


Abstract
The purpose of the present study was to evaluate the effects of bovine serum albumin (BSA) and essentially fatty acid-free BSA (BSA-FAF) on the biliary clearance of compounds in sandwich-cultured rat hepatocytes. Unbound fraction, biliary excretion index (BEI), and unbound intrinsic biliary clearance (intrinsic  ) were determined for digoxin, pravastatin, and taurocholate in the absence or presence of BSA or BSA-FAF. BSA had little effect on the BEI or intrinsic
) were determined for digoxin, pravastatin, and taurocholate in the absence or presence of BSA or BSA-FAF. BSA had little effect on the BEI or intrinsic  of these compounds. Surprisingly, BSA-FAF decreased both BEI and intrinsic
of these compounds. Surprisingly, BSA-FAF decreased both BEI and intrinsic  for digoxin and pravastatin, which represent low and moderately bound compounds, respectively. The BEI and intrinsic
for digoxin and pravastatin, which represent low and moderately bound compounds, respectively. The BEI and intrinsic  of taurocholate, a highly bound compound, were not altered significantly by BSA-FAF. Neither BSA nor BSA-FAF had a discernable effect on the bile canalicular networks based on carboxydichlorofluorescein retention. Neither the addition of physiological concentrations of calcium nor the addition of fatty acids to BSA-FAF was able to restore the BEI or intrinsic
of taurocholate, a highly bound compound, were not altered significantly by BSA-FAF. Neither BSA nor BSA-FAF had a discernable effect on the bile canalicular networks based on carboxydichlorofluorescein retention. Neither the addition of physiological concentrations of calcium nor the addition of fatty acids to BSA-FAF was able to restore the BEI or intrinsic  of the model compounds to similar values in the absence or presence of BSA. Careful consideration is warranted when selecting the type of BSA for addition to in vitro systems such as sandwich-cultured rat hepatocytes.
of the model compounds to similar values in the absence or presence of BSA. Careful consideration is warranted when selecting the type of BSA for addition to in vitro systems such as sandwich-cultured rat hepatocytes.
In vitro models are used widely to investigate hepatic metabolism and transport. Sandwich-cultured rat hepatocytes are one such model system. Culturing primary hepatocytes between two layers of extracellular matrix, which resembles basement membranes in vivo to which cells attach, allows the cells to maintain many in vivo structural and functional characteristics. Apical (canalicular) and basolateral (sinusoidal) membrane domains are re-established following cellular repolarization and sealing of tight junction complexes; liver-specific proteins are expressed; and intact functional bile canalicular networks are formed (LeCluyse et al., 1994, 2000; Liu et al., 1998, 1999b; Hoffmaster et al., 2004). This model has been established as an in vitro method to predict in vivo biliary clearance of drug candidates (Liu et al., 1999a; Fukuda et al., 2008). It is a useful in vitro model to study hepatic uptake and biliary excretion mechanisms simultaneously. Data from medium, cell, and bile compartments of sandwich-cultured hepatocytes can be used for pharmacokinetic modeling of hepatobiliary drug disposition (Turncliff et al., 2006). Sandwich-cultured rat hepatocytes also are used to predict drug and xenobiotic interactions that may occur in vivo at the level of hepatic transporters (Annaert and Brouwer, 2005; Kemp et al., 2005; McRae et al., 2006).
Albumin and α1-acid glycoprotein are the primary proteins responsible for binding low-molecular-weight compounds in plasma. As a general rule, cations bind to α1-acid glycoprotein, whereas anions bind primarily to albumin (Peters, 1996). Albumin is the most abundant protein present in plasma and often is added to in vitro systems (Aungst et al., 2000; Yamashita et al., 2000; Krishna et al., 2001; Cross et al., 2003; Neuhoff et al., 2006) to prevent precipitation of water-insoluble drugs and to more closely mimic physiologic conditions (Katneni et al., 2008). Bovine serum albumin (BSA) is used commonly in vitro as a replacement for albumin from other species (Saha and Kou, 2002; Taub et al., 2002; Neuhoff et al., 2006).
A major determinant of drug disposition in vivo is binding to plasma proteins (Otagiri, 2005). Many drug candidates are extensively protein-bound, tend to be hydrophobic and of limited solubility, and are surface-adsorptive, thus leading to experimental challenges in the drug development process (Artursson, 1990; Wilson et al., 1990). The ability to deliver a compound to, and stabilize a compound in, in vitro systems can be enhanced by the addition of protein. However, such manipulation may significantly alter pharmacokinetic/pharmacodynamic behavior (Wiseman and Nelson, 1964; Bennhold, 1965; Fisher et al., 1999; Liu et al., 2005; Neuhoff et al., 2006). Metabolic studies using human liver microsomes, fresh or cryopreserved hepatocytes, or transfected systems underpredict in vivo hepatic clearance and metabolic inhibition potential (for review, see Rowland et al., 2007). Studies investigating the effect of BSA on hepatic metabolism by UDP-glucuronosyltransferase (UGT) 2B7, UGT1A9, and cytochrome P450 2C9 in these systems suggested an increase in in vitro intrinsic clearance in the presence of BSA (for review, see Rowland et al., 2007, 2008a,b). Similar results were observed for UGT2B7 despite the preparation of BSA used [“crude,” essentially fatty acid-free (BSA-FAF), essentially globulin-free, or essentially fatty acid-free and globulin-free] (Rowland et al., 2007). It was hypothesized that BSA, despite the preparation type, sequesters fatty acids that are inhibitory to the activity of metabolic enzymes (Rowland et al., 2007); however, not all metabolic enzymes (UGT1A1, UGT1A6) are inhibited by fatty acids (Rowland et al., 2008b).
Little work evaluating the effect of BSA on transport proteins and the hepatobiliary disposition of compounds has been published. Previous investigations using transfected cells have focused on transport proteins involved in the uptake of compounds into hepatocytes (Shi et al., 1995; Cui et al., 2001; Hata et al., 2003). The primary objective of the present study was to characterize the effect of BSA on the biliary excretion index (BEI) and biliary clearance of compounds in sandwich-cultured rat hepatocytes. Because several preparations of albumin are available commercially, we investigated the effects of BSA and BSA-FAF on these parameters. The compounds selected for investigation (digoxin, pravastatin, and taurocholate) span a range of unbound fractions: digoxin (fu ∼ 0.7; Lacy et al., 2002), pravastatin (fu ∼ 0.5; Hatanaka, 2000), and taurocholate (fu ∼ 0.15; Forker and Luxon, 1981; Hung et al., 2005). Additional studies were designed to characterize 1) the effect of albumin preparations on bile canalicular networks and 2) the effect of long-chain fatty acids, which are present in BSA preparations, on BEI and biliary clearance.
Materials and Methods
Materials. Collagenase (type 1, class 1) was obtained from Worthington Biochemicals (Lakewood, NJ). Dulbecco's modified Eagle's medium (DMEM, no phenol red), insulin, minimal essential medium (MEM) nonessential amino acids solution (100×), l-glutamine, penicillin G-streptomycin solution, and 5 (and 6)-carboxy-2′,7′-dichlorofluorescein diacetate (CDFDA) were purchased from Invitrogen (Carlsbad, CA). Fetal bovine serum, sodium taurocholate, digoxin, Triton X-100, dexamethasone, methanol, Hanks' balanced salts modified with (H-1387) or without (H-4891) calcium chloride, BSA, BSA-FAF (A-8806), linoleic acid (18:2n-6), linolenic acid (18:3n-3), oleic acid (18:1n-9), palmitic acid (16:0), palmitoleic acid (16:1n-7), and stearic acid (18:0) were obtained from Sigma-Aldrich (St. Louis, MO). BioCoat collagen I plates, Matrigel basement membrane matrix, and ITS+ (insulin/transferrin/selenium) culture supplement were purchased from BD Biosciences Discovery Labware (Bedford, MA). Pravastatin and d6-rosuvastatin were purchased from Toronto Research Chemicals (North York, ON, Canada). [3H]Taurocholate (5 Ci/mmol, >97% purity) and [3H]digoxin (40 Ci/mmol, >97% purity) were obtained from PerkinElmer Life and Analytical Sciences (Boston, MA). Bio-Safe II liquid scintillation mixture was obtained from Research Products International (Mt. Prospect, IL). Centrifree micropartition devices were obtained from Millipore (Billerica, MA). Bicinchoninic acid (BCA) protein assay reagents and BSA for the protein assay standard were purchased from Pierce Chemical Co. (Rockford, IL). All the other chemicals and reagents were of analytical grade and available from commercial sources.
Animals. Male Wistar rats (200–325 g), obtained from Charles River Laboratories, Inc. (Raleigh, NC), served as liver donors for hepatocyte isolation. Animals were maintained in a controlled environment with a 12-h light/dark cycle and had free access to water and food before surgery. All the animal procedures were compliant with the guidelines of The University of North Carolina at Chapel Hill Institutional Animal Care and Use Committee.
Isolation and In Vitro Culture of Primary Rat Hepatocytes. Hepatocytes were isolated from male Wistar rats using a two-step collagenase perfusion, as described previously (LeCluyse et al., 1996). Cell viability, determined by trypan blue exclusion, was >88%. Hepatocytes were seeded (∼1.75 × 106 cells/well) in six-well BioCoat plates in DMEM supplemented with 2 mM l-glutamine, 1% (v/v) MEM nonessential amino acids, 100 units of penicillin G sodium/ml, 100 μg of streptomycin sulfate/ml, 1 μM dexamethasone, 5% (v/v) fetal bovine serum, and 10 μM insulin (day 0 of culture) and cultured in a humidified incubator (95% O2, 5% CO2) at 37°C. To achieve a sandwich configuration 24 h postplating (day 1 of culture), hepatocytes were overlaid with Matrigel basement membrane matrix (0.25 mg/ml) in 2 ml/well cold serum-free DMEM containing 2 mM l-glutamine, 1% (v/v) MEM nonessential amino acids, 100 units of penicillin G sodium/ml, 100 μg of streptomycin sulfate/ml, 0.1 μM dexamethasone, and 1% (v/v) ITS+. The culture medium was changed every 24 h until experiments were performed on day 4 of culture.
Accumulation Experiments. The method to determine substrate accumulation in sandwich-cultured rat hepatocytes has been described previously (Liu et al., 1999b). Briefly, hepatocytes were rinsed twice with 2 ml of warm Hanks' balanced salt solution (HBSS) containing Ca2+ (standard buffer) or Ca2+-free HBSS and incubated with 2 ml of the same buffer for 10 min at 37°C (to maintain tight junction integrity and bile canalicular networks or disrupt tight junctions and open bile canalicular networks, respectively). The buffer was removed, and the cells were incubated for 10 min at 37°C with 1.5 ml of [3H]taurocholate (1 μM), [3H]digoxin (10 μM), or pravastatin (5 μM) in standard buffer in the presence or absence of 4% (w/v) BSA or BSA-FAF. For experiments in which long-chain fatty acids were added, standard buffer with or without BSA or BSA-FAF remained at room temperature for ∼1 h following the addition of long-chain fatty acids (diluted in dimethyl sulfoxide) to allow binding of the fatty acids to albumin (Peters, 1996) before the addition of pravastatin and subsequent incubation with hepatocytes for 10 min. The fatty acids added were linoleic acid (5.48 μM), linolenic acid (0.20 μM), oleic acid (6.94 μM), palmitic acid (9.38 μM), palmitoleic acid (0.76 μM), and stearic acid (7 μM). Hepatocytes were rinsed vigorously three times with 2 ml of ice-cold standard buffer following the incubation. For rinsing of [3H]digoxin-treated hepatocytes, 10 μM unlabeled digoxin was added to the rinsing buffer to reduce nonspecific binding as described previously (Annaert et al., 2001). Additional digoxin accumulation studies were performed with or without 1 mM Ca2+ (calcium chloride) added to the digoxin-containing incubation solution. Taurocholate- and digoxin-treated hepatocytes were lysed with 1 ml of 0.5% (v/v) Triton X-100 in phosphate-buffered saline by placing plates on an orbital shaker for a minimum of 20 min at room temperature. The samples were analyzed by liquid scintillation spectroscopy in a Packard Tri-Carb scintillation counter (PerkinElmer Life and Analytical Sciences). Pravastatin-treated hepatocytes were lysed with 1 ml of 70% (v/v) ice-cold methanol/water, scraped off the plates, and sonicated for 20 s with a sonic dismembrator. The samples were analyzed by liquid chromatography with detection by tandem mass spectrometry (LC/MS/MS).
LC/MS/MS Analysis. Hepatocyte lysates from pravastatin accumulation experiments were centrifuged at 12,000g for 5 min at 4°C. The resulting supernatants were diluted 1:6 with water and methanol containing the internal standard (d6-rosuvastatin). Analysis of these samples was performed with a Shimadzu solvent delivery system (Shimadzu Scientific Instruments, Columbia, MD) and a Leap HTC Pal thermostated autosampler (LEAP Technologies, Carrboro, NC) connected to an Applied Biosystems API 4000 triple quadrupole mass spectrometer with a TurboSpray ion source (Applied Biosystems, Foster City, CA). Tuning, operation, integration, and data analysis were performed in positive mode using multiple reaction monitoring (Analyst software version 1.4.1; Applied Biosystems). Ten microliters of sample was injected onto an Aquasil 5-μm C18 column (2.1 mm i.d. × 50 mm; Thermo Electron Corporation, Waltham, MA). The mobile phase consisted of 0.1% (v/v) formic acid diluted in water (buffer A) and methanol containing 0.1% (v/v) formic acid (buffer B). Analytes were eluted with a high-pressure linear gradient program at a flow rate of 0.75 ml/min as follows: 20% buffer B for 0.75 min, increased to 40% buffer B over 0.64 min and held for 1.91 min, rapidly increased to 90% buffer B and held for 0.7 min, and returned rapidly to 20% buffer B. The system was allowed to re-equilibrate for 1 min, with a total run time of 5 min/injection. The entire column effluent was diverted from the TurboSpray ion probe of the mass spectrometer for the first 0.75 min and last 1 min. An eight-point calibration curve (2–1000 nM) was constructed for pravastatin using peak area ratios of analyte and internal standard using the following transitions: pravastatin (447.1→327.4) and d6-rosuvastatin (488.2→264.2). All the points on the curve back-calculated to within ± 15% of the nominal value.
Nonspecific Binding and Protein Determination. All the accumulation data were corrected for nonspecific binding of the relevant substrate to Matrigel-overlaid hepatocyte-free BioCoat plates. The data also were normalized to the protein concentration in each well (lysed with Triton X-100), determined in duplicate aliquots using BCA protein assay reagents as instructed by the manufacturer. BSA, as supplied by the manufacturer, was used as a standard (0.2–1 mg/ml). Because of the incompatibility of methanol with the BCA protein assay, the average protein concentration of samples from one liver preparation lysed with Triton X-100 was used to normalize the accumulation data of samples lysed with methanol from the same liver preparation.
Determination of Protein Binding. The fu of each compound was determined over a range of concentrations: taurocholate (0.1–100 μM), digoxin (0.1–30 μM), and pravastatin (1–10 μM). This determination was not performed to examine the binding kinetics of each compound but rather to ensure that binding was stable and reproducible under the conditions planned for the experiment. Solutions in standard buffer were prepared in the absence or presence of 4% (w/v) BSA or BSA-FAF and incubated at 37°C for 30 min. Solutions containing long-chain fatty acids were prepared in a manner similar to that described under Accumulation Experiments before the addition of pravastatin and incubation for 30 min at 37°C. Media samples from sandwich-cultured rat hepatocyte experiments also were taken to determine the fu of digoxin. Samples (1 ml) were placed in Centrifree micropartition devices and centrifuged at 1500g at 37°C for either 30 s (non-BSA-containing samples) or 4 min (BSA-containing samples) to pass ∼10% of the original volume through the filter. The absence-of-protein condition was used to assess nonspecific binding. Samples were obtained from above (total concentration in the presence of protein) and below (unbound concentration) the filter. Substrate concentrations, determined by either liquid scintillation spectroscopy (taurocholate and digoxin) or LC/MS/MS (pravastatin), were used to calculate fu. The fu of digoxin in the absence or presence of BSA or BSA-FAF in the cell-free system was similar to the fu in sandwich-cultured rat hepatocyte experiments (results not shown); therefore, the fu of pravastatin and taurocholate was not determined in sandwich-cultured rat hepatocytes.
Fluorescence Microscopy. Retention of 5 (and 6)-carboxy-2′,7′-dichlorofluorescein (CDF) in bile canalicular networks in the absence or presence of BSA or BSA-FAF was examined. Hepatocytes were rinsed twice with 2 ml of standard buffer and incubated with 2 ml of the same buffer at 37°C for 10 min. The buffer was removed, and the cells were incubated for 10 min at 37°C with 1.5 ml of CDFDA (2 μM) in standard buffer to preload the bile canaliculi with CDF. Subsequently, hepatocytes were incubated with standard buffer in the absence or presence of 4% (w/v) BSA or BSA-FAF for 10 min, during which time the cells and bile canaliculi were imaged with a Zeiss Axiovert 100TV inverted fluorescent microscope (Carl Zeiss Inc., Thornwood, NY).
Data Analysis. The BEI (%) and unbound intrinsic biliary clearance (intrinsic  , ml/min/kg) were calculated using B-CLEAR technology (Qualyst, Inc., Raleigh, NC) (Liu et al., 1999b):
, ml/min/kg) were calculated using B-CLEAR technology (Qualyst, Inc., Raleigh, NC) (Liu et al., 1999b):  where substrate accumulation in the cell + bile compartments was determined in hepatocytes preincubated in standard buffer, and cellular accumulation of substrate was determined in hepatocytes preincubated with Ca2+-free HBSS.
where substrate accumulation in the cell + bile compartments was determined in hepatocytes preincubated in standard buffer, and cellular accumulation of substrate was determined in hepatocytes preincubated with Ca2+-free HBSS.  where fu represents the unbound fraction of substrate (fu = 1 in the case of no BSA), and AUCmedia represents the area under the substrate concentration-time curve, determined by dividing the sum of the substrate concentration in the incubation medium at the beginning and end of the incubation period by 2 and multiplying by the incubation time (10 min). Intrinsic
where fu represents the unbound fraction of substrate (fu = 1 in the case of no BSA), and AUCmedia represents the area under the substrate concentration-time curve, determined by dividing the sum of the substrate concentration in the incubation medium at the beginning and end of the incubation period by 2 and multiplying by the incubation time (10 min). Intrinsic  values were converted to milliliter per minute per kilogram based on 200 mg of protein/g of liver and 40 g of liver/kg of rat body weight (Seglen, 1976).
values were converted to milliliter per minute per kilogram based on 200 mg of protein/g of liver and 40 g of liver/kg of rat body weight (Seglen, 1976).
Statistical Analysis. Data are represented as mean and the associated S.E.M., S.D., or range, as appropriate. Statistical comparisons were performed with SigmaStat (SPSS Inc., Chicago, IL). The effects of BSA and BSA-FAF, at a single concentration of compound and a single concentration of BSA, on BEI and intrinsic  were evaluated by one-way analysis of variance with Tukey's post hoc test to determine whether the presence and type of BSA affected either of these parameters. The effects on fu were evaluated by two-way analysis of variance with Tukey's post hoc test, using log-transformed data when appropriate for statistical comparisons, to determine whether the fu of each compound was significantly altered by the type of BSA used or by the concentration of compound. In all the cases, p < 0.05 was considered statistically significant.
were evaluated by one-way analysis of variance with Tukey's post hoc test to determine whether the presence and type of BSA affected either of these parameters. The effects on fu were evaluated by two-way analysis of variance with Tukey's post hoc test, using log-transformed data when appropriate for statistical comparisons, to determine whether the fu of each compound was significantly altered by the type of BSA used or by the concentration of compound. In all the cases, p < 0.05 was considered statistically significant.

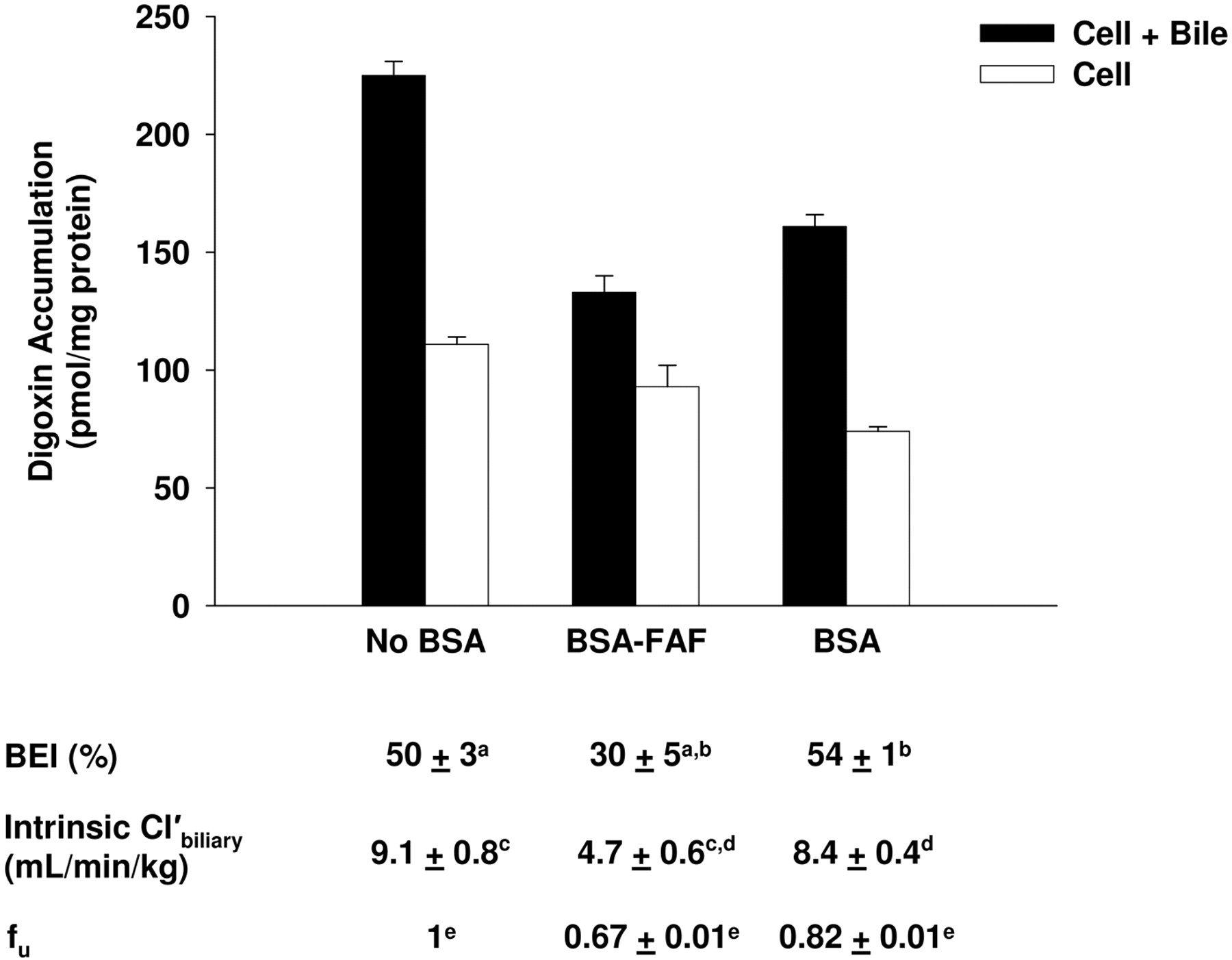
Hepatobiliary disposition of digoxin in the absence or presence of BSA or BSA-FAF. Sandwich-cultured rat hepatocytes were incubated with [3H]digoxin (10 μM) in the absence or presence of 4% (w/v) BSA or BSA-FAF for 10 min following incubation in standard or Ca2+-free buffer. Filled bars represent accumulation in hepatocytes and bile canaliculi (cell + bile). Open bars represent accumulation in hepatocytes (cell). BEI, intrinsic  , and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 3 livers in triplicate. Groups with the same letter indicate a statistically significant difference (p < 0.05).
, and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 3 livers in triplicate. Groups with the same letter indicate a statistically significant difference (p < 0.05).
Results
Effect of BSA and BSA-FAF on Digoxin Hepatobiliary Disposition.Figure 1 shows the effect of BSA and BSA-FAF on the fu, BEI, and intrinsic  of digoxin. The fu, determined by ultrafiltration, was concentration-independent from 0.1 to 30 μM (results not shown) but was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). Digoxin binding to albumin was minimal: 18% in the presence of BSA and 33% in the presence of BSA-FAF. The BEI and intrinsic
of digoxin. The fu, determined by ultrafiltration, was concentration-independent from 0.1 to 30 μM (results not shown) but was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). Digoxin binding to albumin was minimal: 18% in the presence of BSA and 33% in the presence of BSA-FAF. The BEI and intrinsic  of digoxin were not altered by the presence or absence of BSA. However, BSA-FAF significantly decreased both BEI and intrinsic
of digoxin were not altered by the presence or absence of BSA. However, BSA-FAF significantly decreased both BEI and intrinsic  compared with no BSA or BSA conditions (p < 0.05).
compared with no BSA or BSA conditions (p < 0.05).
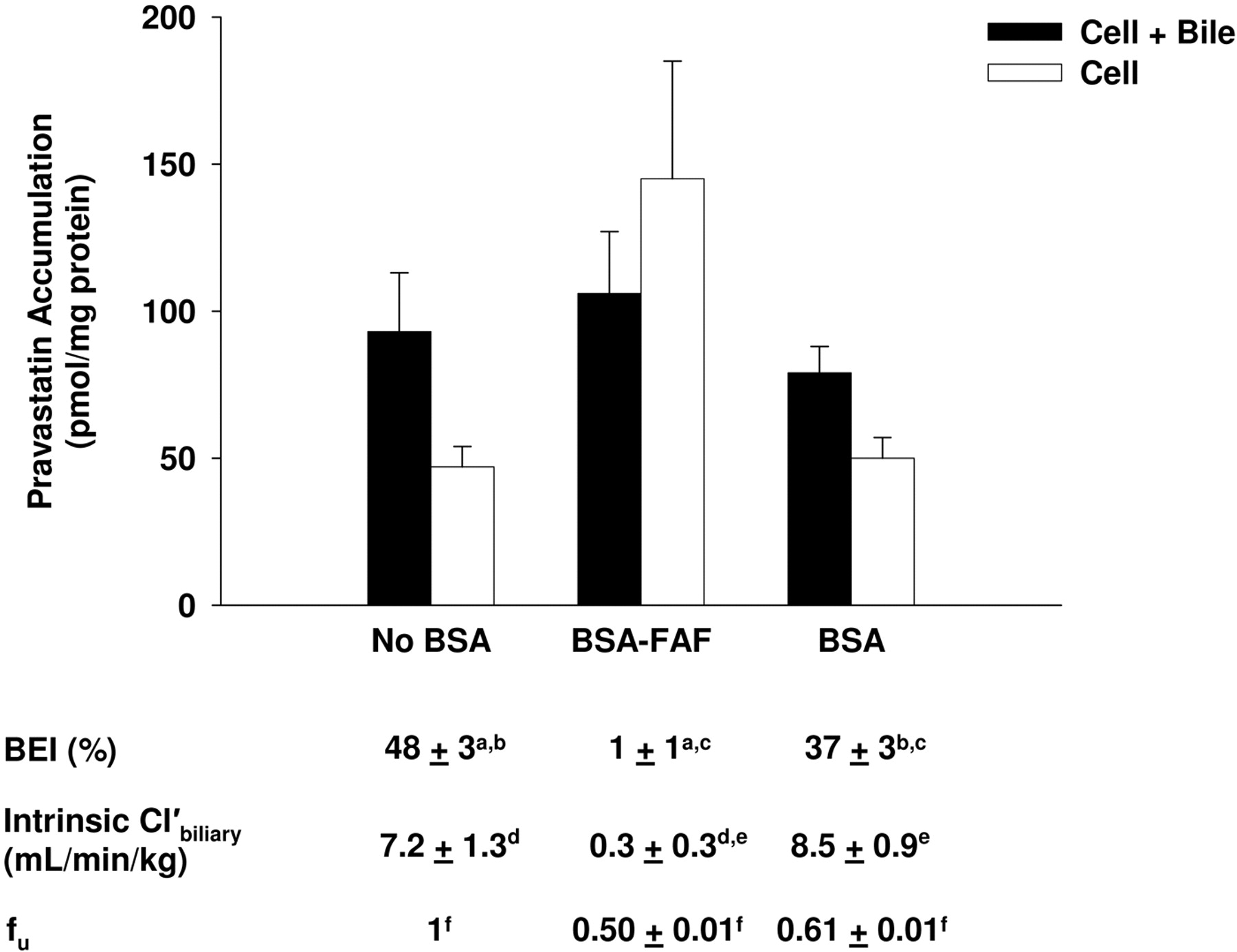
Effect of BSA and BSA-FAF on Pravastatin Hepatobiliary Disposition. The effects of BSA and BSA-FAF on pravastatin fu, BEI, and intrinsic  are illustrated in Fig. 2. The fu was concentration-independent from 1 to 10 μM (results not shown) but was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). Pravastatin was moderately bound to albumin: 39% in the presence of BSA and 50% in the presence of BSA-FAF. The intrinsic
are illustrated in Fig. 2. The fu was concentration-independent from 1 to 10 μM (results not shown) but was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). Pravastatin was moderately bound to albumin: 39% in the presence of BSA and 50% in the presence of BSA-FAF. The intrinsic  of pravastatin was not significantly different in the presence or absence of BSA. However, BSA-FAF significantly decreased both BEI and intrinsic
of pravastatin was not significantly different in the presence or absence of BSA. However, BSA-FAF significantly decreased both BEI and intrinsic  of pravastatin compared with values in the absence or presence of BSA (p < 0.05).
of pravastatin compared with values in the absence or presence of BSA (p < 0.05).
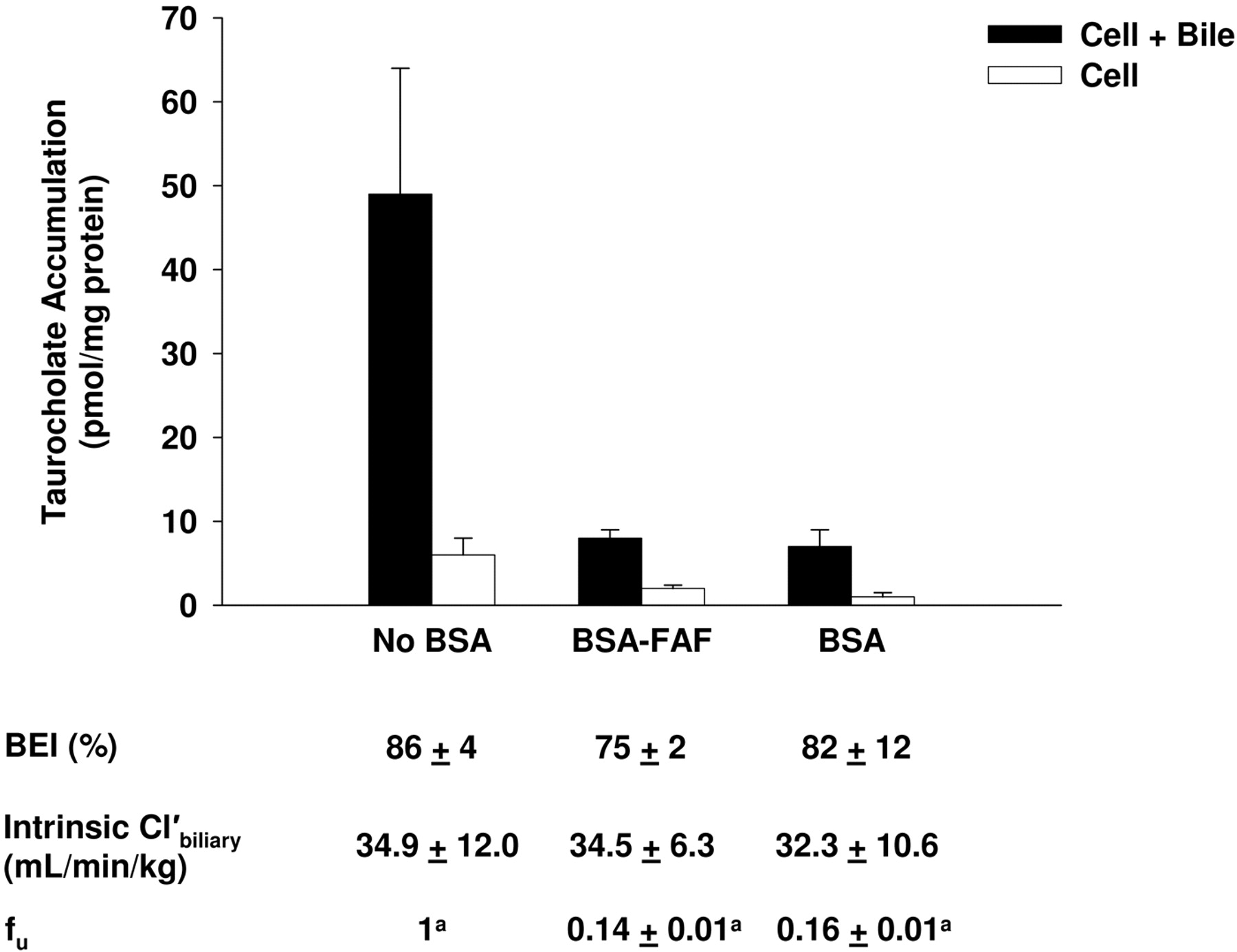
Effect of BSA and BSA-FAF on Taurocholate Hepatobiliary Disposition.Figure 3 shows the effect of BSA and BSA-FAF on the fu, BEI, and intrinsic  of taurocholate. The fu was concentration-independent from 0.1 to 100 μM (results not shown) but was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). Despite the fact that taurocholate was highly bound to albumin (84 and 86% in the presence of BSA and BSA-FAF, respectively), the BEI and intrinsic
of taurocholate. The fu was concentration-independent from 0.1 to 100 μM (results not shown) but was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). Despite the fact that taurocholate was highly bound to albumin (84 and 86% in the presence of BSA and BSA-FAF, respectively), the BEI and intrinsic  of taurocholate were unaffected by BSA.
of taurocholate were unaffected by BSA.
Hepatobiliary disposition of pravastatin in the absence or presence of BSA or BSA-FAF. Sandwich-cultured rat hepatocytes were incubated with pravastatin (5 μM) in the absence or presence of 4% (w/v) BSA or BSA-FAF for 10 min following incubation in standard or Ca2+-free buffer. Filled bars represent accumulation in hepatocytes and bile canaliculi (cell + bile). Open bars represent accumulation in hepatocytes (cell). BEI, intrinsic  , and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 3 livers in triplicate. Groups with the same letter indicate a statistically significant difference (p < 0.05).
, and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 3 livers in triplicate. Groups with the same letter indicate a statistically significant difference (p < 0.05).

Hepatobiliary disposition of taurocholate in the absence or presence of BSA or BSA-FAF. Sandwich-cultured rat hepatocytes were incubated with [3H]taurocholate (1 μM) in the absence or presence of 4% (w/v) BSA or BSA-FAF for 10 min following incubation in standard or Ca2+-free buffer. Filled bars represent accumulation in hepatocytes and bile canaliculi (cell + bile). Open bars represent accumulation in hepatocytes (cell). BEI, intrinsic  , and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 3 livers in triplicate. The taurocholate fu data were log-transformed for statistical comparison. Groups with the same letter indicate a statistically significant difference (p < 0.05).
, and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 3 livers in triplicate. The taurocholate fu data were log-transformed for statistical comparison. Groups with the same letter indicate a statistically significant difference (p < 0.05).
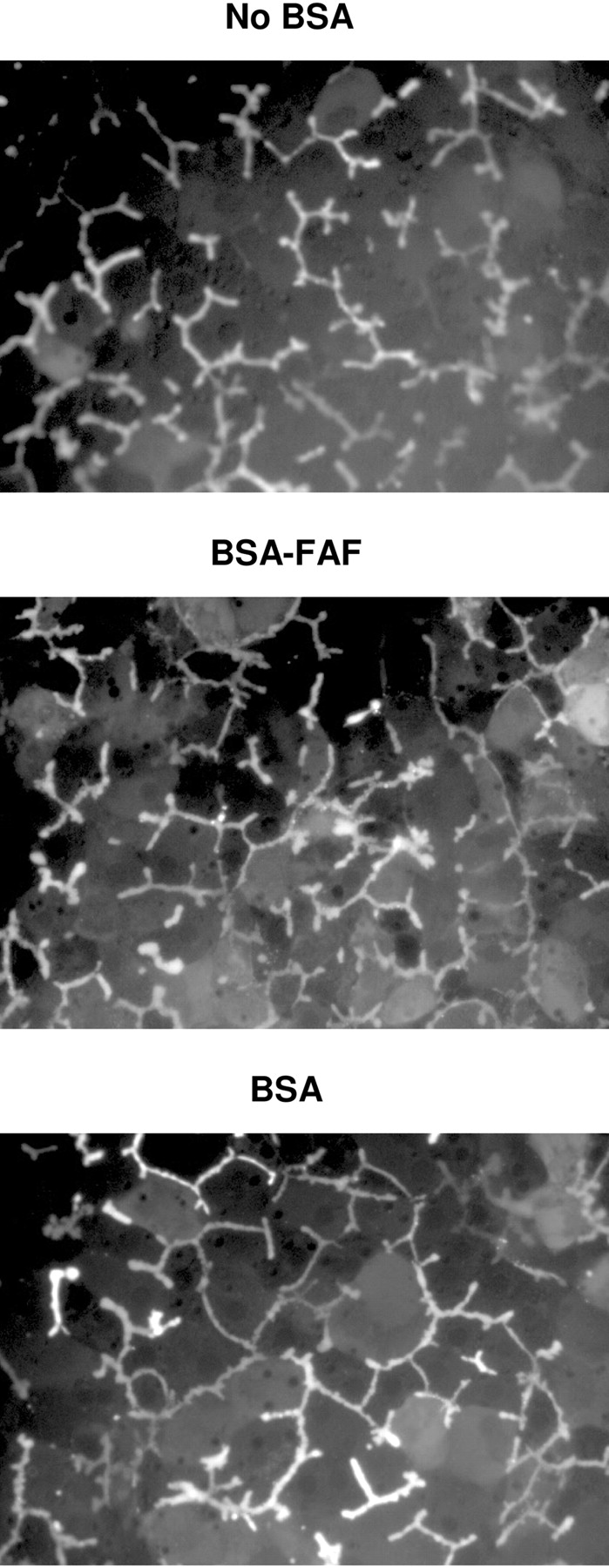
Effect of BSA and BSA-FAF on CDF Retention in Bile Canaliculi. CDFDA readily diffuses into hepatocytes, where it is hydrolyzed rapidly to CDF and excreted into the bile canaliculi by multidrug resistance-associated protein 2. After preincubating hepatocytes for 10 min with CDFDA, CDF retention was evaluated over 10 min in the absence or presence of BSA or BSA-FAF to assess the maintenance of tight junction integrity (Fig. 4). BSA and BSA-FAF had no discernable effects on CDF accumulation in bile canalicular networks over the 10-min period compared with the no BSA condition.

Retention of CDF in bile canaliculi in the absence or presence of BSA or BSA-FAF. Sandwich-cultured rat hepatocytes were incubated with CDFDA (2 μM) in standard buffer for 10 min to preload the bile canaliculi with CDF. Subsequently, hepatocytes were imaged for 10 min in the absence or presence of BSA or BSA-FAF. Representative images of CDF fluorescence during the 10-min period are shown.
Effect of Additional Ca2+on the Hepatobiliary Disposition of Digoxin in the Absence or Presence of BSA or BSA-FAF.Figure 5 shows the effect of BSA and BSA-FAF on the BEI and intrinsic  of digoxin in the presence of additional Ca2+. Similar to the results in Fig. 1, the BEI and intrinsic
of digoxin in the presence of additional Ca2+. Similar to the results in Fig. 1, the BEI and intrinsic  of digoxin were not affected by BSA compared with no BSA, whereas BSA-FAF decreased both parameters. The addition of 1 mM Ca2+ was unable to restore digoxin BEI and intrinsic
of digoxin were not affected by BSA compared with no BSA, whereas BSA-FAF decreased both parameters. The addition of 1 mM Ca2+ was unable to restore digoxin BEI and intrinsic  values in the presence of BSA-FAF to similar values in the presence or absence of BSA. Similar findings were obtained with the addition of 2.5 mM Ca2+ (results not shown).
values in the presence of BSA-FAF to similar values in the presence or absence of BSA. Similar findings were obtained with the addition of 2.5 mM Ca2+ (results not shown).
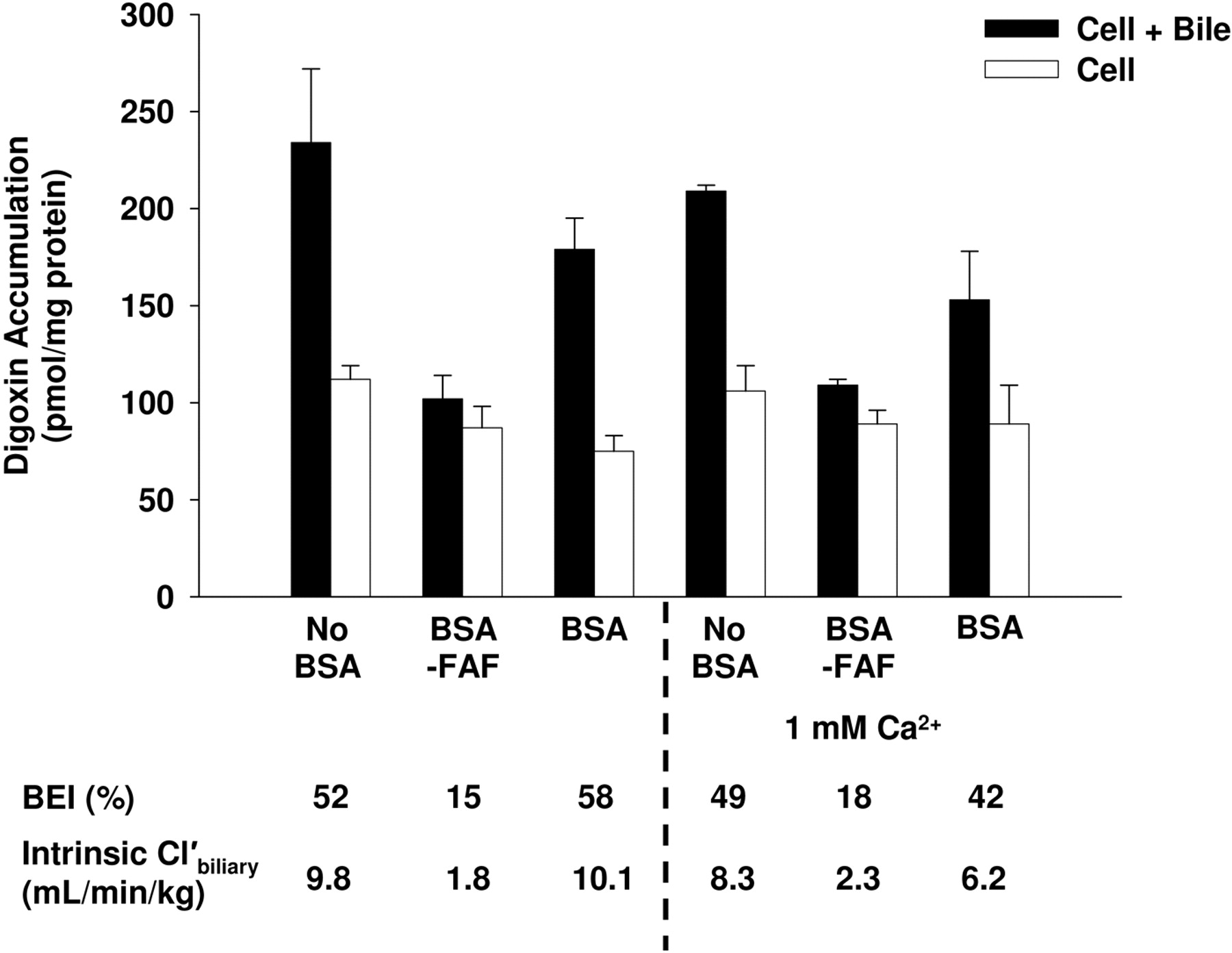
Effect of additional Ca2+ on the hepatobiliary disposition of digoxin in the absence or presence of BSA or BSA-FAF. Sandwich-cultured rat hepatocytes were incubated with [3H]digoxin (10 μM) in the absence or presence of 4% (w/v) BSA or BSA-FAF with or without 1 mM Ca2+ (calcium chloride) for 10 min following incubation in standard or Ca2+-free buffer. Filled bars represent accumulation in hepatocytes and bile canaliculi (cell + bile). Open bars represent accumulation in hepatocytes (cell). The BEI and intrinsic  were determined as described under Materials and Methods. Data are presented as mean ± range; n = 1 liver in duplicate.
were determined as described under Materials and Methods. Data are presented as mean ± range; n = 1 liver in duplicate.
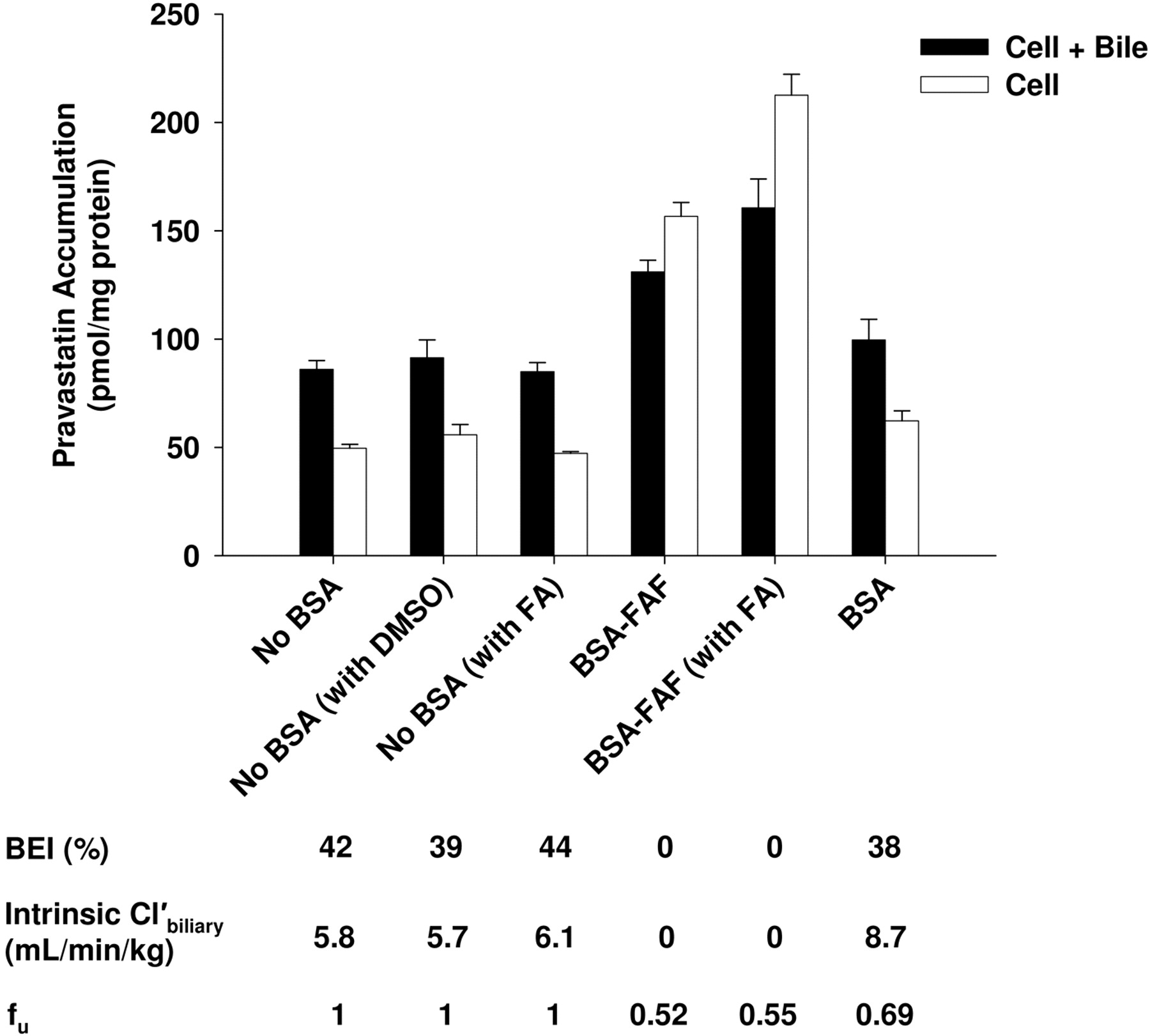
Effect of the Addition of Fatty Acids to BSA or BSA-FAF on Pravastatin Hepatobiliary Disposition. Rowland et al. (2007) reported that palmitic acid (16:0), stearic acid (18:0), oleic acid (18: 1n-9), linoleic acid (18:2n-6), palmitoleic acid (16:1n-7), and linolenic acid (18:3n-3) are six of the predominant long-chain fatty acids present in BSA. To determine whether addition of these fatty acids to BSA-FAF restored the BEI and intrinsic  of pravastatin, additional experiments were conducted (Fig. 6). As described previously (Fig. 2), the fu was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). The BEI and intrinsic
of pravastatin, additional experiments were conducted (Fig. 6). As described previously (Fig. 2), the fu was significantly higher in the presence of BSA compared with BSA-FAF (p < 0.05). The BEI and intrinsic  of pravastatin in the presence of BSA were similar to values in the absence of BSA with or without the addition of fatty acids. BSA-FAF decreased the fu, BEI, and intrinsic
of pravastatin in the presence of BSA were similar to values in the absence of BSA with or without the addition of fatty acids. BSA-FAF decreased the fu, BEI, and intrinsic  of pravastatin compared with the absence or presence of BSA, and the addition of fatty acids to BSA-FAF was unable to mitigate this effect.
of pravastatin compared with the absence or presence of BSA, and the addition of fatty acids to BSA-FAF was unable to mitigate this effect.
Discussion
Protein often is added to in vitro systems to facilitate delivery and improve the stability of drug candidates, to prevent precipitation of water-insoluble compounds, and to minimize adsorption to experimental apparatuses (Artursson, 1990; Wilson et al., 1990; Saha and Kou, 2002; Katneni et al., 2008). Albumin is the most abundant plasma protein and therefore is added most frequently, typically in the form of BSA (Aungst et al., 2000; Yamashita et al., 2000; Krishna et al., 2001; Cross et al., 2003; Neuhoff et al., 2006). Different BSA preparations are commercially available, and BSA-FAF is used frequently for in vitro experiments. The effects of different types of BSA on biliary clearance estimates in sandwich-cultured hepatocytes have not been reported to date. This study evaluated the effect of BSA and BSA-FAF on BEI and biliary clearance in sandwich-cultured rat hepatocytes for three compounds representing different degrees of protein binding: digoxin (low binding), pravastatin (moderate binding), and taurocholate (high binding).

Effect of the addition of fatty acids to BSA-FAF on pravastatin hepatobiliary disposition. Sandwich-cultured rat hepatocytes were incubated with pravastatin (5 μM) in the absence or presence of 4% (w/v) BSA or BSA-FAF with or without addition of long-chain fatty acids (with FA) or DMSO (dimethyl sulfoxide vehicle) for 10 min following incubation in standard or Ca2+-free buffer. Long-chain fatty acids [linoleic acid (5.48 μM), linolenic acid (0.20 μM), oleic acid (6.94 μM), palmitic acid (9.38 μM), palmitoleic acid (0.76 μM), and stearic acid (7 μM)] were added to standard buffer with or without BSA-FAF and preincubated at room temperature for ∼1 h before the addition of pravastatin. Filled bars represent accumulation in hepatocytes and bile canaliculi (cell + bile). Open bars represent accumulation in hepatocytes (cell). BEI, intrinsic  , and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 1 liver in triplicate.
, and fu were determined as described under Materials and Methods. Data are presented as mean ± S.E.M.; n = 1 liver in triplicate.
The equilibrium fu of a compound is an important determinant of hepatic uptake; it is assumed that only the unbound species is available to traverse the hepatic basolateral membrane. Although this is a common assumption, it might not be true in all the cases. For example, if protein binding of a particular compound is labile, such that the rate constants governing binding onset and offset are rapid, uptake may appear to occur for both the bound and unbound pool if the uptake process is very efficient (i.e., if the compound could be classified as a “high extraction” substrate). The fu must be taken into account when calculating the intrinsic  (eq. 2) of digoxin, pravastatin, and taurocholate. As expected, the fu of each compound was significantly decreased in the presence, compared with the absence, of BSA (Figs. 1, 2, 3). Our data consistently show that intrinsic
(eq. 2) of digoxin, pravastatin, and taurocholate. As expected, the fu of each compound was significantly decreased in the presence, compared with the absence, of BSA (Figs. 1, 2, 3). Our data consistently show that intrinsic  , when corrected for fu, is similar in the absence or presence of BSA (Figs. 1, 2, 3). In addition, the BEI of digoxin and taurocholate showed comparable results in the absence and presence of BSA. Similar to the present observations, Liu et al. (2005) reported no difference in digoxin clearance between isolated rat livers perfused with Krebs-Henseleit bicarbonate buffer alone or Krebs-Henseleit bicarbonate buffer containing BSA and red blood cells, after correcting for the fu of digoxin. These results confirm the importance of accounting for the fu of a compound when calculating intrinsic
, when corrected for fu, is similar in the absence or presence of BSA (Figs. 1, 2, 3). In addition, the BEI of digoxin and taurocholate showed comparable results in the absence and presence of BSA. Similar to the present observations, Liu et al. (2005) reported no difference in digoxin clearance between isolated rat livers perfused with Krebs-Henseleit bicarbonate buffer alone or Krebs-Henseleit bicarbonate buffer containing BSA and red blood cells, after correcting for the fu of digoxin. These results confirm the importance of accounting for the fu of a compound when calculating intrinsic  .
.
Surprisingly, the intrinsic  of digoxin and pravastatin in the presence of BSA-FAF was significantly decreased compared with values determined in the presence or absence of BSA when fu was taken into account (Figs. 1 and 2); comparable differences were observed for the BEI. The fu of each compound also was significantly lower in the presence of BSA-FAF (Figs. 1 and 2). Fatty acids may either compete for binding sites on albumin, produce electrostatic effects that influence the binding of compounds to albumin, or bind to albumin and produce conformational changes that affect the binding of other compounds (Bertucci and Domenici, 2002; Kragh-Hansen et al., 2002; Otagiri, 2005). The type of effect that occurs may be substrate-dependent. In this in vitro system, BSA-FAF decreased the fu of digoxin, pravastatin, and taurocholate (Figs. 1, 2, 3). Despite the difference in fu in the presence of BSA-FAF, the increased binding of digoxin and pravastatin to BSA-FAF did not account for the differences observed in BEI and intrinsic
of digoxin and pravastatin in the presence of BSA-FAF was significantly decreased compared with values determined in the presence or absence of BSA when fu was taken into account (Figs. 1 and 2); comparable differences were observed for the BEI. The fu of each compound also was significantly lower in the presence of BSA-FAF (Figs. 1 and 2). Fatty acids may either compete for binding sites on albumin, produce electrostatic effects that influence the binding of compounds to albumin, or bind to albumin and produce conformational changes that affect the binding of other compounds (Bertucci and Domenici, 2002; Kragh-Hansen et al., 2002; Otagiri, 2005). The type of effect that occurs may be substrate-dependent. In this in vitro system, BSA-FAF decreased the fu of digoxin, pravastatin, and taurocholate (Figs. 1, 2, 3). Despite the difference in fu in the presence of BSA-FAF, the increased binding of digoxin and pravastatin to BSA-FAF did not account for the differences observed in BEI and intrinsic  for these compounds.
for these compounds.
Addition of BSA to incubations of HeLa cells stably transfected with rat sodium-taurocholate cotransporting polypeptide (Ntcp) or organic anion-transporting polypeptide (Oatp) 1 (Oatp1a1) decreased the uptake of [35S]sulfobromophthalein (Shi et al., 1995; Hata et al., 2003). Similarly, Cui et al. (2001) reported that addition of human serum albumin to human embryonic kidney 293 cells stably expressing recombinant human OATP2 (OATP1B1) or OATP8 (OATP1B3) abolished OATP8-mediated [3H]sulfobromophthalein uptake but did not significantly affect OATP2-mediated [3H]sulfobromophthalein uptake. These findings suggest that decreased substrate accumulation may depend on whether albumin inhibits the transport protein(s) involved in the uptake of a particular substrate, as well as on the relative contribution of each transport protein to uptake. The hepatic uptake of digoxin and pravastatin is Oatp-mediated (Noé et al., 1997; Hsiang et al., 1999; Tokui et al., 1999; Shitara et al., 2002; Sasaki et al., 2004). Although taurocholate is a substrate for multiple Oatps, Ntcp is the primary transport protein responsible for hepatic uptake of taurocholate in rat hepatocytes (Oude Elferink et al., 1995). In the present study, the accumulation of taurocholate was notably decreased in the presence of BSA or BSA-FAF (Fig. 3). The present findings are consistent with the hypothesis that BSA and BSA-FAF inhibit Ntcp-mediated transport in sandwich-cultured rat hepatocytes. Based on these data, a decrease in the intrinsic  of taurocholate was expected. The observation that BSA-FAF significantly decreased BEI and intrinsic
of taurocholate was expected. The observation that BSA-FAF significantly decreased BEI and intrinsic  values for digoxin and pravastatin, but not taurocholate, further emphasizes the complexity of the albumin effect and suggests that the findings cannot be explained simply by direct inhibition of specific transport proteins.
values for digoxin and pravastatin, but not taurocholate, further emphasizes the complexity of the albumin effect and suggests that the findings cannot be explained simply by direct inhibition of specific transport proteins.
One possible mechanism by which BSA-FAF might affect BEI and intrinsic  in sandwich-cultured rat hepatocytes involves alterations in the maintenance of tight junction integrity. To investigate this possibility, CDF retention in sandwich-cultured rat hepatocytes in the absence or presence of BSA and BSA-FAF was studied over a 10-min period (Fig. 4). CDFDA is hydrolyzed to CDF within hepatocytes and is transported into the bile canalicular networks by multidrug resistance-associated protein 2. CDF accumulates in these networks if the integrity of the tight junction complexes has not been compromised by the absence of Ca2+ (Liu et al., 1999b) or some other factor. No obvious differences were observed in CDF retention in bile canalicular networks in the presence of BSA or BSA-FAF compared with accumulation in networks in the absence of BSA. These results suggest that the bile canalicular networks remain sealed in the presence of BSA and BSA-FAF, and that the proteins comprising the tight junctions are not modulated.
in sandwich-cultured rat hepatocytes involves alterations in the maintenance of tight junction integrity. To investigate this possibility, CDF retention in sandwich-cultured rat hepatocytes in the absence or presence of BSA and BSA-FAF was studied over a 10-min period (Fig. 4). CDFDA is hydrolyzed to CDF within hepatocytes and is transported into the bile canalicular networks by multidrug resistance-associated protein 2. CDF accumulates in these networks if the integrity of the tight junction complexes has not been compromised by the absence of Ca2+ (Liu et al., 1999b) or some other factor. No obvious differences were observed in CDF retention in bile canalicular networks in the presence of BSA or BSA-FAF compared with accumulation in networks in the absence of BSA. These results suggest that the bile canalicular networks remain sealed in the presence of BSA and BSA-FAF, and that the proteins comprising the tight junctions are not modulated.
Preparation of BSA-FAF involves the incubation of BSA with activated charcoal. In addition to removal of fatty acids, other components may be removed, including calcium and other ions. Thus, BSA-FAF may have increased potential to bind calcium. Therefore, we evaluated the addition of physiological concentrations of calcium. Additional 1 mM Ca2+ (Fig. 5) or 2.5 mM Ca2+ (data not shown) did not mitigate the effect of BSA-FAF on the BEI or intrinsic  of digoxin, suggesting that the mechanism of the BSA-FAF effect is not through the modulation of Ca2+ levels. It is unlikely that a contaminant in the BSA-FAF preparation is chelating Ca2+ and preventing the maintenance of tight junction integrity and formation of sealed bile canalicular networks.
of digoxin, suggesting that the mechanism of the BSA-FAF effect is not through the modulation of Ca2+ levels. It is unlikely that a contaminant in the BSA-FAF preparation is chelating Ca2+ and preventing the maintenance of tight junction integrity and formation of sealed bile canalicular networks.
Fatty acids have been reported to inhibit the activity of some metabolic enzymes (Rowland et al., 2007, 2008a,b), and any BSA preparation can protect against this by sequestering the inhibitory fatty acids (Rowland et al., 2007). Therefore, we evaluated whether the addition of six of the predominant long-chain fatty acids present in BSA [linoleic acid, linolenic acid, oleic acid, palmitic acid, palmitoleic acid, and stearic acid (Rowland et al., 2007)] would mitigate the decrease in the BEI and intrinsic  of pravastatin observed in the presence of BSA-FAF compared with values in the presence or absence of BSA. Addition of fatty acids did not modulate the effect of BSA-FAF on the BEI and intrinsic
of pravastatin observed in the presence of BSA-FAF compared with values in the presence or absence of BSA. Addition of fatty acids did not modulate the effect of BSA-FAF on the BEI and intrinsic  of pravastatin (Fig. 6). Further work is required to elucidate the mechanism(s) responsible for the decreased BEI and intrinsic
of pravastatin (Fig. 6). Further work is required to elucidate the mechanism(s) responsible for the decreased BEI and intrinsic  values observed in the presence of BSA-FAF.
values observed in the presence of BSA-FAF.
In conclusion, the results of this study indicate that the type of BSA that is added to sandwich-cultured rat hepatocytes can affect the BEI and predicted in vivo biliary clearance values. Addition of BSA yielded results consistent with the BEI and intrinsic  determined in the absence of BSA. In contrast, BSA-FAF significantly decreased the BEI and intrinsic
determined in the absence of BSA. In contrast, BSA-FAF significantly decreased the BEI and intrinsic  of some substrates. The selection of proteins for addition to in vitro systems requires careful consideration. The present observations suggest that, when necessary, BSA rather than BSA-FAF should be added in vitro to sandwich-cultured rat hepatocytes for determination of BEI and biliary clearance values.
of some substrates. The selection of proteins for addition to in vitro systems requires careful consideration. The present observations suggest that, when necessary, BSA rather than BSA-FAF should be added in vitro to sandwich-cultured rat hepatocytes for determination of BEI and biliary clearance values.
Acknowledgments
We thank Yi-Wei Rong for technical expertise in the isolation of rat hepatocytes, Dr. Arlene Bridges for LC/MS/MS analysis of pravastatin samples, and Dr. Koji Abe and Dr. Daniel Bow for insightful comments regarding this manuscript.
Footnotes
-
This work was supported by National Institutes of Health Grant GM41935. K.K.W. was supported by a postdoctoral fellowship from GlaxoSmithKline Inc. Data were presented at the American Association of Pharmaceutical Scientists Annual Meeting and Exposition in San Diego, CA, November 2007.
-
K.L.R.B. is cofounder and Chair of the Scientific Advisory Board for Qualyst, Inc., which has exclusively licensed the sandwich-cultured hepatocyte technology for quantification of biliary excretion (B-CLEAR).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.108.020842.
-
ABBREVIATIONS: BSA, bovine serum albumin; UGT, UDP-glucuronosyltransferase; BSA-FAF, essentially fatty acid-free bovine serum albumin; BEI, biliary excretion index; fu, unbound fraction; DMEM, Dulbecco's modified Eagle's medium; MEM, minimal essential medium; CDFDA, 5 (and 6)-carboxy-2′,7′-dichlorofluorescein diacetate; HBSS, Hanks' balanced salt solution; BCA, bicinchoninic acid; LC/MS/MS, liquid chromatography with detection by tandem mass spectrometry; CDF, 5 (and 6)-carboxy-2′,7′-dichlorofluorescein; intrinsic
 , unbound intrinsic biliary clearance; Ntcp, sodium-taurocholate cotransporting polypeptide; Oatp/OATP, organic anion-transporting polypeptide.
, unbound intrinsic biliary clearance; Ntcp, sodium-taurocholate cotransporting polypeptide; Oatp/OATP, organic anion-transporting polypeptide. - Received February 6, 2008.
- Accepted July 22, 2008.

- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}