


Abstract
Aryl hydrocarbon receptor (AhR), constitutive androstane receptor (CAR), pregnane X receptor (PXR), peroxisome proliferator-activated receptor-α (PPARα), and nuclear factor-E2-related factor 2 (Nrf2) are transcription factors that mediate xenobiotic induction of biotransformation enzymes and transporters. The purpose of this study was to determine the tissue distribution and xenobiotic induction of these transcription factors and their associated target genes in mice. Many of these transcription factors were most highly expressed in extrahepatic tissues. CAR expression in female liver was twice that in male liver. This corresponded with greater induction of the CAR target genes Cyp2b10 and multidrug resistance-associated protein (Mrp) 4 by the CAR activator 1,4-bis-[2-(3,5-dichloropyridyloxy)]benzene (TCPOBOP) in female liver than in male liver. Mice were treated with xenobiotic activators of AhR, CAR, PXR, PPARα, or Nrf2 and their associated marker genes were highly induced in liver by these xenobiotic activators. Transcription factor target gene induction occurred with minimal induction of their associated transcription factors. CAR expression was induced by the AhR ligand 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), leading to increased basal expression of Cyp2b10 mRNA and enhanced induction of Cyp2b10 by TCPOBOP. Mrp2, 3, and 4 induction was augmented by cotreatment with TCDD and TCPOBOP compared with treatment with either compound alone. These studies illustrate CAR induction by TCDD in mice, indicating that AhR may transcriptionally regulate CAR and thus enhance induction of key metabolism and transporter genes by the CAR activator TCPOBOP. Collectively, these studies illustrate the fact that some xenobiotic inducers may elicit their response through mechanisms involving transcription factor regulation.
Emerging evidence illustrates the fact that physiological regulation of transporters and biotransformation enzymes is under the control of transcription factors that are responsive to an array of xenobiotics and endogenous activators (Xie and Evans, 2001; Staudinger et al., 2003; Klaassen and Slitt, 2005). Specifically, the aryl hydrocarbon receptor (AhR), constitutive androstane receptor (CAR), pregnane-X receptor (PXR), peroxisome proliferator-activated receptor-α (PPARα), and nuclear factor-E2-related factor 2 (Nrf2), are transcription factors through which classic microsomal enzyme inducers (MEIs) and other chemicals increase the expression of transporters and biotransformation enzymes (Klaassen and Slitt, 2005).
The AhR is activated by dioxin and other aromatic hydrocarbons, leading to activation of target gene transcription through binding to xenobiotic response elements found within their promoter regions (Reyes et al., 1992; Harper et al., 2006). AhR activation markedly induces mRNA expression and enzyme activity of cytochromes P450 (P450s) CYP1A1 and CYP1A2 in rats (Kedderis et al., 1991) and Cyp1a1 and Cyp1a2 in mice (Beebe et al., 1990). Furthermore, AhR is also important in regulating inducible expression of xenobiotic-metabolizing enzymes including NAD(P)H quinone oxidoreductase 1 (Nqo1), aldehyde dehydrogenase 3a1, UDP glucuronosyl transferase (Ugt) 1a6, and glutathione S-transferase (Gst) α1 (Nebert et al., 2000).
The orphan nuclear receptor CAR mediates Cyp2b10 induction in response to phenobarbital (PB) and 1,4-bis-[2-(3,5-dichloropyridyloxy)] benzene (TCPOBOP) in mice (Wei et al., 2000). CAR is also associated with xenobiotic-induced transactivation of human CYP2B6 and CYP3A4 (Kawamoto et al., 1999). Target gene activation by CAR is mediated through binding to phenobarbital response element modules found within target gene promoter regions (Honkakoski and Negishi, 1997). Although CAR is a critical transcription factor mediating P450 induction, CAR also mediates hepatic induction of some phase II xenobiotic-metabolizing enzymes such as Ugt1a1, Gstα1, Gstα2, and sulfotransferase 2a1 (Huang et al., 2003; Assem et al., 2004). CAR has also been implicated in the induction of canalicular export transporters in liver, namely the multidrug resistance-associated proteins (Mrp) Mrp2, Mrp3, and Mrp4 (Cherrington et al., 2002, 2003; Johnson and Klaassen, 2002; Slitt et al., 2003; Assem et al., 2004; Maher et al., 2005a). Furthermore, CAR activation by TCPOBOP augmented cholic acid induction of mouse biotransformation enzymes and uptake transporters (Guo et al., 2003). Thus, xenobiotics that activate CAR can regulate both xenobiotic metabolism and transport.
PXR is an orphan nuclear receptor that facilitates hepatic and intestinal induction of Cyp3a expression by xenobiotics, synthetic glucocorticoids, and naturally occurring steroids (Kliewer et al., 1998), a function mediated through PXR response element binding. The pivotal role of PXR in regulating induction of Cyp3a11 and organic anion-transporting polypeptide 2 (new nomenclature, Oatp1a4) in mice by the PXR agonist pregnenolone-16α-carbonitrile (PCN) and by lithocholic acid (a bile acid), was established using PXR-null mice (Staudinger et al., 2001).
The nuclear receptor PPARα regulates target gene transactivation through binding to peroxisome proliferator response elements (reviewed by Kliewer et al., 1994). Mouse PPARα mediates peroxisome proliferation and hepatomegaly in response to the classic peroxisomal proliferators Wy-14,643 and clofibrate (Lee et al., 1995). As evidenced by studies in PPARα-null mice, PPARα also mediates hepatic mRNA induction of fatty acyl-CoA oxidase and the fatty acid β-oxidation and ω-oxidation genes, Cyp4a10 and Cyp4a14, respectively (Lee et al., 1995; Barclay et al., 1999).
Nrf2 is an oxidative stress and antioxidant inducible transcription factor that mediates target gene transactivation through antioxidant response element (ARE) binding (McMahon et al., 2001). Nrf2 is thought to regulate expression of a battery of antioxidant genes including its classic target gene Nqo1 (Venugopal and Jaiswal, 1996), Gsts, and the glutamate cysteine ligase catalytic and modifier subunits (McMahon et al., 2001). Furthermore, Nrf2 has been implicated in regulation of inducible expression of the Mrp1 (Hayashi et al., 2003) and Mrp2 (Vollrath et al., 2006) transporters in mice. Nrf2 is thus an important regulator of both xenobiotic metabolism and transport.
AhR, CAR, PXR, PPARα, and Nrf2 are important molecular mediators that regulate expression of xenobiotic-metabolizing enzymes and transporters. In the present study, the tissue distribution and induction of these key transcription factors and their associated target genes was investigated in mice to better understand the expression of these genes in vivo. The present studies illustrate transcription factor induction by xenobiotics that are known to induce biotransformation enzymes and transporters and thus imply an important role for transcription factors in xenobiotic induction of drug metabolism and disposition.
Materials and Methods
Animals. Male and female C57BL/6 mice (age 7 weeks, 18-20 g) were obtained from Charles River Laboratories, Inc. (Wilmington, MA) and were housed in an American Animal Association for Laboratory Animal Care accredited facility. Mice were given access to Teklad 8064 rodent chow (Harlan, Madison, WI) and water ad libitum. Mice were acclimated to the housing facility for 1 week before experiments (four mice per cage, 50% relative humidity, 12-h light/dark cycle).
Chemicals. 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) was a gift from Dr. Karl Rozman (University of Kansas Medical Center, Kansas City, KS). Oltipraz was a gift from Dr. Steve Safe (Texas A&M University, College Station, TX). 3,3′,4,4′,5-Pentachlorobiphenyl (PCB-126) was purchased from AccuStandard (New Haven, CT). All other chemicals were obtained from Sigma-Aldrich (St. Louis, MO). Names, designated abbreviations, and CAS Registry numbers for all chemicals are cited in Table 1.
Dosing regimen, target receptors, and marker genes of xenobiotic activator compounds used for inducer studies
Compounds were dosed i.p. or by oral gavage (p.o.).
Transcription Factor Tissue Distribution. Eight-week-old male and female C57BL/6 mice were euthanized, and the following organs collected: liver, kidney, lung, stomach, duodenum, jejunum, ileum, large intestine, brain, heart, testes, ovaries, and uterus. Placentae were isolated from pregnant C57BL/6 female mice 2 days before parturition. The contents of the stomach and intestinal segments were removed by cutting these tissues open and washing them in 0.9% saline. All tissues were snap-frozen in liquid nitrogen and stored at -80°C before RNA isolation. All data represent n = 5 per tissue. Heart and lung data represent n = 5 of three pooled organs each. Placenta data represent n = 5 of five pooled organs each. Ovary data represent n = 5 of 25 pairs of pooled organs each.
Transcription Factor, Cyp, and Mrp Induction Studies. Eight-week-old male and female C57BL/6 mice were treated with microsomal enzyme inducers of AhR, CAR, PXR, PPARα, and Nrf2. The microsomal enzyme inducer compounds, dosing regimens, target receptors, and marker genes are given in Table 1. Control animals were dosed with either corn oil or saline vehicle. Mice were treated for 4 days using an injection volume of 5 ml/kg (n = 5 per treatment). Livers were removed from animals on day 5, snap-frozen in liquid nitrogen, and stored at -80°C.
TCDD and TCPOBOP Coexposure Studies. Eight-week-old male C57BL/6 mice were treated with TCDD (AhR ligand) and/or TCPOBOP (CAR activator) over the course of 4 days. Treatment groups were as follows: 1) corn oil: corn oil i.p. (5 ml/kg) once per day for 4 days and one corn oil i.p. injection (5 ml/kg) 12 h before organ harvesting; 2) TCDD: 37 μg/kg/day TCDD i.p. for 4 days and one corn oil i.p. injection (5 ml/kg) 12 h before organ harvesting; 3) TCPOBOP: corn oil i.p. for 4 days, and a single 3 mg/kg i.p. TCPOBOP dose 12 h before organ harvesting; and 4) TCDD plus TCPOBOP: 37 μg/kg/day TCDD i.p. for 4 days and a single 3 mg/kg i.p. TCPOBOP dose 12 h before harvesting. All treatments were given using an injection volume of 5 ml/kg (n = 5 per treatment). Corn oil was used as a vehicle for all treatment groups. All mice were treated once a day for 4 days and were given a final treatment 12 h before organ harvesting. Livers were removed, snap-frozen in liquid nitrogen, and stored at -80°C.
Total RNA Isolation. Total RNA was isolated from frozen mouse tissues using the RNA-Bee RNA isolation reagent (Tel-Test Inc., Friendswood, Texas), as per the manufacturer's protocol. RNA pellets were resuspended in diethylpyrocarbonate-treated deionized water. The RNA concentration was evaluated spectrophotometrically by its absorbance at 260 nm. Agarose gel electrophoresis was used to evaluate RNA integrity by observation of 18S and 28S ribosomal RNA bands visualized via ethidium bromide fluorescence under ultraviolet illumination.
Branched DNA Analysis of Gene Expression. The bDNA method has been used extensively in our laboratory, and the methods have been described in detail previously (Maher et al., 2005b). Multiple sequence-specific probes were developed to each mRNA transcript of interest (Table 2). Probe sequences for Mrp2, Mrp3, and Mrp4 were published previously (Maher et al., 2005b), as were the probe sequences for Cyp1a1, Cyp2b10, Cyp3a11, Cyp4a14, and Nqo1 (Cheng et al., 2005). Gene expression analysis was performed using the QuantiGene Screen bDNA signal amplification kit (Genospectra, Fremont, CA), and luminescence was quantified using a Quantiplex 320 bDNA luminometer interfaced with Quantiplex data management software (version 5.02; Bayer Corp., Diagnostics Div., Tarrytown, NY). All data are expressed as relative luminescence units per 10 μg of total RNA.
List of oligonucleotide probes generated for use in bDNA signal amplification assay
Statistics. Data from male and female mice were compared in the tissue distribution study using Student's t test. CAR target gene analysis between males and females or between control and treated animals were also analyzed using Student's t-tests. Control groups (corn oil p.o., corn oil i.p., and saline i.p.) for the transcription factor inducer study were analyzed by one-way analysis of variance, followed by a Bonferroni post hoc test. This analysis indicated no statistical difference between corn oil or saline controls, regardless of route of exposure. Controls (n = 13) were thus pooled for the remaining analyses. Inducer data were evaluated for statistical significance using analysis of variance, followed by Duncan's multiple range post hoc test. All statistical flags represent a statistically significant difference (p < 0.05), as described in the figure legends.
Results
Tissue Distribution of Transcription Factors in Mice. AhR mRNA was ubiquitously expressed in the tissues analyzed in this study, with the highest expression observed in the lung (Fig. 1). Expression of AhR mRNA did not exhibit gender differences in any of the tissues examined. CAR mRNA expression was highest in liver, followed by duodenum, jejunum, lung, heart, and placenta (Fig. 1). CAR expression was female-predominant in liver and male-predominant in brain and heart. Expression of PXR mRNA was highest in the intestinal tract and was moderately expressed in liver (Fig. 1). PXR expression exhibited a slight, but statistically significant female predominance in liver and brain. PPARα mRNA expression was highest in liver, kidney, and heart (Fig. 1). PPARα was moderately expressed in the intestinal tract, lung, and gonads and exhibited a slight female predominance in lung and duodenum (Fig. 1). Expression of Nrf2 mRNA was highest in the stomach, followed by the intestinal tract and lung (Fig. 1). Nrf2 was moderately expressed in liver, kidney, heart, ovary, uterus, and placenta. Nrf2 exhibited a female predominance in the lung and in gonads, as expression was higher in the ovary than in the testes.
Induction of Transcription Factor and Marker Gene mRNA in Liver by Microsomal Enzyme Inducers. AhR was not consistently induced by any class of transcriptional inducers. However, AhR mRNA expression in liver was induced 250% by the AhR activator β-naphthoflavone (BNF), but not by other AhR activators (Fig. 2). AhR was induced 170% by both the PXR activator spironolactone (SPR) and the PPARα activator clofibrate (CLOF) and was induced 300% by the Nrf2 activator butylated hydroxyanisole (BHA) (Fig. 2). The AhR target gene Cyp1a1 was highly induced (up to 1200-fold) by AhR activators, but not by any activators of CAR, PXR, PPARα,or Nrf2 (Fig. 2).
Hepatic expression of CAR mRNA was induced by all three AhR activators: 160% by TCDD, 120% by PCB-126, and 60% by BNF (Fig. 3). CAR expression was doubled by the PPARα activator 2-diethylhexyl phthalate (DEHP) and was modestly induced by the Nrf2 activator BHA (Fig. 3). CAR expression, however, was not induced by activators of CAR or PXR. The CAR marker gene Cyp2b10 was highly induced by CAR activating compounds: 118-fold by TCPOBOP, 13-fold by diallyl sulfide (DAS), and 17-fold by PB (Fig. 3). The AhR ligand TCDD modestly induced expression of Cyp2b10 (Figs. 3 and 8). The PXR activators PCN and dexamethasone (DEX) induced Cyp2b10 mRNA expression by 9.4- and 25.7-fold, respectively. The PPARα activators CLOF and DEHP induced Cyp2b10 mRNA expression 3.5- and 2.8-fold, respectively. Furthermore, the Nrf2 activators oltipraz (OPZ), ethoxyquin (EXQ), and BHA induced expression of Cyp2b10 by 8.9-, 9.1-, and 4.9-fold, respectively (Fig. 3).
Expression of PXR in liver was modestly induced by the PXR activator SPR, by the PPARα activators CLOF and DEHP, and by the Nrf2 activator BHA (Fig. 4). Neither AhR nor CAR activators altered expression of PXR (Fig. 4). Expression of the PXR target gene Cyp3a11 was reduced 41% by the AhR ligand BNF (Fig. 4). The CAR activators TCPOBOP, DAS, and PB induced Cyp3a11 expression 190, 140, and 110%, respectively (Fig. 4). Cyp3a11 was induced markedly by PXR activators: 320% by PCN, 200% by SPR, and 310% by DEX (Fig. 4). Expression of Cyp3a11 was modestly induced by the PPARα activator CLOF.
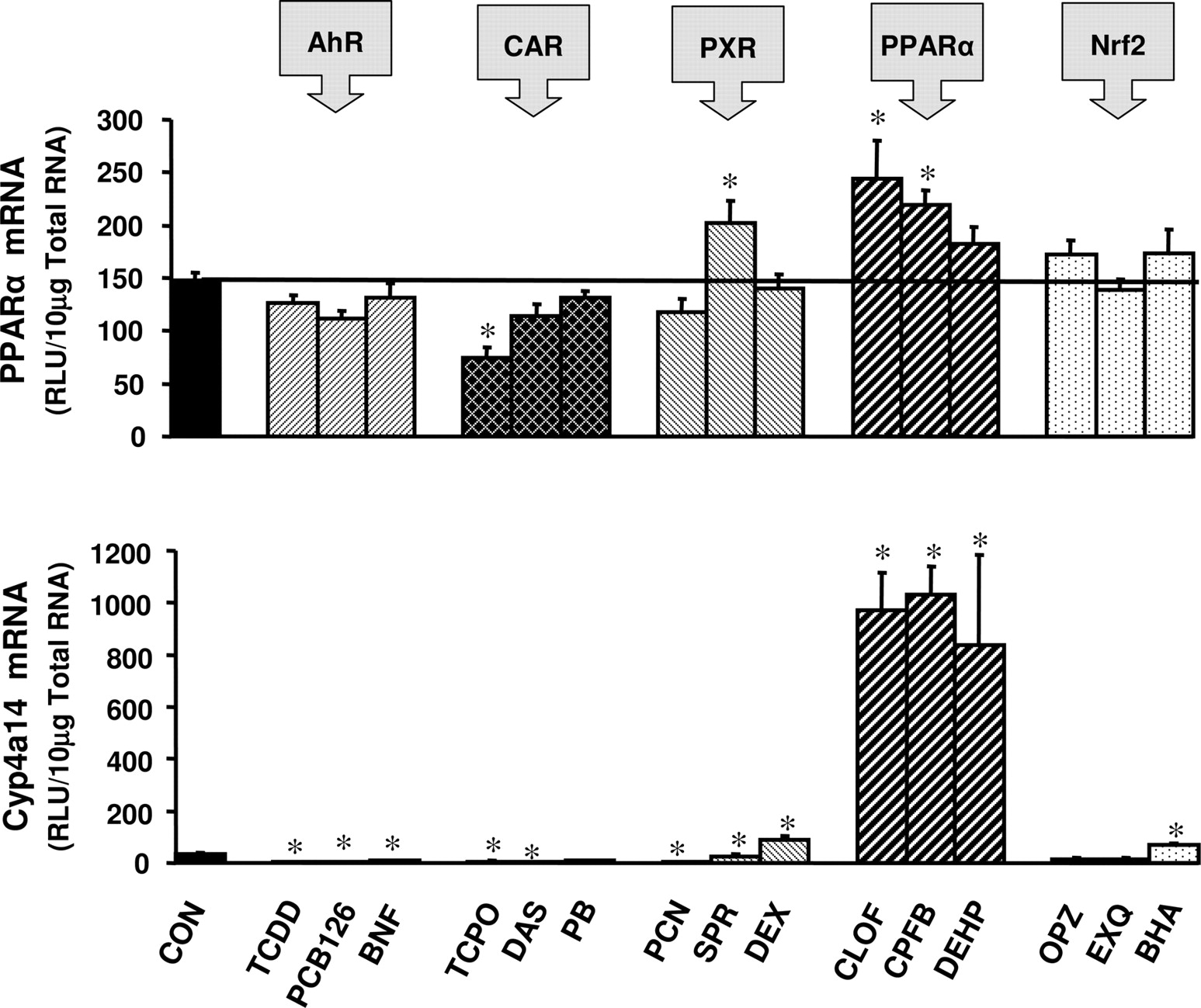
Hepatic expression of PPARα was reduced 50% after treatment with the CAR activator TCPOBOP (Fig. 5). PPARα expression was slightly induced by the PXR activator SPR and by the PPARα activators CLOF and CPFB (Fig. 5). Expression of PPARα was not altered in response to any AhR or Nrf2 activator. Hepatic expression of the PPARα marker gene Cyp4a14 was down-regulated by treatment with activators of AhR and CAR (Fig. 5). The PXR activators PCN and SPR decreased expression of Cyp4a14, whereas DEX induced Cyp4a14 expression 170% (Fig. 5). The PPARα activators CLOF, CPFB, and DEHP induced Cyp4a14 expression 29, 30, and 25 fold, respectively (Fig. 5). Cyp4a14 was also modestly induced by the Nrf2 activator BHA.
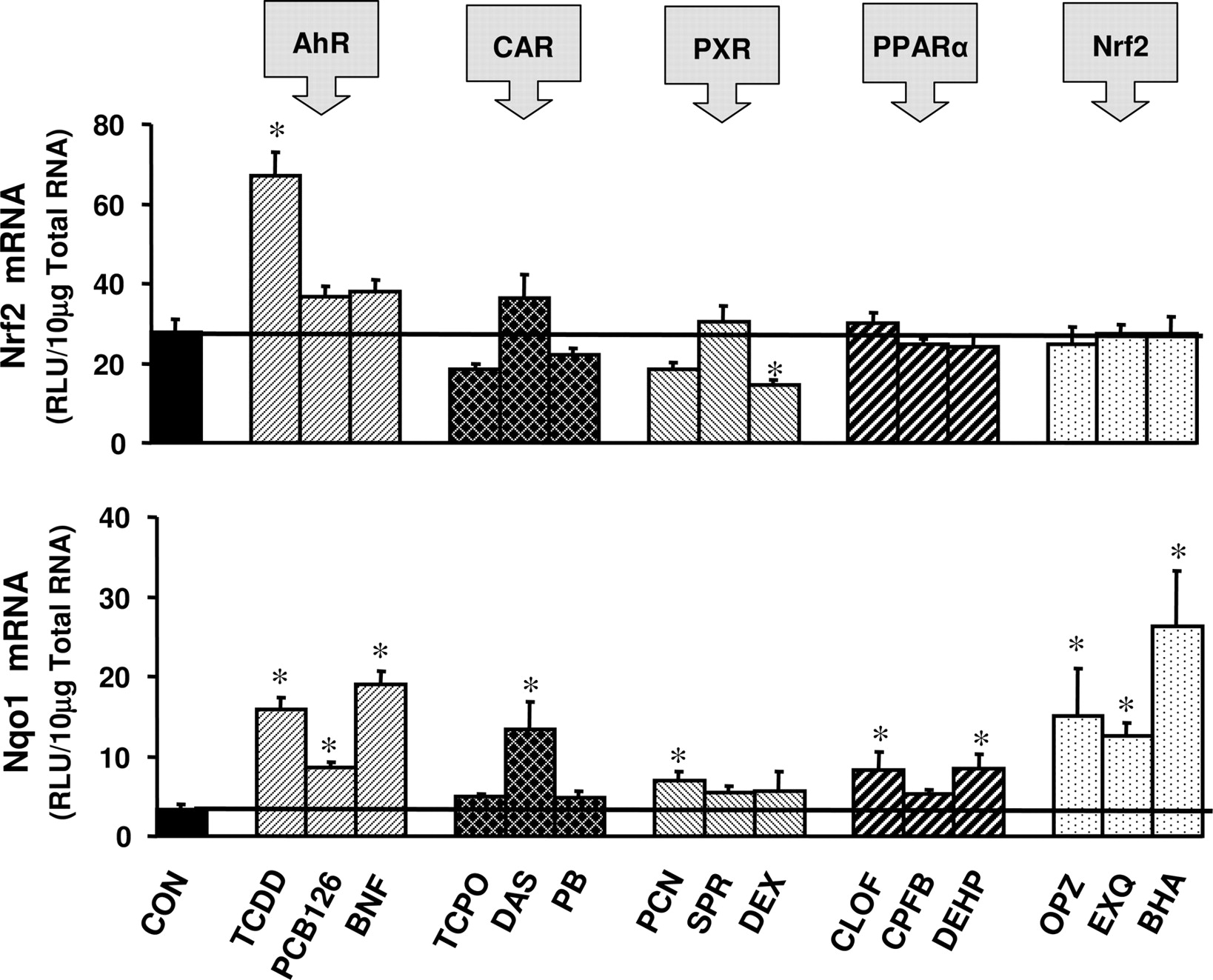
Nrf2 expression was induced 230% by the AhR activator TCDD (Fig. 6). The PXR activator DEX reduced expression of Nrf2 by 47% (Fig. 6). Activators of CAR, PPARα, and Nrf2 did not alter expression of Nrf2 mRNA. Expression of the Nrf2 target gene Nqo1 in mouse liver was induced 370, 160, and 470% by the AhR activators TCDD, PCB-126, and BNF, respectively (Fig. 6). Hepatic expression of Nqo1 was induced 300% by the CAR activator DAS and 110% by the PXR activator PCN (Fig. 6). Nqo1 expression was induced 150% by the PPARα activators CLOF and DEHP. Furthermore, Nqo1 induction was observed in response to the Nrf2 activators OPZ, EXQ, and BHA (350, 280, and 680%, respectively) (Fig. 6).
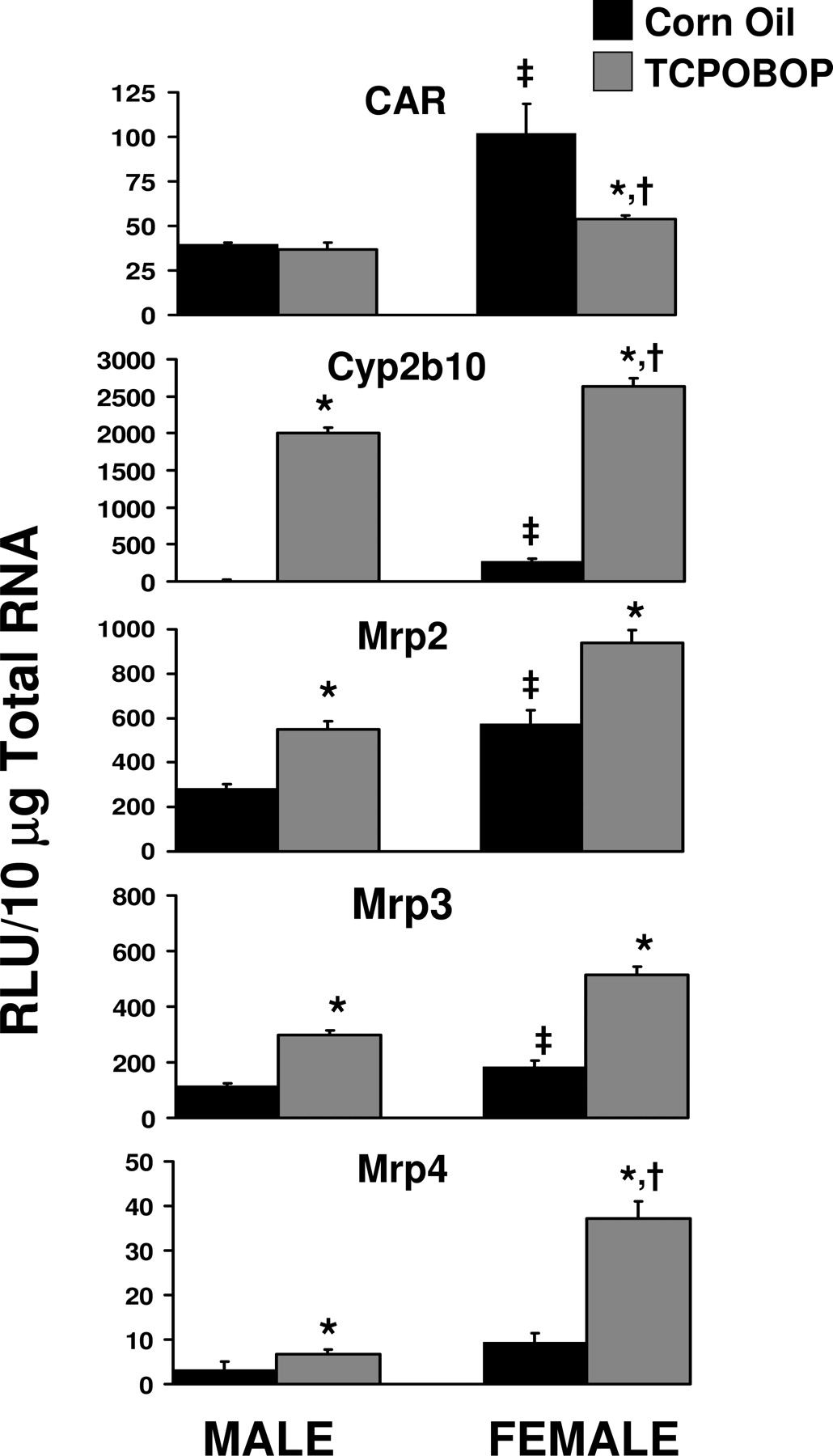
Gender-Divergent Induction of CAR Target Genes in Liver by the CAR Ligand TCPOBOP. Basal expression of CAR mRNA was significantly higher (260%) in livers of female C57BL/6 mice treated with corn oil than in male livers (Fig. 7). CAR expression was not inducible by TCPOBOP treatment in males or females. TCPOBOP reduced basal expression of CAR in females. The CAR target gene Cyp2b10 exhibited 14-fold higher basal expression in females than in males. Cyp2b10 was highly induced by TCPOBOP in male and female mouse liver; however, Cyp2b10 was induced to a higher level in females. The fold induction of Cyp2b10 by TCPOBOP in males was greater than that observed in females. Basal expression of Mrp2 and Mrp3 in female liver was double that observed in males, and this same trend was observed for Mrp4 (although not statistically significant) (Fig. 7). The fold induction of Mrp2 and Mrp3 by TCPOBOP did not exhibit a gender difference, although the magnitude of the expression of these transporters was induced to a greater extent in females than in males. Mrp4 induction by TCPOBOP in female liver was twice than observed in males.

Tissue distribution of AhR, CAR, PXR, PPARα, and Nrf2 mRNA expression in C57BL/6 mice. Tissues were harvested from 8-week-old male and female C57BL/6 mice, as described under Materials and Methods. Total RNA isolates from mouse tissues (n = 5) were analyzed for gene expression using the bDNA assay. Data represent means ± S.E.M. *, statistically significant difference between male and female mice (p < 0.05). RLU, relative light units.
Hepatic Induction of CAR and Cyp2b10 mRNA by AhR Ligands in Mice. CAR expression in mouse liver was induced 360% by treatment with TCDD for 4 days (Fig. 8). CAR expression was not induced in liver following a 12-h treatment with TCPOBOP, although a 4-day TCDD treatment followed by a 12-h TCPOBOP treatment induced CAR expression by 250% over corn oil control and by 320% compared with treatment with TCPOBOP alone. Cyp2b10 expression was induced 330% by treatment with TCDD alone and was markedly induced (95-fold) by TCPOBOP alone (Fig. 8). Pretreatment with TCDD for 4 days followed by a 12-h TCPOBOP treatment induced Cyp2b10 by 180 fold, nearly twice the induction observed in animals treated with TCPOBOP alone.
Hepatic Induction of Mrp2, Mrp3, and Mrp4 in Mice. Mrp2 mRNA expression was not significantly induced by TCDD treatment alone; however, Mrp2 was induced 190% by TCPOBOP treatment alone (Fig. 9). Pretreatment with TCDD for 4 days, followed by a 12-h TCPOBOP treatment induced Mrp2 expression by 285% over control, by 190% over TCDD alone, and by 150% over TCPOBOP alone (Fig. 9). Mrp3 mRNA expression was induced 265% over corn oil controls in mice pretreated with TCDD and subsequently given TCPOBOP, but not by treatment with either compound alone (Fig. 9). Mrp4 mRNA expression was not induced by treatment with TCDD (4 days) or TCPOBOP (12 h) alone. Although a statistically significant reduction in Mrp4 mRNA expression was also observed after 12 h of TCDD treatment, the extremely low level of expression of Mrp4 at levels near the limit of detection call into question any potential biological significance for this observation. However, in mice pretreated with TCDD, followed by TCPOBOP treatment, Mrp4 mRNA was induced to a significant extent relative to those animals administered corn oil vehicle or either compound alone.

Induction of AhR and Cyp1a1 mRNA expression in male C57BL/6 mouse liver by microsomal enzyme inducer compounds. Animals were treated with inducer agents as described in Table 1. Gene expression from livers of five mice was determined using the bDNA assay. Data represent means ± S.E.M. *, statistically significant difference between treated and control mice (p < 0.05). RLU, relative light units.
Discussion
The present studies examined tissue distribution, gender-divergent expression, and hepatic induction of AhR, CAR, PXR, PPARα, and Nrf2 mRNA in mice. Gender-divergent expression and hepatic induction of CAR by TCDD produced concomitant alterations in expression of its associated target genes. Data from the present studies show that in some cases, AhR, CAR, PXR, PPARα, and Nrf2 were modestly induced by chemical activators that markedly induce expression of their associated target genes. Collectively, these data indicate that inducible expression of several of these transcription factors may play an important role in induction of their associated target genes.
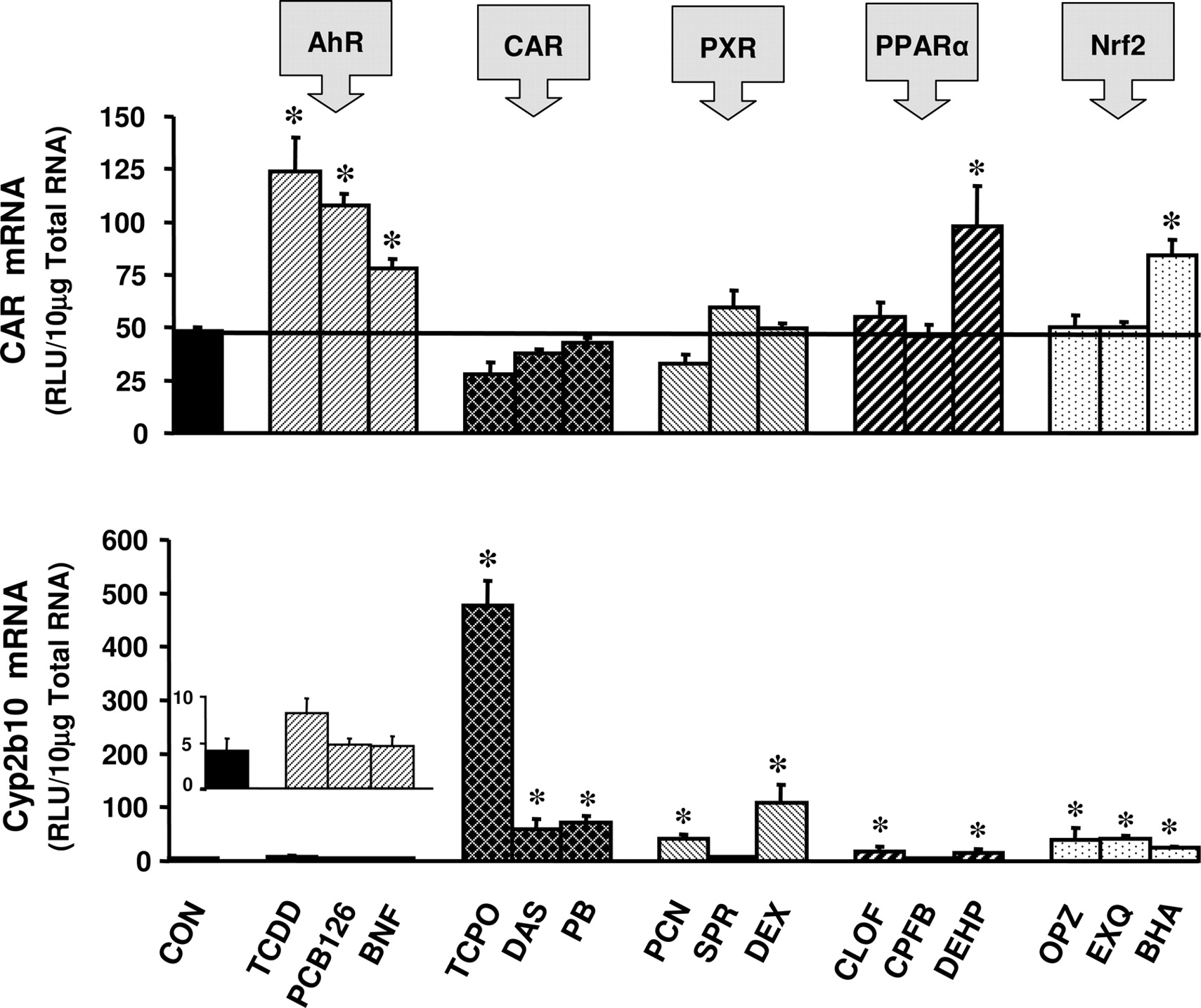
Induction of CAR and Cyp2b10 mRNA expression in male C57BL/6 mouse liver by microsomal enzyme inducer compounds. Animals were treated with inducer agents as described in Table 1. Gene expression from livers of five mice was determined using the bDNA assay. Data represent means ± S.E.M. *, statistically significant difference between treated and control mice (p < 0.05). RLU, relative light units.
AhR, CAR, PXR, PPARα, and Nrf2 were ubiquitously expressed in the observed tissues and typically exhibited higher expression levels in extrahepatic tissues. The tissue expression patterns of these transcription factors coincided with those reported in other mouse studies appearing in the literature (Kliewer et al., 1994, 1998; Li et al., 1994; Moi et al., 1994; Choi et al., 1997; McMahon et al., 2001; Bookout et al., 2006). The present study is the first to quantitatively compare expression of Nrf2 in various segments of the small intestine, the large intestine, and in several portions of the female reproductive tract. Nrf2 expression was highest in stomach and intestinal tract, consistent with earlier studies noting that the highest expression of Nrf2 is in extrahepatic tissues (Moi et al., 1994; McMahon et al., 2001).

Induction of PXR and Cyp3a11 mRNA expression in male C57BL/6 mouse liver by microsomal enzyme inducer compounds. Animals were treated with inducer agents as described in Table 1. Gene expression from livers of five mice was determined using the bDNA assay. Data represent means ± S.E.M. *, statistically significant difference between treated and control mice (p < 0.05). RLU, relative light units.
In the present studies, xenobiotic induction of AhR, CAR, PXR, PPARα, and Nrf2 was evaluated in the liver, along with induction of their associated target genes, Cyp1a1, Cyp2b10, Cyp3a11, Cyp4a14, and Nqo1, respectively. The xenobiotic compounds used in this study (Table 1) are classic MEIs thought to induce these target genes through the above-mentioned transcription factors. The specificity of this induction was evaluated by examining the induction of these transcription factors by MEIs and by observing corresponding induction of their associated target genes. In most cases, transcription factor induction was modest, whereas marked induction of their associated marker genes was observed. Xenobiotic inducers exhibited selective induction of the target genes and minimal selectivity in induction of their associated transcription factors.

Induction of PPARα and Cyp4a14 mRNA expression in male C57BL/6 mouse liver by microsomal enzyme inducer compounds. Animals were treated with inducer agents as described in Table 1. Gene expression from livers of five mice was determined using the bDNA assay. Data represent means ± S.E.M. *, statistically significant difference between treated and control mice (p < 0.05). RLU, relative light units.
AhR ligands were potent inducers of Cyp1a1 mRNA expression and also modestly induced expression of the Nrf2 marker gene Nqo1. Induction of Nqo1 expression by AhR ligands requires Cyp1a1 expression and is thus likely to involve the AhR (Marchand et al., 2004). Furthermore, Cyp1a1 oxidative metabolites may initiate ARE-dependent activation of Nqo1 through a Nrf2-dependent signaling pathway, as TCDD induction of Nqo1 expression requires the presence of Nrf2 (Ma et al., 2004).

Induction of Nrf2 and Nqo1 mRNA expression in male C57BL/6 mouse liver by microsomal enzyme inducer compounds. Animals were treated with inducer agents as described in Table 1. Gene expression from livers of five mice was determined using the bDNA assay. Data represent means ± S.E.M. *, statistically significant difference between treated and control mice (p < 0.05). RLU, relative light units.
Interestingly, the Nrf2 activator BHA significantly induced expression of AhR mRNA, indicating a further relationship between these two important transcription factor pathways. AhR mRNA expression was modestly induced in response to the AhR activator BNF; however, AhR was not induced significantly by other AhR activators or in response to more than one activator from any other class of microsomal enzyme inducer used in the present studies (Fig. 2).
CAR and PXR activators markedly enhanced hepatic Cyp2b10 and Cyp3a11 mRNA expression, as these transcription factors bind to both the phenobarbital response element modules and PXR response element (Wang and Negishi, 2003). Induction of Cyp2b10 mRNA expression by PPARα and Nrf2 activators in the present study may also implicate these transcription factors in Cyp2b regulation. In the present studies, CAR mRNA expression was not induced by CAR activators (Fig. 3); however, expression of CAR in the liver was induced by all three AhR ligands, and TCDD modestly induced basal expression of the CAR target gene Cyp2b10. Furthermore, induction of Cyp2b10 and Mrp2, 3, and 4 mRNA expression by TCPOBOP was also enhanced after CAR induction by TCDD (Figs. 8 and 9). The present studies thus present the novel finding that AhR can regulate expression of CAR and its downstream target genes. Because of the important role played by CAR in regulation of xenobiotic metabolism and transport, induction of CAR by TCDD implicates AhR as an important regulator of drug metabolism and disposition.
In the present studies, hepatic expression of PXR was modestly inducible by the PXR ligand SPR and by the Nrf2 activator BHA. PXR expression was doubled by the PPARα agonists CLOF and DEHP in liver, findings that may merit further investigation. Only PPARα ligands markedly induced Cyp4a14 expression in liver, thus showing the high degree of selectivity of these ligands as Cyp4a14 inducers. Hepatic expression of PPARα mRNA was also moderately induced by the PPARα ligands CLOF and CPFB, making this the only transcription factor in the present study to be induced by its activator compounds.
Nrf2 activators induced hepatic Nqo1 expression up to 3-fold, consistent with intestinal induction of Nqo1 by the Nrf2 activators BHA and OTZ (McMahon et al., 2001). Nqo1 was markedly induced in liver by AhR ligands, and Nqo1 induction was modest in response to CAR, PXR, and PPARα activators. The present data illustrate that marked induction of Nrf2 is not required for Nqo1 induction in vivo. Nrf2 expression in liver was not appreciably inducible by CAR, PXR, PPARα, or Nrf2 activators; however, Nrf2 expression was more than doubled by treatment with the AhR ligand TCDD. In rats, Nqo1 induction by AhR activators depends on CYP1A1 expression, thus implicating the AhR as a mediator of Nqo1 induction (Marchand et al., 2004). This induction may also be mediated via ARE-dependent Nqo1 activation by Cyp1a1 oxidative metabolites as evidenced by Nrf2-dependent induction of Nqo1 by TCDD (Ma et al., 2004).
The present studies and numerous reports in the literature thus illustrate the complex nature of the interaction between xenobiotic activator compounds and transcription factors. Furthermore, the present studies also demonstrate the nonselectivity of these classic xenobiotic activator compounds. In addition to tissue-specific variations in transcription factor expression and xenobiotic-dependent induction of these genes and their associated target genes, a number of these transcription factors also exhibited gender-divergent expression in certain tissues. Most notably, CAR expression in female liver was found in the present study to be twice that in male liver.
A gender difference in mouse hepatic CAR expression has been noted previously (Kawamoto et al., 2000; Ledda-Columbano et al., 2003). Furthermore, basal and inducible expression of Cyp2b10 mRNA in female liver is greater than that observed in males (Honkakoski et al., 1992; Ledda-Columbano et al., 2003). In vitro studies demonstrated that estrogen stimulates activation of mouse CAR (Kawamoto et al., 2000). Further evidence that genderdivergent responses to CAR activators in mice are mediated by sex hormones is provided by the suppression of TCPOBOP-mediated CAR and Cyp2b10 induction after treatment with androstenol (Ledda-Columbano et al., 2003). In the present study, CAR was not induced by TCPOBOP in the livers of male mice, a finding that was reproduced in several independent experiments (Figs. 2, 7, and 8), thus indicating an apparent strain difference between the C57BL6/J mouse used in the present study and the CD-1 mouse, in which CAR was induced by TCPOBOP in both males and females (Ledda-Columbano et al., 2003). Furthermore, in the present studies, CAR expression was reduced after 4 days of TCPOBOP administration, indicating another possible strain difference between C57BL6/J and CD-1 mice.
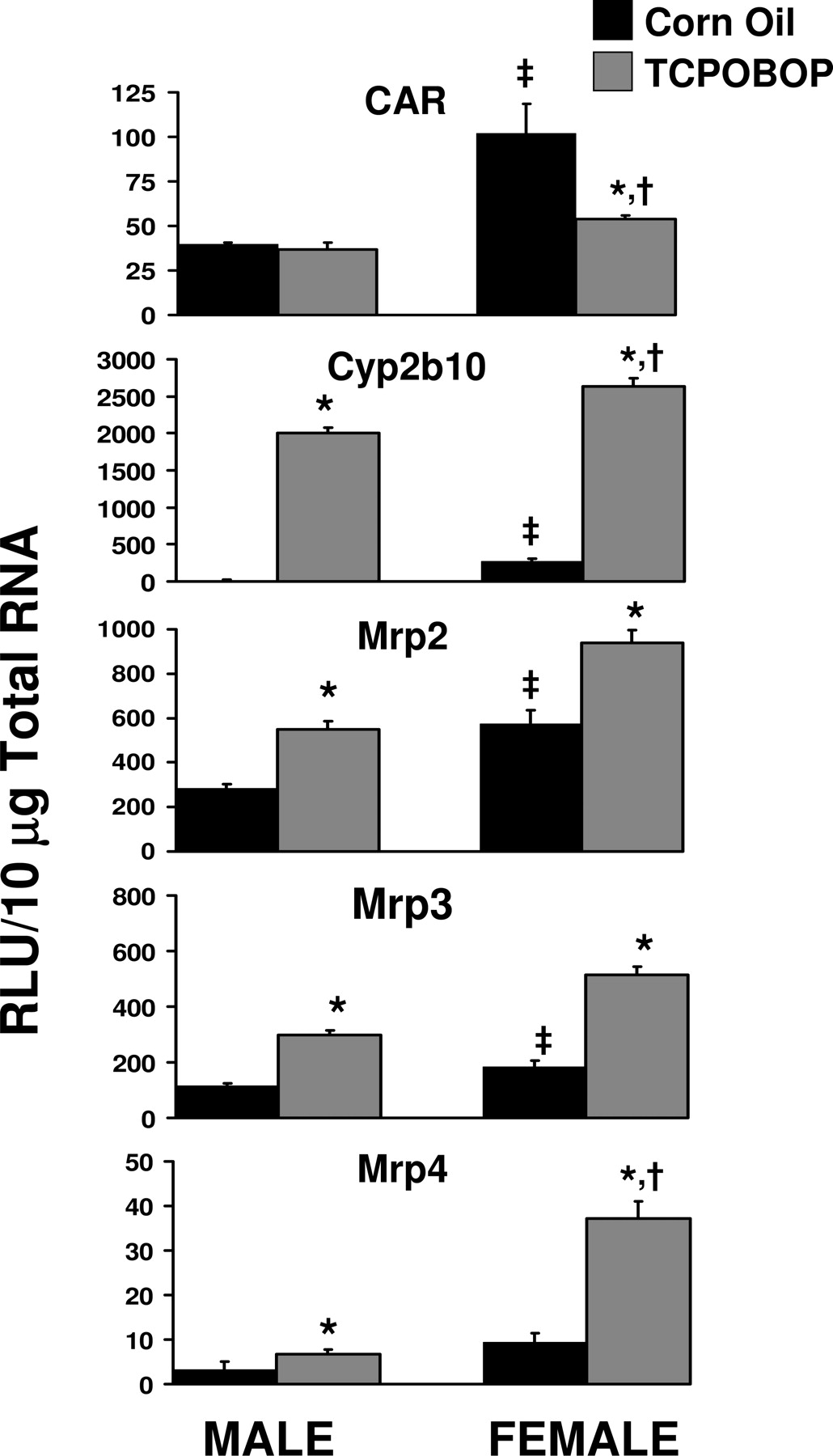
Gender differences in expression of CAR, Cyp2b10, Mrp2, Mrp3, and Mrp4 in C57BL/6 mouse liver and gender-divergent responses to the CAR ligand TCPOBOP. Male and female mice were treated with corn oil or TCPOBOP for 4 days, as described in Table 1. Gene expression from livers of five mice was determined using the bDNA assay. Data represent means ± S.E.M. Statistical significance was evaluated using Student's t test and significant differences (p < 0.05) are indicated as follows: ‡, between male and female; *, between corn oil and TCPOBOP treatment; †, ratio of corn oil to TCPOBOP treated in males versus females. RLU, relative light units.
In addition to regulation of Cyp2b10, CAR has been implicated in xenobiotic induction of the Mrp transporters Mrp2, 3, and 4 (Cherrington et al., 2002; Assem et al., 2004; Maher et al., 2005a). The present studies illustrate that basal expression of Mrp2, 3, and 4 is female-predominant. Although relative expressions of Mrp2, 3, and 4 were higher in TCPOBOP-treated female mice than in TCPOBOP-treated male mice, only Mrp4 was induced to a greater extent in females than in males. Taken together, these data illustrate that elevated CAR expression in females produces elevated basal and inducible expression of the CAR target genes Cyp2b10, Mrp2, Mrp3, and Mrp4.
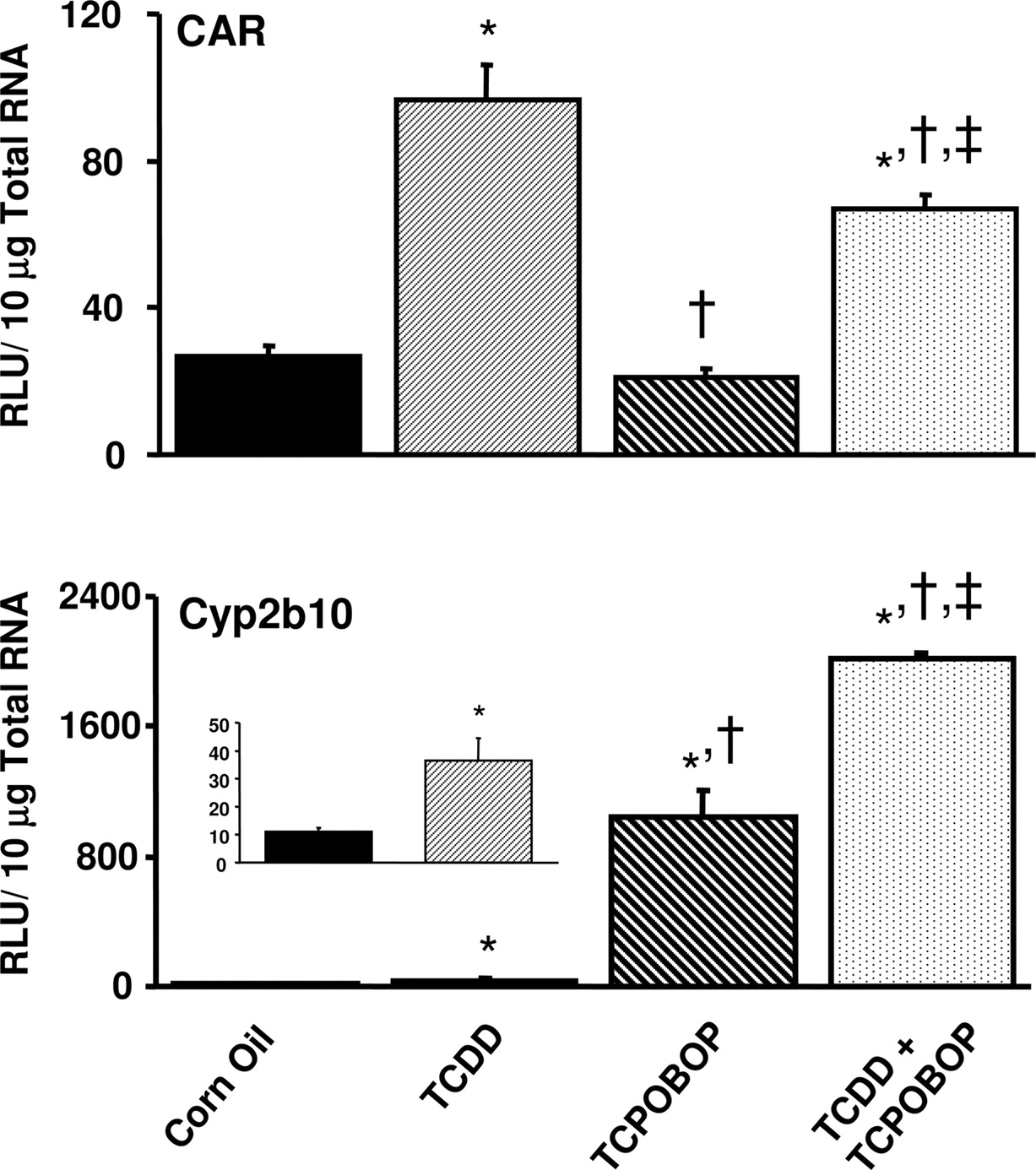
Induction of CAR and Cyp2b10 mRNA in male C57BL/6 mouse liver by TCCD, TCPOBOP, or combined treatment of TCDD and TCPOBOP. Animals were treated with 1) 5 ml/kg/day corn oil for 4 days, 2) 37 μg/kg/day TCDD for 4 days, 3) 5 ml/kg/day corn oil for 4 days followed by 3 mg/kg TCPOBOP for 12 h, or 4) 37 μg/kg/day TCDD for 4 days followed by a 12-h 3 mg/kg TCPOBOP treatment (see details under Materials and Methods). Gene expression (n = 5) was analyzed using the bDNA assay (see Materials and Methods). Data represent means ± S.E.M. Statistically significant differences (p < 0.05) are indicated as follows: *, significantly different from corn oil treatment; †, significantly different from TCDD treatment; ‡, significantly different from TCPOBOP treatment. RLU, relative light units.
Despite marked induction of target genes by activators of AhR, CAR, PXR, PPARα, and Nrf2, these transcription factors were modestly inducible or not induced by their associated chemical activators. With the exception of CAR induction by AhR and its effect on CAR target genes, those compounds that significantly induce any of the selected transcription factors tended to have little or no effect on expression of their associated target genes. In addition, the present studies show that these classic inducer compounds activate their targeted nuclear receptor pathways and that they may also selectively activate other transcription factor pathways and their associated target genes.
Collectively, the present data provide important insight into the tissue-specific expression, gender-divergent expression, and xenobiotic regulation of AhR, CAR, PXR, PPARα, and Nrf2 mRNA and of their associated target genes. The present studies provide useful information that may enhance the understanding of the role of both gender and xenobiotics in the regulation of xenobiotic-metabolizing enzymes and transporters. Understanding this gene regulation is critical, as dysregulation of transporters and drug metabolizing enzymes can produce significant variations in drug absorption, distribution, metabolism, and elimination and can thus have an adverse impact on the clinical outcome of drug treatment.

Induction of Mrp2, Mrp3, and Mrp4 transporter mRNA in C57BL/6 mouse liver by TCCD, TCPOBOP, or combined treatment of TCDD and TCPOBOP. Animals were treated with 1) 5 ml/kg/day corn oil for 4 days, 2) 37 μg/kg/day TCDD for 4 days, 3) 5 ml/kg/day corn oil for 4 days followed by 3 mg/kg TCPOBOP for 12 h, or 4) 37 μg/kg/day TCDD for 4 days followed by a 12-h 3 mg/kg TCPOBOP treatment (see details under Materials and Methods). Gene expression (n = 5) was analyzed using the bDNA assay (see Materials and Methods). Data represent means ± S.E.M. Statistically significant differences (p < 0.05) are indicated as follows: *, significantly different from corn oil treatment; †, significantly different from TCDD treatment; ‡, significantly different from TCPOBOP treatment. RLU, relative light units.
Acknowledgments
We thank Dr. David Buckley for assistance with laboratory work on this study. We also thank the members of the Klaassen laboratory for thorough review of the manuscript.
Footnotes
-
Funding for this project was provided by National Institutes of Health Grants ES 07079, ES 09649, and ES 09716.
-
doi:10.1124/dmd.107.015974.
-
ABBREVIATIONS: AhR, aryl hydrocarbon receptor; CAR, constitutive androstane receptor; PXR, pregnane X receptor; PPARα, peroxisome proliferator activated receptor-α; Nrf2, nuclear factor-E2-related factor 2; MEI, microsomal enzyme inducer; P450, cytochrome P450; Nqo1, NAD(P)H quinine oxidoreductase 1; Ugt, UDP glucuronosyl transferase; Gst, glutathione S-transferase; PB, phenobarbital; TCPOBOP, 1,4-bis-[2-(3,5-dichloropyridyloxy)]benzene; Mrp, multidrug resistance-associated protein; PCN, pregnenolone-16α-carbonitrile; Wy-14,643, pirinixic acid, 4-chloro-6-(2,3-xylidino)-2-pyrimidinyl)thioacetic acid; ARE, antioxidant response element; TCDD, 2,3,7,8-tetrachlorodibenzo-p-dioxin; PCB-126; polychlorinated biphenyl 126; bDNA, branched DNA; BNF, β-naphthoflavone; SPR, spironolactone; CLOF, clofibrate; BHA, butylated hydroxyanisole; DEHP, di-(2-diethylhexyl)phthalate; DAS, diallyl sulfide; DEX, dexamethasone; OPZ, oltipraz; EXQ, ethoxyquin.
- Received March 30, 2007.
- Accepted July 9, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}