


Abstract
Silymarin, an extract from seeds of Silybum marianum, is used by 8 to 33% of patients to self-treat chronic viral hepatitis C in the United States. Studies in humans and rodents suggest that biliary excretion of glucuronide and sulfate conjugates is the major route for silymarin's elimination. To determine the role of multidrug resistance-associated protein 2 (Mrp2) (Abcc2) in the biliary excretion of silymarin, the hepatobiliary disposition of the six major silymarin flavonolignans was studied using isolated perfused livers (IPRLs) from wild-type (WT) and Mrp2-deficient (TR-) Wistar rats. For all the flavonolignans, approximately 96% of the dose was cleared from perfusate within 30 min in both WT and TR- livers, and <5% of parent was recovered in bile or perfusate by the end of the perfusion. In WT livers, the percentage of dose excreted as conjugates into bile varied for each flavonolignan (silychristin, 51.6 ± 9.3%; silydianin, 101.5 ± 28.3%; silybin A, 21.0 ± 8.3%; silybin B, 31.7 ± 13.2%; isosilybin A, 50.5 ± 23.6%; isosilybin B, 42.8 ± 19.3%). Among the flavonolignans, only silydianin was primarily glucuronidated and almost completely recovered in bile. In TR- livers, biliary excretion of flavonolignan conjugates was reduced 80 to 92%, with 30 to 83% of each flavonolignan conjugate recovered in perfusate compared with only 5 to 30% at 90 min. Biliary excretion of glucuronide and sulfate conjugates of all the flavonolignans were reduced by 94 to 98% and 73 to 84%, respectively, in TR- IPRLs. These data indicate a primary role for Mrp2 in the biliary elimination of silymarin flavonolignan conjugates.
Silymarin, an herbal extract commonly known as milk thistle, is used by approximately 8 to 33% of patients in the United States to self-treat chronic hepatitis C virus (HCV) (Seeff et al., 2008). Silymarin is a complex mixture of six major flavonolignans [silybin A (SA) and silybin B (SB), isosilybin A (ISA) and isosilybin B (ISB), silychristin (SC), silydianin (SD)], the most common class of flavonoids in milk thistle extract, as well as other polyphenolic compounds (Kim et al., 2003). SA and SB are the most abundant flavonolignans in milk thistle extracts and are used to standardize commercial preparations. Silymarin is hepatoprotective in laboratory animals to the toxic effects of carbon tetrachloride, ethanol, and acetaminophen (Crocenzi and Roma, 2006). Recently, two independent laboratories have shown the antiviral effects of silymarin in hepatitis C virus replicon assays (Bonifaz et al., 2007; Polyak et al., 2007). However, the beneficial effects of silymarin in liver disease have not been established because of inconsistencies in results obtained from several randomized clinical trials that may reflect various trial designs and the use of different formulations (Mayer et al., 2005). In addition, recent data suggest that the pharmacokinetics of silymarin is influenced by the type and stage of liver disease (Schrieber et al., 2008b). Although the precise pathways of metabolism and excretion have not been identified, limited data from human and animal studies suggest that flavonolignans primarily undergo rapid and extensive phase II metabolism and biliary excretion (Morazzoni et al., 1992, 1993; Schandalik et al., 1992).
In rats and humans, silymarin is characterized by low bioavailability, rapid conjugation, and biliary excretion of silymarin and flavonolignan conjugates. In rats, only 2% of the p.o. administered dose of silymarin was recovered in the bile unchanged, whereas the remaining was recovered as sulfate and glucuronide conjugates (Morazzoni et al., 1993). However, the transport proteins primarily responsible for biliary excretion of silymarin and its metabolites have not been identified.
Silymarin is a potent inhibitor of breast cancer resistance protein in MCF-7 cells (Zhang et al., 2004). Additionally, it has been shown that silymarin can inhibit P-glycoprotein (P-gp)-mediated efflux of digoxin in human intestinal Caco-2 cells (Zhang and Morris, 2003). Multidrug resistance-associated protein 2 (Mrp2), which is expressed in the liver, gut, kidney, and brain, mediates the biliary elimination of many flavonol conjugates (Hoffmann and Kroemer, 2004; Cermak and Wolffram, 2006). Mrp2 (Abcc2) is a member of the ATP-binding cassette superfamily of membrane transporters, which is responsible for the biliary excretion of organic anions, glucuronide, glutathione, and sulfate conjugates of endogenous compounds, as well as xenobiotics (König et al., 1999; Zamek-Gliszczynski et al., 2005, 2006b). The specific role of each of these canalicular transport proteins in the excretion of silymarin and its conjugates has not been established.
The isolated perfused rat liver (IPRL) is a valuable and commonly used ex vivo model for studying the metabolism and transport mechanisms involved in the hepatobiliary disposition of drugs. This model permits exposure of the liver to different test substances, allows repeated sampling to determine disposition of parent compound and metabolites in perfusate and bile, and is independent of the influence of other organ systems (Gores et al., 1986). Compared with isolated hepatocytes, the hepatic architecture, cell polarity, and bile flow are preserved in the isolated perfused liver (Jansen et al., 1985), making this a useful model to study metabolism and biliary excretion of various substances. The IPRL technique has been used successfully in various genetic animal models to investigate the precise mechanisms of hepatobiliary disposition of drugs. For example, a number of studies have used IPRL to examine the role of Mrp2 using TR- rats, which are mutant rats that lack functional Mrp2 at the canalicular membrane (Jansen et al., 1985; McDonagh et al., 2001), similar to patients with Dubin-Johnson syndrome (Paulusma et al., 1997). Such studies have shown marked reduction in biliary excretion of glucuronide and sulfate conjugates of drugs and other compounds in TR- rats, implying that Mrp2 is responsible for the biliary excretion of organic anions.
The purpose of this study was to determine the role of Mrp2 in the biliary excretion of the six major silymarin flavonolignans and their conjugates by using IPRLs from Mrp2-competent [wild-type (WT)] and Mrp2-deficient (TR-) Wistar rats. Pharmacokinetic parameters were determined for both the perfusate and bile compartments.
Materials and Methods
Chemicals. Silymarin, dimethyl sulfoxide, β-glucuronidase (EC 3.3.1.3.1; type B-10 from bovine liver), sulfatase (EC3.1.6.1; type H-1 from Helix pomatia), d-saccharic acid 1,4-lactone, naringenin, and taurocholate were purchased from Sigma-Aldrich (St. Louis, MO). Reference standards for SC were obtained from ChromaDex (Santa Ana, CA), and SD was purchased from U.S. Pharmacopoeia (Rockville, MD). The composition of silybin (Silibinin; Sigma-Aldrich) was confirmed to be a mixture of SA and SB by liquid chromatography/electrospray ionization/mass spectrometry (LC/ESI/MS), and the specific contents of SA and SB were analyzed to be 48 and 52%, respectively. ISA and ISB reference standards were obtained as a gift from Ulrich Mengs (Madaus GmbH, Cologne, Germany). The purity of all six silymarin flavonolignan standards was between 97 and 99%. All the other chemicals and reagents were of analytical grade and available from commercial sources.
Animals. Male Wistar rats (200-250 g) from Charles River Laboratories, Inc. (Raleigh, NC) and male TR- rats bred in our facility (275-300 g; breeding stock obtained from Dr. Mary Vore, University of Kentucky, Lexington, KY) were used as liver donors for perfused liver studies. Retired male Wistar breeder rats (400-500 g; Charles River Laboratories, Inc.) were used as blood donors. Rats were housed in an alternating 12-h light/dark cycle with rat chow and water provided ad libitum. All the animals were allowed to acclimate for at least 1 week before experimentation. Rats were anesthetized with ketamine/xylazine (60:12 mg/kg i.p). The Institutional Animal Care and Use Committee at the University of North Carolina approved all the animal procedures.
Liver Perfusion Studies. Recirculating IPRL studies were conducted with WT and TR- rat livers as described previously (Brouwer and Thurman, 1996). Briefly, after portal vein and bile duct cannulation, livers were perfused in situ with oxygenated buffer, pH 7.4. Livers were removed from the body cavity and placed in a humidified perfusion chamber heated to maintain liver temperature at 37°C. Perfusion was continued with oxygenated Krebs-Henseleit bicarbonate buffer containing 20% (v/v) heparinized male rat blood at a flow rate of 20 ml/min (80 ml, total volume). Livers and perfusate were allowed to acclimate 10 min before addition of 26 μg/ml silymarin. Liver viability was assessed by monitoring portal pressure (<15 cm H2O), observing gross morphology, and measuring initial bile flow in WT and TR- IPRLs (>0.8 and >0.3 μl/min/g liver, respectively). Taurocholate (0.5 μmol/min, in saline) was infused into the perfusate reservoir to maintain bile flow. Bile was collected continuously at 10-min intervals, and the volume was determined gravimetrically (specific gravity 1.0). Perfusate (0.5 ml) was sampled every 10 min and immediately centrifuged to obtain supernatant for analysis. After perfusion, livers were blotted dry and weighed. All the samples collected from this experiment were stored at -80°C until further analysis.
Analysis of Silymarin Flavonolignans in Perfusate and Bile. Identification and quantification of silymarin flavonolignans in the perfusate and bile was carried out by LC/ESI/MS using a previously established method with slight modifications (Wen et al., 2008). Briefly, separation of silymarin flavonolignans was performed using an Agilent Technologies (Palo Alto, CA) HP 1100 LC system with a C18 SecurityGuard cartridge (4 × 2.0 mm i.d.; Phenomenex, Torrance, CA) and a Luna C18 (2) analytical column (50 × 2.0 mm i.d., 3 μm; Phenomenex). High-performance liquid chromatography conditions were as follows: mobile phase, methanol: 1% glacial acetic acid, pH 2.8 (43:57, v/v) with isocratic elution; flow rate, 0.3 ml/min; injection volume, 25 μl; run time, 13 min. Typical retention times of silymarin flavonolignans SC, SD, SA, SB, ISA, ISB, and naringenin (internal standard) under the experimental conditions were 1.8, 2.1, 4.4, 5.5, 7.4, 8.2, and 5.0 min, respectively. MS analysis and detection were conducted by an HP 1100 LC-MSD system (Agilent Technologies) with an electrospray interface in the negative ESI ionization mode. MS parameters used for quantitative analysis were capillary voltage, -3500 V; fragmentor voltage, 150 V; drying gas temperature, 350°C; nebulizer gas pressure, 35 psig; drying gas flow, 8 l/ml; scan mode, selective ion monitoring with [M-H]- for silymarin flavonolignans (m/z 481), silymarin sulfates (m/z 561), silymarin glucuronides (m/z 657), and naringenin (m/z 271), respectively. Split calibration curves for the free (unconjugated) and total (unconjugated plus conjugated) flavonolignans in the concentration range of 10 to 500 ng/ml and 500 to 10,000 ng/ml were generated using reference standards of SC, SD, SA, SB, ISA, and ISB. Mixed standard solutions containing SC, SD, SA, SB, ISA, and ISB were spiked into blank perfusates, and bile was treated as described in the perfusate and bile sample preparation (see below). Concentrations of silymarin flavonolignans in the samples were estimated with 1/x2 weighted least-squares regression equations derived from the peak area ratios of individual silymarin flavonolignans to that of the internal standard. The limit of detection was 5 ng/ml, and limit of quantification for the silymarin flavonolignans was 10 ng/ml. Intraday and interday precisions were 1.7 to 11% and 4.5 to 14%, respectively.
Perfusate and bile samples were initially diluted 1:5 and 1:50, respectively. Aliquots of perfusate and bile samples, as well as blank perfusate and bile spiked with standards of SC, SD, SA, SB, ISA, and ISB, were protein-precipitated by the addition of 300 μl of ice-cold acetonitrile and 1% glacial acetic acid containing internal standard, naringenin (200 ng). After the removal of protein by centrifugation at 15,000g for 15 min at 4°C, the supernatants were transferred and evaporated with a gentle stream of nitrogen at 45°C in a water bath. The residue was reconstituted in 100 μl of the high-performance liquid chromatography mobile phase and centrifuged at 10,000g for 10 min at 4°C; 25 μl of the reconstituted supernatants was introduced for LC/ESI/MS analysis.
Enzyme Hydrolysis of Sulfate and Glucuronide Conjugates of Silymarin. Concentrations of sulfated, glucuronidated, and total (unconjugated and conjugated) silymarin flavonolignans in perfusate and bile were measured after hydrolysis with sulfatase, β-glucuronidase, and a mixture of sulfatase and β-glucuronidase, respectively (Wen et al., 2008). Unconjugated silymarin flavonolignan concentrations in perfusate and bile were directly determined without enzyme hydrolysis. Conjugated silymarin flavonolignan concentrations were determined as the difference between total (determined with enzyme hydrolysis) and unconjugated (without enzyme hydrolysis) and expressed as parent equivalents of silymarin flavonolignans. Briefly, aliquots of 100-μl diluted perfusate and bile were treated with sulfatase (80 U/ml in the final incubation) containing d-saccharic acid 1,4-lactone (10 mM), β-glucuronidase containing 0.1 M phosphate buffer (8000 U/ml in the final incubation), and a mixture of sulfatase (80 U/ml) and β-glucuronidase (8000 U/ml), respectively. The individual mixtures with different hydrolytic enzymes were buffered by 0.25 M sodium acetate, pH 5.0, and incubated (final volume 120 μl) at 37°C with gentle shaking for 6 h. The reactions were terminated by the addition of 300 μl of ice-cold acetonitrile containing 1% glacial acetic acid and internal standard naringenin (200 ng). After the removal of protein by centrifugation at 15,000g for 15 min at 4°C, the supernatants were evaporated and treated as described in the analysis of silymarin flavonolignans in perfusate and bile; the final reconstituted samples were analyzed by LC/ESI/MS.

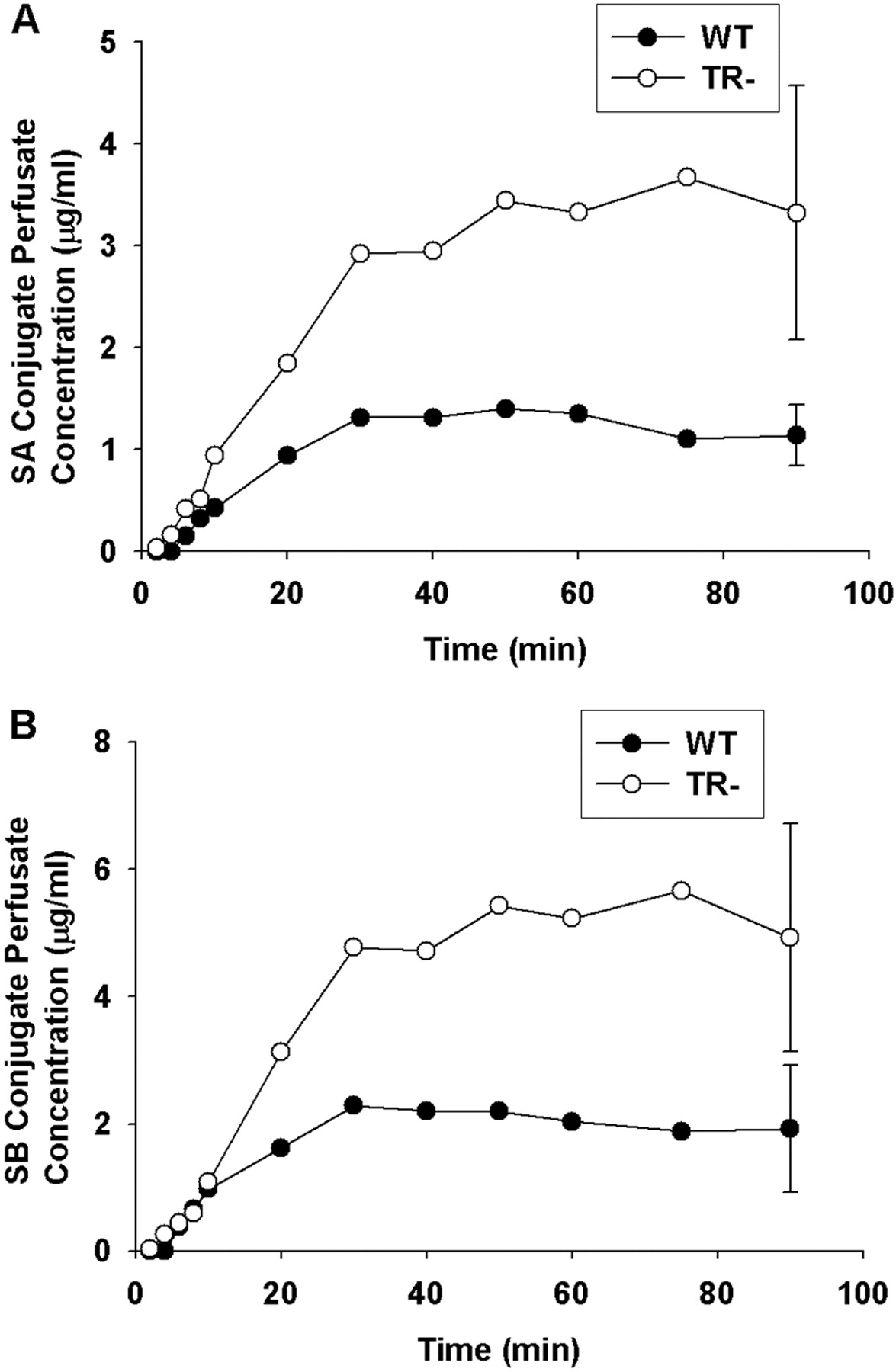
Perfusate concentration versus time profiles of unconjugated SA (A) and SB (B) in isolated perfused livers from wild-type (•) and TR- (○) rats. Silymarin (26 μg/ml) was added to the recirculating perfusate reservoir, and livers from male Wistar rats were perfused for 90 min. Concentrations of SA and SB were determined by LC/MS analysis as described under Materials and Methods. Data are expressed as mean of n = 3 livers in each group; mean ± S.E.M. at the 90-min time point.
Pharmacokinetics and Statistical Analyses. The six major flavonolignans comprised 2.08 mg or 53.88% of the 3.86-mg milk thistle extract added to the 80 ml of perfusate. Initial quantities of the six flavonolignans added to the perfusate were as follows: 470 μg of SC, 188 μg of SD, 443 μg of SA, 726 μg of SB, 172 μg of ISA, and 81 μg of ISB (Wen et al., 2008). Thus, the initial total perfusate concentration of flavonolignans was 26 μg/ml. All the data are expressed as mean ± S.E.M. from three individual experiments. Pharmacokinetic parameters for individual silymarin flavonolignans (unconjugated and conjugates, respectively) were calculated using WinNonlin 4.1 (Pharsight, Mountain View, CA). The individual unconjugated silymarin flavonolignan concentrations over 90 min in the perfusate and extrapolated to infinity were used to calculate the clearance (Cl) from the perfusate, and the estimated initial perfusate concentration (C0), using a noncompartmental i.v. model. The area under the curve during the 90-min perfusion (AUC0→90 min) was calculated according to the log-linear trapezoidal rule. Biliary clearance (Clb) for each silymarin flavonolignan during the 90-min collection interval was calculated as the cumulative amount of unconjugated flavonolignan excreted in the bile (Ae) divided by the perfusate AUC0→90 min of the unconjugated flavonolignans. Apparent hepatobiliary clearance of conjugated silymarin flavonolignans was defined as the cumulative amounts of conjugated flavonolignans excreted into the bile (Ae) divided by the plasma AUC0→90 min for each of the flavonolignans. The following assumptions are included in the definition of hepatobiliary clearance: 1) conjugates excreted into the perfusate return to the liver for biliary excretion; and 2) there is no competition between flavonolignans and flavonolignan conjugates for biliary excretion. Thus, the apparent hepatobiliary clearance encompasses the formation clearance to conjugates that are ultimately excreted in bile, as well as biliary excretion of conjugates. Statistical differences between WT and TR- IPRL data were assessed using analysis of variance followed by Student's t test at a significance level of p < 0.05.
Results
Pharmacokinetics of Silymarin Flavonolignans in IPRL. Pharmacokinetic parameters describing the disposition of the six major silymarin flavonolignans in the WT and TR- IPRLs during the 90-min perfusion are presented in Table 1. In addition, the complete concentration-time profiles from which pharmacokinetic data were obtained for SA and SB are depicted in Fig. 1 because they comprise approximately 60% of the total flavonolignans in silymarin extracts and are the most well studied flavonolignans in vivo. As shown in Fig. 1, similar perfusate concentration-time profiles were obtained from WT and TR- IPRLs; approximately 97% of SA and SB was cleared from the perfusate by 30 min. Similar concentration-time profiles also were observed for SC, SD, ISA, and ISB in both WT and TR- IPRLs (data not shown). When the extrapolated initial perfusate concentrations (C0) for each of the six flavonolignans were summed, the total concentration of silymarin flavonolignans in the perfusate amounted to 21.9 μg/ml for WT and 21.3 μg/ml for TR- IPRLs, which were comparable with the theoretical perfusate silymarin concentration of 26 μg/ml at the start of each perfusion. Consistent with the concentration-time profiles, no significant differences in the clearance of each silymarin flavonolignan from the perfusate were observed between WT and TR- IPRLs. These data suggest that the exposure of WT and TR- IPRLs to each flavonolignan was similar, allowing for mass-balance comparisons.
Pharmacokinetic parameters of unconjugated silymarin flavonolignans in perfusate of WT and TR− rats
Pharmacokinetic parameters of unconjugated silymarin flavonolignans after addition of silymarin (26 μg/ml) to the perfusate. Data are expressed as mean ± S.E.M. (n = 3).

Perfusate concentration versus time profiles of SA (A) and SB (B) conjugates (glucuronide + sulfate) in isolated perfused livers from wild-type (•) and TR- (○) rats. Silymarin (26 μg/ml) was added to the recirculating perfusate reservoir, and perfusate samples were treated with or without a mixture of β-glucuronidase and sulfatase to obtain the total and unconjugated flavonolignan concentrations, respectively. Perfusate conjugate concentrations of SA and SB were determined as the difference between the total and unconjugated concentrations for each flavonolignan. Data are expressed in equivalents of the unconjugated flavonolignans as mean of n = 3 livers in each group; mean ± S.E.M. at the 90-min time point.
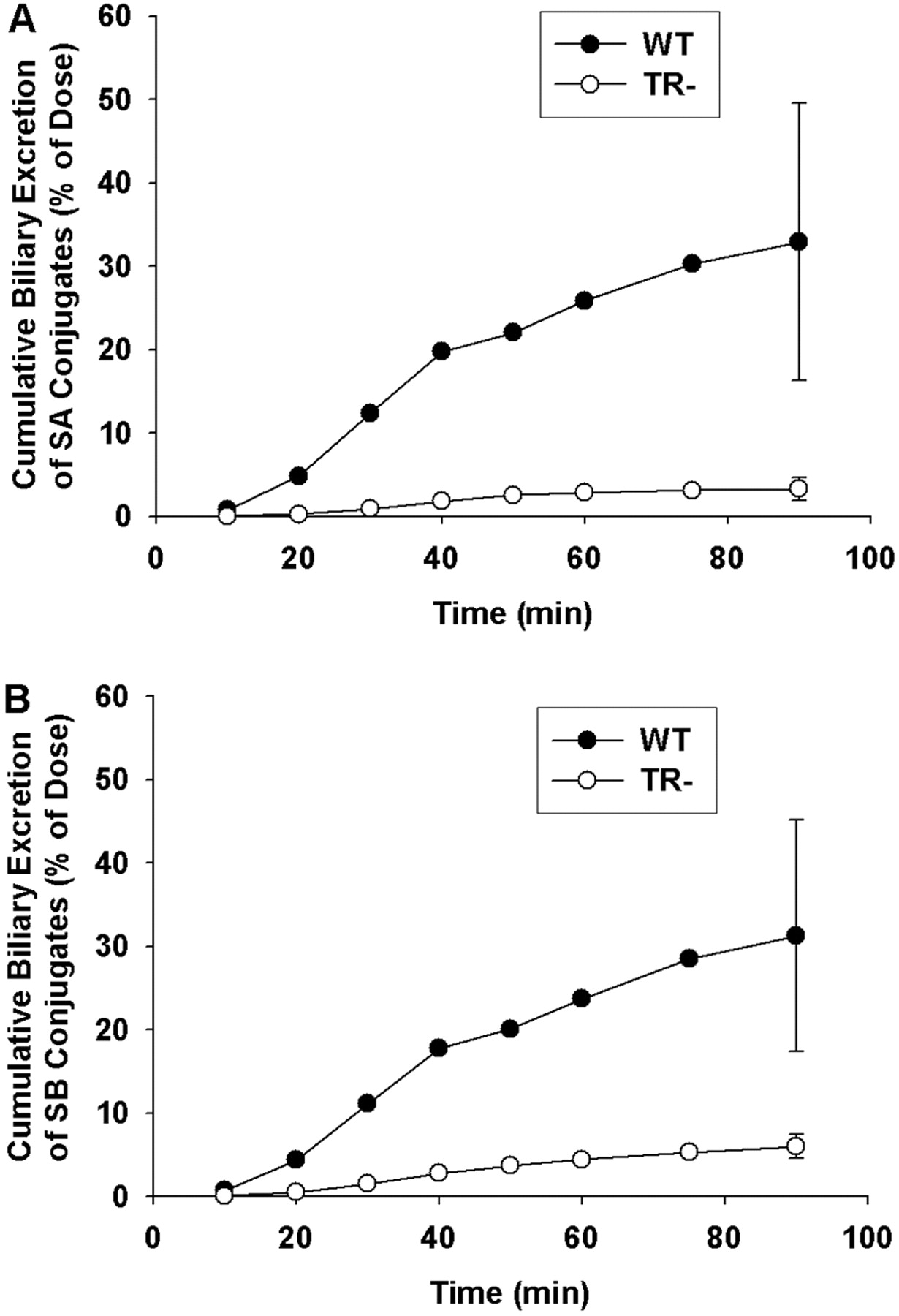
Cumulative biliary excretion of SA (A) and SB (B) conjugates (glucuronide + sulfate) in isolated perfused livers from wild-type (•) and TR- (○) rats. Data are expressed in equivalents of the unconjugated flavonolignans as mean of n = 3 livers in each group; mean ± S.E.M. at the 90-min time point.
Concentration-Time Profiles of Flavonolignan Conjugates in Perfusate. Concentration-time profiles for SA and SB conjugates in the perfusate are shown in Fig. 2, A and B, respectively. After an initial lag time of approximately 6 min, concentrations of flavonolignan conjugates increased rapidly in perfusate and reached a plateau after ∼50 min. The concentrations of SA and SB as conjugates in the perfusate of WT IPRLs at 90 min were 1.2 and 1.9 μg/ml, respectively, or approximately 21 to 22% of the initial dose of the parent flavonolignans. Similar concentration-time profiles were observed for the other flavonolignans (data not shown). Perfusate conjugate concentrations of ISA (0.6 μg/ml), ISB (0.1 μg/ml), SC (0.3 μg/ml), and SD (0.1 μg/ml) at 90 min accounted for approximately 28, 10, 5, and 4% of the dose for each flavonolignan, respectively.
In contrast to WT IPRLs, approximately 78 to 80% of the dose for SA and SB was excreted into perfusate as conjugates at 90 min in TR- IPRLs. Similar conjugate concentration-time profiles were observed for the other flavonolignans (data not shown).
Biliary Excretion of Silymarin Flavonolignans and Flavonolignan Conjugates. Representative cumulative biliary excretion profiles of SA and SB conjugates by WT and TR- IPRLs are shown in Fig. 3, A and B, respectively. Similar biliary excretion profiles also were obtained for SC, SD, ISA, and ISB (data not shown). For WT IPRLs, the cumulative biliary excretion profiles for SA and SB revealed a time lag between 10 and 20 min, after which the amount of flavonolignan conjugates excreted into bile increased rapidly until 40 min. This lag period coincided with the 20-min period where most of the silybins are extracted from the perfusate and into the liver (see Fig. 1), and flavonolignan conjugate concentrations plateau in the perfusate (see Fig. 2). Also depicted in Fig. 3 are the cumulative biliary excretion profiles for SA and SB conjugates during perfusions of TR- livers. Perfusate concentrations of silybin conjugates increased more rapidly during the first 20 min of the perfusion compared with that observed with WT IPRLs (see Fig. 2). Only 2.5 and 4% of the doses for SA and SB, respectively, were excreted into bile as conjugates in TR- IPRLs at 90 min compared with 21 and 32%, respectively, with WT IPRLs. In addition, the biliary excretion profiles for SC, SD, ISA, and ISB also were significantly reduced in TR- livers (data not shown).
For WT livers, 7.2 μg of SC, 2.6 μg of SD, 2.9 μg of SA, 3.5 μg of SB, 0.5 μg of ISA, and 0.3 μg of ISB were excreted unchanged into the bile at the end of the 90-min perfusion. Similar amounts of each flavonolignan also were excreted into the bile by TR- IPRLs, and no differences in the biliary clearances of the unconjugated flavonolignans between livers from WT and TR- rats were evident (Table 1). These data suggest that biliary excretion of the parent flavonolignans may not be mediated by Mrp2.

Cumulative excretion of conjugated (glucuronide + sulfate) silymarin flavonolignans in bile (A) and perfusate (B) at 90 min in isolated perfused livers from wild-type and TR- rats. Data are expressed in equivalents of the unconjugated silymarin flavonolignans as mean ± S.E.M. of n = 3 livers in each group; *, p < 0.05, WT versus TR-.
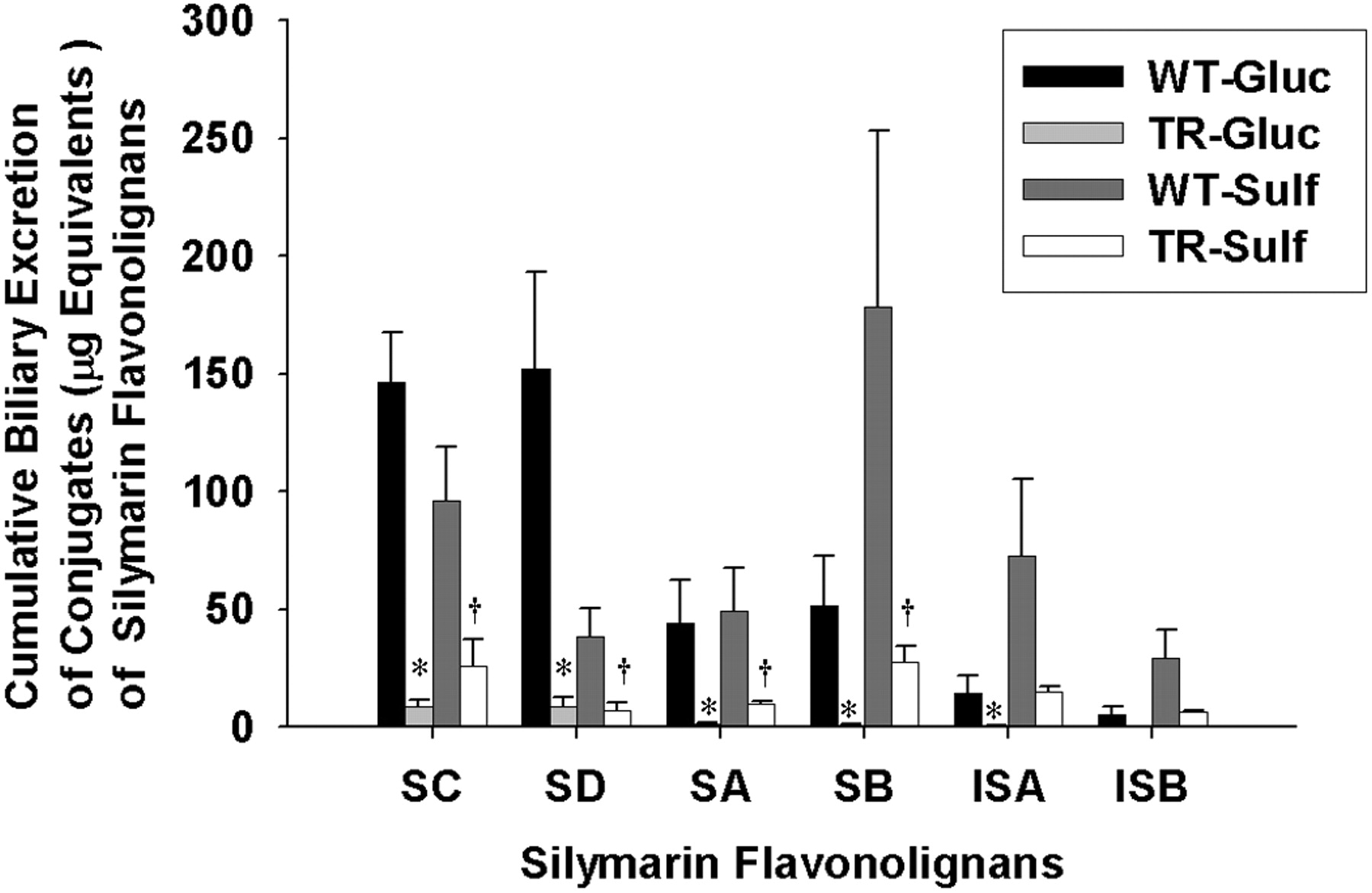
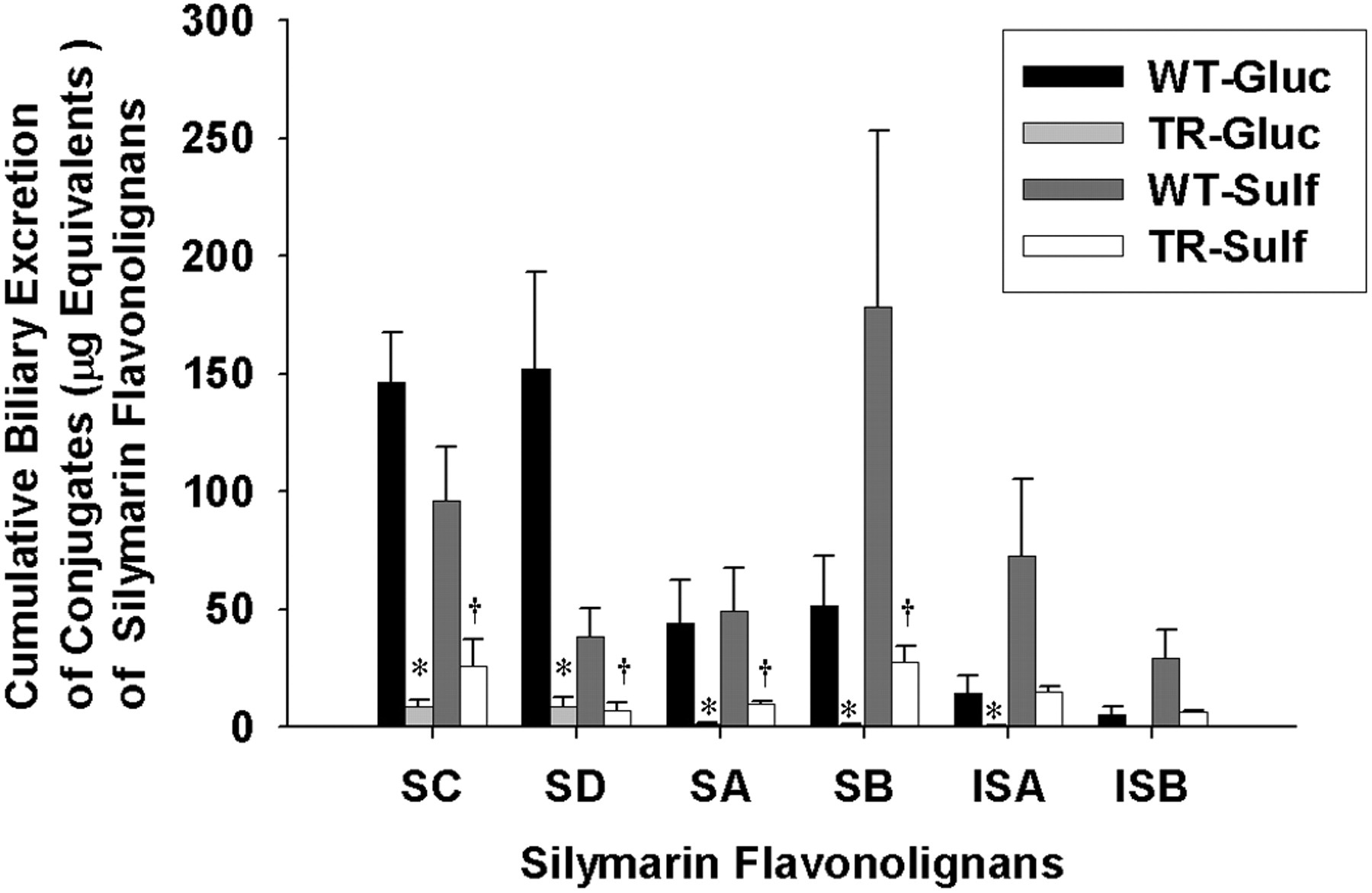
Cumulative biliary excretion of the glucuronide and sulfate conjugates of silymarin flavonolignans in isolated perfused livers from wild-type and TR- rats. Data are expressed in equivalents of unconjugated silymarin flavonolignans as mean ± S.E.M. of n = 3 livers in each group; *, p < 0.05, WT versus TR- for glucuronide conjugates; †, p < 0.05, WT versus TR- for sulfate conjugates.
Figure 4A depicts the cumulative percentage of the dose excreted in bile through 90 min as conjugates in WT and TR- IPRLs for each of the six silymarin flavonolignans. For WT livers, 242.7 μg of SC, 190.8 μg of SD, 93.1 μg of SA, 230.3 μg of SB, 86.9 μg of ISA, and 34.7 μg of ISB were excreted in the bile as conjugates at the end of the 90-min perfusion. Compared with WT IPRLs, the biliary excretion of flavonolignan conjugates was reduced by 80 to 92% in TR- IPRLs (SC, 86%; SD, 92%; SA, 88%; SB, 88%; ISA, 82%; and ISB, 80%). These results suggest that the biliary excretion of flavonolignan conjugates is primarily dependent on Mrp2. Figure 4B summarizes the percentage of the dose recovered as conjugates (glucuronide and sulfate) in the perfusate after 90 min in both WT and TR- IPRLs for each of the six silymarin flavonolignans. Among the flavonolignans, between 30 and 83% of the administered dose was excreted into the perfusate in TR- IPRLs compared with only 5 to 30% of the dose in WT IPRLs. SC conjugates exhibited the greatest increase in perfusate exposure in TR- IPRLs. However, the total amount of flavonolignan conjugates (glucuronides and sulfates) excreted in both bile and perfusate after 90 min was not different between TR- IPRLs (1552.8 ± 321.6 μg) and WT IPRLs (1227.1 ± 265.3 μg).
The total recovery of the 2.08-mg perfusion dose of silymarin flavonolignans ranged between 50 and 74% for WT IPRLs (mean 1.26 ± 0.26 mg) and between 64 and 91% (mean 1.60 ± 0.28 mg) for TR- IPRLs. Another 7 to 9% of the silymarin flavonolignan dose was recovered in three perfused livers that were examined (data not shown). These data suggest that the major effect of Mrp2 deficiency on the excretion of flavonolignan conjugates in rat IPRLs is a shift in excretion from bile to the perfusate.
Cumulative Biliary Excretion of Silymarin Glucuronide and Sulfate Conjugates. To determine whether the extent of biliary excretion of each silymarin flavonolignan was governed by the type of conjugation reaction, the cumulative biliary excretion of flavonolignan glucuronide and sulfate conjugates was determined. As shown in Fig. 5 in the WT IPRLs, the primary route of conjugation for SD was by glucuronidation (80.5%), whereas sulfation was the major pathway for SB (78%), ISA (84%), and ISB (85%). Approximately equal amounts of glucuronide and sulfate conjugates were recovered in bile for SA and SC. These data suggest that each silymarin flavonolignan has a preferred route of phase II metabolism. Also depicted in Fig. 5 is the cumulative biliary excretion of glucuronide and sulfate conjugates for each flavonolignan by TR- IPRLs, which were significantly reduced irrespective of the type of conjugation reaction. Compared with WT IPRLs, the amount of glucuronide and sulfate conjugates for each of the flavonolignans excreted into bile was reduced between 94 and 98% and 73 and 84%, respectively, in TR- IPRLs.
The apparent hepatobiliary clearances for the six flavonolignans are summarized in Table 2. Consistent with the data presented in Fig. 5, hepatobiliary clearances for both glucuronide and sulfate conjugates of each silymarin flavonolignan were markedly reduced by ∼89 to 99% and 75 to 90%, respectively, in TR- compared with wild-type IPRLs. The apparent hepatobiliary clearances for SD more closely reflect its formation clearance because this flavonolignan is almost completely excreted in the bile. Collectively, these data indicate for the first time a significant role of Mrp2 in biliary elimination of glucuronide and sulfate metabolites of each of the six silymarin flavonolignans.
Apparent hepatobiliary clearances of glucuronide and sulfate conjugates and total (glucuronide + sulfate) silymarin flavonolignans conjugates in WT versus TR− rats
Data are expressed as mean ± S.E.M. (n = 3). Apparent hepatobiliary clearance (Clb'app) is calculated as cumulative amount of conjugated flavonolignans excreted in bile (Ae) divided by the respective AUC0→90 min of the parent flavonolignan in the perfusate.
Discussion
In the present study, we compared the hepatobiliary disposition of silymarin using isolated perfused livers from WT and TR- rats. The initial perfusate concentration of total silymarin flavonolignans used in this study (26 μg/ml) was within the range of plasma concentrations for total flavonolignans attained in vivo with p.o. dosing. Peak concentrations of SA and SB between 0.06 μg/ml with silymarin doses of 200 mg/kg (Morazzoni et al., 1993) and 9.02 μg/ml with silipide doses of 200 mg/kg (Morazzoni et al., 1992, 1993) have been observed in pharmacokinetic studies in rats. Concentrations of SA and SB as high as 10 to 40 μg/ml have been observed in clinical studies (Flaig et al., 2007).
High first-pass phase II metabolism is believed to be the major factor responsible for the poor bioavailability of silymarin in humans because glucuronide conjugates account for approximately 90% of the total amount of SA and SB in blood following p.o. doses (Weyhenmeyer et al., 1992; Gatti and Perucca, 1994). In rats, glucuronide and sulfate conjugates accounted for 98% of SA and SB in blood and bile (Morazzoni et al., 1992, 1993). In humans, silymarin conjugates are excreted primarily into bile because less that 5% of the dose is excreted in urine (Weyhenmeyer et al., 1992). In addition, reduced clearance of silybin conjugates, consistent with impaired biliary excretion of conjugates, was observed in patients with extrahepatic biliary obstruction (Schandalik et al., 1992). In our study, all six silymarin flavonolignans were cleared rapidly from the perfusate, and up to 100% of silymarin flavonolignans were excreted as conjugates into bile at the end of the 90-min perfusion. Although between 70 and 85% of the total dose was recovered in perfusate, bile, and perfused liver, conclusions regarding mass balance in our study cannot be stated with certainty given the high variability observed with IPRL studies. It is possible that formation of additional metabolites may account for a portion of the dose because the silymarin concentration used in our study has been associated with the formation of cytochrome P450-catalyzed demethylated and hydroxylated products in vitro (Gunaratna and Zhang, 2003; Jančová et al., 2007).
Although the specific transport proteins involved in the elimination of silymarin and its conjugates have not been identified, previous studies have shown that silymarin has inhibitory effects on the active transport of drugs mediated by P-gp, Mrp1, Mrp4, and Mrp5 (Zhang and Morris, 2003; Nguyen et al., 2003; Lania-Pietrzak et al., 2005; Wu et al., 2005, 2008). However, there have been no direct investigations of the biliary excretion of silymarin and its conjugates, which could present a complex problem because each of the six major silymarin flavonolignans could have different affinities for the various hepatobiliary transport proteins. Mrp2 is a canalicular organic anion transport protein that is responsible for the biliary elimination of organic anions such as the anticancer drug methotrexate (Luo et al., 2007). Mrp2 also has been shown to be an important transport protein for the biliary excretion of polyphenolic natural products such as quercetin (van Zanden et al., 2007), resveratrol (Maier-Salamon et al., 2008), and 4-methylumbelliferone (Zamek-Gliszczynski et al., 2006a). To establish the dependence of silymarin flavonolignans on Mrp2 for biliary excretion, the present studies were conducted in isolated perfused livers from Mrp2-deficient TR- rats, which are naturally occurring mutants of the Wistar rat (Jansen et al., 1985; König et al., 1999). TR- rats have hereditary conjugated hyperbilirubinemia and exhibit elevated bile acid concentrations, reduced bile flow, up-regulation of basolateral transport protein Mrp3, and display impaired biliary excretion of most organic anions. Livers from TR- rats also exhibit negligible biliary excretion of glucuronide and glutathione conjugates, whereas biliary excretion of sulfate conjugates may only be partially impaired (Maier-Salamon et al., 2008). Our study using isolated perfused livers from TR- rats indicated that Mrp2 is the major canalicular transport protein involved in the biliary excretion silymarin flavonolignan conjugates. Although biliary excretion of the parent silymarin flavonolignans remained unchanged in the TR- and WT IPRLs, the rapid phase II metabolism to glucuronide and sulfate conjugates may limit their availability for active transport. For each parent flavonolignan, <5% of the dose was recovered in bile as unchanged flavonolignan at the end of each perfusion in both WT and TR- IPRLs, indicating that other canalicular transport proteins such as P-gp or breast cancer resistance protein may mediate high affinity transport.
The basolateral organic anion transport protein, Mrp3, is up-regulated as a compensatory mechanism for the absence of Mrp2 in TR- rats (Ogawa et al., 2000; Johnson et al., 2006). In TR- rats, increased basolateral efflux of methotrexate (Luo et al., 2007), gemfibrozil (Kim et al., 2003), acetaminophen glucuronide (Xiong et al., 2002), which are Mrp2 substrates, have been observed. During the first 30 min of the perfusion when parent flavonolignans were almost completely extracted from perfusate by both TR- and WT IPRLs, the rate of appearance of SA and SB conjugates in the perfusate was significantly greater for TR- compared with WT IPRLs. Although this increase in basolateral efflux of flavonolignan conjugates in TR- rats is consistent with an up-regulation of Mrp3, redirection of flavonolignan conjugates from canalicular to basolateral excretory pathways would be expected to occur in TR- rats as a result of the loss of Mrp2.
Information on the influence of phase II conjugation pathways on the biliary excretion of natural products is limited. Structural rather than stereo differences between the flavonolignans appeared to have the greatest influence on the extent of biliary elimination observed in our study. However, the finding that SD was the only silymarin flavonolignan that was primarily glucuronidated and almost completely excreted into bile suggests that flavonolignan glucuronide conjugates may be the preferred substrate for Mrp2. In humans, glucuronidation appears to be a preferred pathway for phase II metabolism of SB, SD, ISB, and SC (Wen et al., 2008).
In addition to Mrp2, other canalicular transport proteins may promote the biliary excretion of sulfate conjugates in rats (Zamek-Gliszczynski et al., 2005, 2006a). For example, the biliary excretion of glucuronide conjugates of resveratrol was decreased to a greater extent than the sulfate conjugates in TR- rats (Maier-Salamon et al., 2008). A similar trend was noted in the present study, where biliary excretion of glucuronide conjugates of each flavonolignan was reduced to a greater extent in TR- IPRLs than the biliary excretion of sulfate conjugates.
An important finding from our study was that the conjugates of SD were almost exclusively excreted into bile in the WT IPRLs, which suggests that SD may have the greatest potential to undergo enterohepatic recycling. In addition, the almost complete recovery of the dose for SD as glucuronide conjugates in the bile of WT IPRLs, which was reduced by ∼92% in TR- rat livers, suggests that this flavonolignan may be useful as a probe substrate for assessing changes in the functional activity of Mrp2. Many transport proteins of the ATP-binding cassette family such as MRP2 appear to be down-regulated in the livers of patients with chronic HCV (Hinoshita et al., 2001). In addition, whereas other studies have failed to show changes in expression detected by immunohistochemical techniques, there appears to be differences in the extent of recycling of canalicular transporters between the canalicular membrane and intracellular pools (Ros et al., 2003) in liver disease. Also, up-regulation of basolateral efflux transport proteins such as MRP3, MRP4, and MRP5 has been observed in liver diseases as an adaptation to the reduced expression of MRP2 (Ros et al., 2003; Barnes et al., 2007). Therefore, a specific MRP2 probe substrate would be useful for quantitating the extent of changes in functional activity irrespective of the expression of total MRP2 protein in diseased liver.
Given the results presented here, which indicate a primary role for Mrp2 in the biliary elimination of silymarin flavonolignan conjugates, it is likely that liver disease may be associated with down-regulation of MRP2 in humans because higher plasma concentrations of silymarin conjugates have been observed in patients with liver disease compared with healthy volunteers (Schrieber et al., 2008b). Similar changes in the disposition of SA and SB have been reported by Wu et al. (2008) using a rat model of cirrhosis where an approximately 2-fold increase in plasma AUC for SA and SB conjugates was correlated with a 50% reduction in the bile to blood exposure ratio for total SA and SB in cirrhotic rats compared with control. In addition, phase II metabolites of SC and SD may have greater specificity for Mrp2 because in this study their conjugates were the most extensively excreted into bile among the six major flavonolignans. A recent observation that among the six flavonolignans enterohepatic recycling was only observed for the conjugates of SC and SD in patients with chronic hepatitis C virus (Schrieber et al., 2008a) suggests that these two flavonolignans may be useful as surrogate markers of functional changes in MRP2 activity. In humans, decreased biliary excretion of flavonolignan conjugates may potentially influence the efficacy of silymarin because of down-regulation of MRP2 and reduced enterohepatic recycling and return of parent flavonolignans in portal blood.
In summary, these data indicate a primary role for Mrp2 in the biliary excretion of silymarin flavonolignan conjugates. Hepatic expression of MRP2 may be altered by chronic liver disease. The finding that SD is primarily glucuronidated and excreted into bile suggests that it might serve as a specific MRP2 probe substrate for quantitating functional changes in MRP2 activity resulting from liver disease. The extent to which alterations in MRP2-mediated biliary excretion of silymarin flavonolignans influence the antioxidant or antiviral activity of silymarin warrants investigation.
Footnotes
-
This work was supported in part by the National Institutes of Health (RO1 GM41935; to K.L.R.B.) and by Madaus GmbH.
-
doi:10.1124/dmd.108.021790.
-
ABBREVIATIONS: HCV, hepatitis C virus; SA, silybin A; SB, silybin B; ISA, isosilybin A; ISB, isosilybin B; SC, silychristin; SD, silydianin; P-gp, P-glycoprotein; Mrp, multidrug resistance-associated protein; IPRL, isolated perfused rat liver; TR-, Mrp2-deficient; WT, wild-type, Mrp2-competent; LC/ESI/MS, liquid chromatography/electrospray ionization/mass spectrometry; AUC0→90, area under the plasma concentration-time curve from time 0 to 90 min.
- Received April 9, 2008.
- Accepted August 4, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}