-transfected cells with (•) or without (▪) NH4Cl pretreatment. Cells were incubated in uptake buffer (pH 7.4) in the absence or presence of 20 mM NH4Cl for 10 min at 37°C and subsequently in uptake buffer (NH4Cl-free) for 5 min at 37°C before TEA uptake was evaluated using uptake buffer containing [14C]TEA. Data are represented as the means ± S.E. (n = 3).
Substrate Specificity of rMATE1. To find specific inhibitors, which include potential substrates, of rMATE1 and to help clarify its substrate specificity, we examined the effect of various compounds (0.2 or 2.0 mM) on the rMATE1-mediated uptake of [14C]TEA (10 μM) and [3H]cimetidine (50 nM), as shown in Table 1. It was found that cimetidine is the most potent inhibitor. Many other organic cations, such as MPP, thiamine, amiloride, diltiazem, imipramine, quinidine, quinine, and verapamil, also inhibited the transport mediated by rMATE1 extensively. Clonidine and estrone sulfate, which were not observed to be transported by rMATE1 in the initial set of experiments (Fig. 2), also showed significant inhibitory effect. Corticosterone, which possesses no positively charged group, also inhibited the transport significantly. However, the transport was not inhibited by NMN, consistent with the result reported for hMATE1, even though it is an endogenous organic cation that has long been suggested to be secreted by the organic cation/H+ antiport system.
Effect of various compounds on the uptake of [14C]TEA and [3H]cimetidine in rMATE1-transfected HEK293 cells
Uptake of [14C]TEA (10 μM) and [3H]cimetidine (50 nM) was evaluated at 37°C and pH 7.4 for 20 s. Uptake rates for control were 0.20 nmol/min/mg protein and 1.78 pmol/min/mg protein, respectively, for TEA and cimetidine. Data are represented as the mean ± S.E. (n = 4).
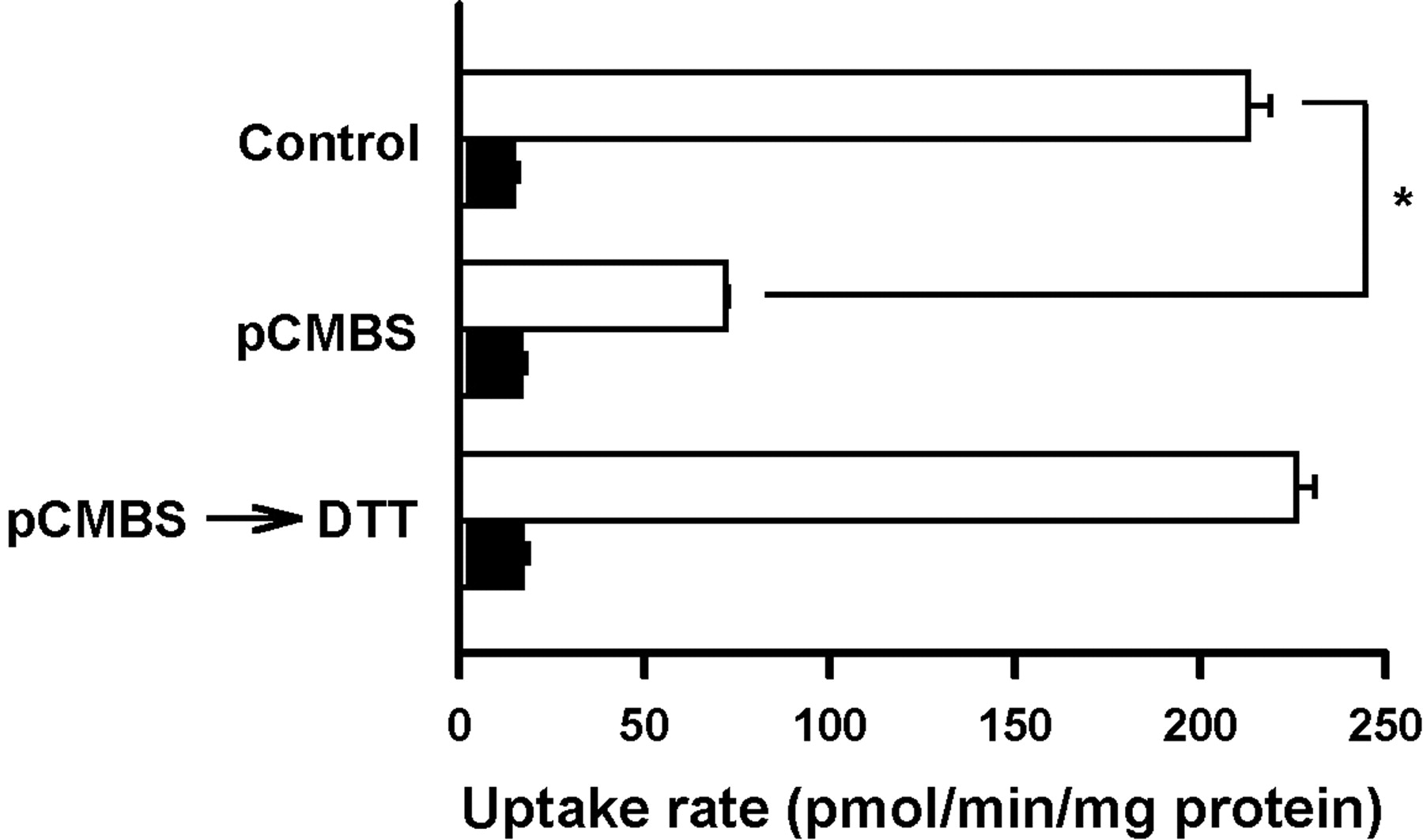
Effect of pCMBS on the TEA Transport by rMATE1. pCMBS (p-chloromercuribenzene sulfonate), an organic mercurial compound, is known to be a potent inhibitor of the renal organic cation/H+ antiport system (Hori et al., 1987). As shown in Fig. 7, exposure of rMATE1 cells to the medium containing pCMBS significantly reduced TEA transport. But the effect of pCMBS was totally reversed by subsequent treatment with dithiothreitol. These results indicate that sulfhydryl groups play an important role in the transport function of rMATE1.
Discussion
We have reported on the cloning and functional characterization of rMATE1, presenting evidence to support a physiological role of rMATE1 as an organic cation/H+ antiporter. rMATE1 is highly expressed in the kidney (Fig. 1), where organic cations are processed for secretion into urine, and this transport activity is stimulated in the presence of proton gradient across the plasma membrane (Fig. 3).
In most experiments, we generated an outwardly directed proton gradient by means of pretreatment with NH4Cl for intracellular acidification to assess the transport of organic cations as an influx process. This transport mimics the efflux of intracellular organic cations to outside the cells, which is the final step in renal tubular secretion. Because physiological proton gradient is generated by Na+/H+ exchanger at the brush-border membrane of renal proximal tubules, with the luminal fluid maintained more acidic than cytosol, at pH 6.7 to 6.8, it has been presumed that the organic cation/H+ antiport system at the brush-border membrane can use the proton gradient as a driving force to mediate the efflux of organic cations into urine (Wright and Dantzler, 2004). Our results clearly showed that the transport of TEA mediated by rMATE1 was significantly increased by intracellular acidification, suggesting that rMATE1 facilitates the exchange of extracellular TEA with intracellular proton. We used the K+-based uptake buffer (K+ buffer) as a standard uptake buffer to exclude the influence of extracellular Na+ because Na+/H+ exchanger isoforms NHE-1, NHE-2, and NHE-3, which are constitutively expressed in HEK293 cells, and to which rMATE1 was heterologously introduced, can work to counteract the intracellular acidification treatment by mediating the efflux of H+ in exchange for Na+ (Lang et al., 2003). Under this experimental condition, a high concentration of extracellular K+ could depolarize the membrane potential and, hence, disrupt electrogenic transport. However, TEA uptake for the K+ buffer was comparable to that for the mannitol-based buffer (Fig. 3) and also to that for a buffer in which K+ was replaced with Na+ (data not shown). Further modulation of the membrane potential by the addition of valinomycin, a K+ ionophore, to K+ buffer and Na+-based uptake buffer did not alter TEA uptake, either (data not shown). These results suggest that TEA transport mediated by rMATE1 is not electrogenic, involving no net flux of electric charge. This is consistent with an earlier finding by Sokol et al. (1985) that, in brush-border membrane vesicles of the canine kidney, organic cation/H+ antiport is independent of the membrane potential, involving exchange of organic cation for proton with stoichiometry of 1:1. Another finding in the present study, that pCMBS reversibly inhibited TEA uptake mediated by rMATE1 (Fig. 7), was also consistent with an earlier study by Hori et al. (1987) reporting that sulfhydryl group-modifying reagents can potently inhibit the proton gradient-dependent transport of TEA in rat renal brush-border membrane vesicles. All these facts indicate that the functional nature of rMATE1 is consistent with that of the hypothetical organic cation/H+ antiporter system in the brush-border membrane of the renal tubular epithelial cells.

Time course of [14C]TEA uptake in rMATE1-transfected HEK293 cells. Uptake of [14C]TEA (7.3 μM) was evaluated at 37°C and pH 7.4 in mock cells (•) and rMATE1-trnsfected cells (○). Data are represented as the means ± S.E. (n = 3).

Concentration dependence of the uptake of [14C]TEA and [3H]cimetidine by rMATE1 expressed in HEK293 cells. The uptake of [14C]TEA (A) and [3H]cimetidine (B) was evaluated at 37°C and pH 7.4 for 20 s in rMATE1-transfected cells and mock cells pretreated with NH4Cl for intracellular acidification. The rMATE1-specific uptake was calculated by subtracting the uptake in mock cells from that in rMATE1-trnsfected cells, and was used for kinetic analysis. Solid lines represent the computer-fitted profiles. Eadie-Hofstee plots are shown in insets, where v and s represent the uptake rate (pmol/min/mg protein) and concentration (μM), respectively.

Effect of pCMBS on the uptake of [14C]TEA in rMATE1-transfected HEK293 cells. Before NH4Cl pretreatment, cells were incubated in uptake buffer (pH 7.4) in the absence or presence of pCMBS (0.1 mM) at 37°C for 5 min and subsequently in uptake buffer in the absence or presence of dithiothreitol (DTT; 0.5 mM) at 37°C for 5 min. The uptake of [14C]TEA (10 μM) was evaluated at 37°C and pH 7.4 for 20 s in rMATE1-transfected cells (open bars) and mock cells (closed bars). Data are represented as the means ± S.E. (n = 3). *, significantly different from the value for control at p < 0.05.
It should be noted that the specific transport of TEA mediated by rMATE1 occurred, although only moderately, when pHi was not modified and when proton gradient across the membrane was collapsed by using the K+ buffer containing nigericin (Fig. 3B), indicating that rMATE1 can operate to some extent, even in the absence of a proton gradient. Studies using renal brush-border membrane vesicles have shown that the organic cation/H+ antiport system can also mediate the exchange of organic cations for organic cations instead of proton (Holohan and Ross, 1980). Therefore, it is possible that the transport of TEA mediated by rMATE1 is in part driven by some endogenous organic cations. Villalobos and Braun (1998) reported that endogenous organic cations such as choline, acetylcholine, and guanidine trans-stimulated TEA transport in avian renal brush-border membrane vesicles, although they were poorly transported as substrates (Villalobos and Braun, 1998). Moreover, we found that the mRNA of rMATE1 is expressed, although to lesser extents, in various tissues including spleen, skeletal muscle, and prostate, where no pH gradient seems to exist across the membrane (Fig. 1). An organic cation/organic cation exchange mechanism could be a possible mode of operation for rMATE1 in these tissues.
It is interesting that the profile of TEA uptake versus extracellular pH in rMATE1 cells treated for intracellular acidification was slightly different from that in untreated rMATE1 cells (Fig. 4). In the latter, TEA transport was optimal at a range of pH from 7.5 to 8.0, similar to results reported for human MATE1 (Otsuka et al., 2005). Intracellular acidification in rMATE1 cells resulted in a narrower pH optimum at 7.5. The uptake in intracellularly acidified cells decreased with pH more extensively at higher pH levels and, at pH 8.5, reached a level close to that in untreated cells, indicating that a greater proton gradient does not lead to a greater TEA transport at such higher pH levels. Since the pKa of TEA is relatively high (pKa = 13), TEA is supposed to be almost totally dissociated in the entire pH range. Therefore, changes in the degree of TEA dissociation do not seem to be the cause of the reduced uptake at higher pH levels. It is also unlikely that changes in pH affect the affinity of TEA for rMATE1. It may be more likely that an elevation of extracellular pH causes a reduction in the activity of rMATE1 by decreasing the affinity for proton.
Transport assessments of various radiolabeled compounds revealed that TEA and cimetidine are favored as substrates by rMATE1, but organic anions are not (Fig. 2). Kinetic analyses revealed that the Km values for TEA and cimetidine are 260 ± 10 μM and 3.01 ± 0.21 μM, respectively. These values are comparable to those reported by previous studies using renal brush-border membrane vesicles of the rat (0.21 mM for TEA; Maeda et al., 1993) and the rabbit (0.33 mM and 4.6 μM, respectively, for TEA and cimetidine; Gisclon et al., 1987; Rafizadeh et al., 1987). Thus, also in terms of these kinetic features, rMATE1-mediated transport is consistent with that of the renal organic cation/H+ antiport system, which can mediate the transport of various compounds that possess a positively charged group(s) in their chemical structures (Ullrich and Rumrich, 1996). In addition, the fact that TEA and cimetidine, which are not endogenous organic cations, are efficiently transported by rMATE1 suggests that it is a multispecific transporter having a physiological role of eliminating xenobiotics from the body. Rennick and colleagues demonstrated that the renal organic cation/H+ antiport system had a greater affinity for xenobiotics than did endogenous compounds (Acara and Rennick, 1972; Acara et al., 1977; Besseghir et al., 1981). Indeed, the transport of TEA and cimetidine mediated by rMATE1 was significantly inhibited by various organic cations (Table 1). These inhibition profiles are similar to those reported previously using renal brush-border membrane vesicles of the rat, rabbit, and human. Moreover, MPP, a very typical probe cation used in this kind of study, inhibited the uptake of TEA and cimetidine in rMATE1 cells, suggesting that MPP may share this transporter. In contrast, NMN did not show any inhibitory effect, although it has also been known as a cation considered to share the renal organic cation/H+ antiport system with TEA (Wright, 1985). This might be due to its lower affinity (Km = 2.0 mM) for the organic cation/H+ antiport system (Hsyu et al., 1988). However, we cannot exclude the possibility that there may be another unidentified organic cation/H+ antiporter that can transport NMN. Hydrophobic organic cations such as thiamine, amiloride, clonidine, diltiazem, imipramine, quinidine, quinine, and verapamil were also found to be effective as inhibitors of rMATE1. But these compounds may not be very suitable as substrates of rMATE1. In the case of clonidine, for example, we could not observe any appreciable transport in rMATE1 cells (Fig. 2), even though more than half of its molecules are dissociated in the experimental condition, since its pKa is 7.8. Its significant inhibitory effect on the transport of TEA and cimetidine (Table 1) may suggest that it can be a noncompetitive inhibitor of rMATE1, and function as a “blocker”. A similar case is reported for rat OCT2. Although TEA uptake mediated by rat OCT2 was inhibited by quinine, which is a hydrophobic and weak base (pKa = 8.4), it was found not to be a substrate (Arndt et al., 2001). Thus, it will obviously be necessary to evaluate the transport of each organic cation that showed inhibitory effect in this study, to identify substrates of rMATE1.
In general, efficient renal secretion of organic cations could be achieved by coordinated operation of transporters located at the basolateral membrane and brush-border membrane in renal tubular epithelial cells. Although the substrate specificity of rat OCT2, the basolateral transporter, seems to be similar to that of rMATE1, rat OCT2 seems to have a higher affinity for organic cations than rMATE1. In the case of TEA, the Km value of 45 μM for rat OCT2 (Urakami et al., 1998) is 5-fold smaller, indicating greater affinity, than that of 260 μM for rMATE1. Therefore, it is probable that OCT2 mediates concentrative transport of organic cations into epithelial cells at the basolateral membrane, and subsequently, MATE1 mediates the efflux of those accumulated intracellularly, accomplishing transcellular transport across the renal epithelial cells physiologically.
In conclusion, we have cloned and characterized rMATE1, which can mediate the organic cation transport energized by the trans-proton gradient. It was demonstrated that this transporter mediates the transport of TEA and cimetidine with affinities comparable to those reported for the organic cation/H+ antiport system in the kidney, and is inhibited by pCMBS. Thus, it is likely that rMATE1 plays a physiological role in renal secretion of organic cations, including cationic drugs used clinically. Since the efflux of a given substrate (organic cation) across the luminal membrane is the final and possibly rate-limiting step in its transepithelial secretion process in renal proximal tubules, the characterization of MATE1 may help us predict and manage adverse events; for example, those originating from drug-drug interactions in renal tubular secretion.
Footnotes
-
This work was supported in part by a Grant-in-Aid for Young Scientists (B) from the Ministry of Education, Culture, Sports, Science and Technology, Japan (#18790125), a Grant-in-Aid from the Nakatomi Foundation, and a Grant-in-Aid for Research in Nagoya City University.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.106.010876.
-
ABBREVIATIONS: TEA, tetraethylammonium; OCT, organic cation transporter; OCTN, novel organic cation transporter; BCECF, 2′,7′-bis-(carboxyethyl)-carboxyfluorescein; AM, acetoxymethyl ester; MPP, 1-methyl-4-phenylpyridinium; MATE, multidrug and toxin extrusion; hMATE1, human MATE transporter 1; mMATE1, mouse MATE transporter 1; rMATE1, rat MATE transporter 1; Mes, 4-morpholineethanesulfonic acid; RT-PCR, reverse transcription-polymerase chain reaction; NMN, N1-methylnicotinamide; pCMBS, p-chloromercuribenzene sulfonate; NHE, Na+/H+ exchanger.
- Received May 1, 2006.
- Accepted August 21, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}