


Abstract
Antitumor 2′-deoxycytidine (dCyd) analogs such as gemcitabine (dFdC), cytarabine (Ara-C), and 2′-C-cyano-2′-deoxy-1-β-d-arabinofuranosylcytosine (CNDAC) are activated by dCyd kinase, whereas cytidine deaminase (CDA) inactivates them by conversion to their uracil forms. To elucidate the relationship between the chemosensitivity to antitumor dCyd nucleosides and CDA expression, we established a stable line of human gastric carcinoma TMK-1 cells constitutively overexpressing CDA (TMK-1/CDA) and examined its chemosensitivity to antitumor dCyd analogs in vitro and in vivo. We observed comparable reactivity for dFdC and Ara-C, and the substrate reactivity of CNDAC to recombinant human CDA was more than 10 times less efficient than those of Ara-C and dFdC. Next, we examined the in vitro chemosensitivity of TMK-1/CDA and observed a marked decrease in the sensitivity of TMK-1/CDA to Ara-C, dFdC, and CNDAC compared with mock-transfected cells. In addition, we transplanted TMK-1/CDA cells into a nude mouse xenograft model and examined their in vivo chemosensitivity to CNDAC. The in vivo antitumor effect of CNDAC on TMK-1/CDA cells was substantially reduced compared with that of mice transplanted with mock-transfected cells. These results indicate that CDA could play an important role in regulating susceptibility to antitumor dCyd analogs in vitro and in vivo. In addition, the expression level of CDA was found to affect the antitumor activity of CNDAC, even though the substrate reactivity of CNDAC to CDA is relatively low.
Introduction
Nucleoside analogs such as cytarabine (Ara-C) and gemcitabine (dFdC) are antimetabolic 2′-deoxycytidine (dCyd) analogs administered therapeutically to patients with malignant tumors (Fig. 1). The antitumor nucleoside 2′-C-cyano-2′-deoxy-1-β-d-arabinofuranosylcytosine (CNDAC) exerts potent antitumor activity against various human tumor cells in vitro and in vivo (Tanaka et al., 1992; Azuma et al., 1993) and is expected to have excellent clinical efficacy. Currently, a clinical phase study involving CNDAC is underway in the United States.

Structures of dCyd analogs.
CNDAC is transported into cells by nucleoside-specific transporters and monophosphorylated by dCyd kinase (dCK). Subsequently, it undergoes enzymatic phosphorylation and is converted to a triphosphate form. The triphosphate group of CNDAC inhibits DNA chain elongation after being incorporated into the DNA strand as a terminator and induces breakage of the DNA strand at its 3′-diester bond (Azuma et al., 1993; Matsuda and Sasaki, 2004). After incorporation into cells, dFdC and Ara-C undergo deamination by cytidine deaminase (CDA) and are inactivated by conversion to their uracil metabolites. In contrast, CNDAC is reported to be a poor substrate for mouse CDA, but the reactivity of CNDAC for human CDA has not been yet been investigated in detail.
Human CDA (EC 3.5.4.5) is a 146-amino acid protein (16.1 kDa) and generates Urd or dUrd by hydrolyzing the 4-amino group of Cyd and dCyd (Vincenzetti et al., 1996). In humans, CDA activity has been reported in tumor cells as well as in normal tissues, including the liver, spleen, intestinal mucosa, and lung, with particularly high activity in the liver and spleen (Camiener and Smith, 1965). The CDA gene is located on chromosome 1 (1p36.2–p35, 31 kilobases) and is composed of four exons (Saccone et al., 1994). Three single nucleotide polymorphisms have been previously reported at nucleotide position 79 (A to C), position 208 (G to A), and position 435 (T to C) in the amino acid-coding region of the human CDA gene (Kirch et al., 1998; Yue et al., 2003). Some studies have demonstrated that CDA gene polymorphism can affect susceptibility to antitumor dCyd analogs. However, there have been conflicting results (Sugiyama et al., 2007; Giovannetti et al., 2008; Tibaldi et al., 2008). Serum CDA activity was recently reported to correlate with toxicity after gemcitabine chemotherapy, but no genotype-to-phenotype relationships were observed (Ciccolini et al., 2010). However, the relationship between CDA activity and sensitivity to dFdC and CNDAC in cancer cells remains unclear.
In this study, we established a recombinant human CDA (rhCDA) expression system and compared the substrate reactivity of dCyd analogs, including dFdC and CNDAC, in rhCDA. In addition, we established a stable cell line overexpressing human CDA by introducing a cDNA expression vector into human solid tumor cells (gastric cancer TMK-1 cells) and compared their susceptibility to various types of antitumor dCyd analogs in vitro and in vivo to further explore the significance of CDA as a factor regulating susceptibility.
Materials and Methods
Animals.
We used 6-week-old female ICRnu/nu nude mice (Charles River Japan, Kanagawa, Japan). Animals were handled in accordance with the Guidelines for the Care and Use of Laboratory Animals of the Takara-machi Campus of Kanazawa University. The guidelines are also in accordance with Fundamental Guidelines for Proper Conduct of Animal Experiments and Related Activities in Academic Research Institutions under the Jurisdiction of the Ministry of Education, Culture, Sports, Science and Technology of Japan (Science Council of Japan, 1996).
Cell Culture and Materials.
Ara-C was purchased from Yamasa Shoyu (Choshi, Japan). CNDAC and gemcitabine were kindly provided by Taiho (Saitama, Japan). Anti-human CDA monoclonal antibody was produced using rhCDA as antigen. Human gastric tumor TMK-1, MKN-45, NUGC-3, and NKPS cells and human fibrosarcoma tumor HT-1080 cells were cultured in RPMI 1640 medium (Nissui Pharmaceutical, Osaka, Japan) containing 10% fetal bovine serum (Invitrogen, Carlsbad, CA) and 50 μg/ml kanamycin at 37°C in a humidified atmosphere.
Production of Human CDA Protein.
The full coding sequence of human CDA cDNA from HT-1080 cells was amplified by PCR using the sense primer 5′-GTCTAGGAGAGGAGCCGCAATCGTGTCT-3′ and antisense primer 5′-AAGTTGCTCGGAACAGGATAGAACC-3′. The CDA cDNA was cloned into a pcDNA4-His-Max TOPO vector (Invitrogen), which was then digested with KpnI and EcoRI. The fragment was inserted into the pRSET B vector (Invitrogen) and digested with KpnI and EcoRI. Finally, the CDA gene of the resulting plasmid pRSET B/CDA was sequenced to confirm the entire nucleotide sequence. The expression plasmid pRSET B/CDA was transformed into Escherichia coli BL21 and induced by adding isopropyl-β-d-thiogalactoside. rhCDA was purified using a Ni-NTA spin kit (QIAGEN, Valencia, CA) according to the manufacturer's instructions.
HPLC Analysis of CDA Activity.
CDA activity of cell lysate, serum, and protein extract samples was measured as described previously (Zhang et al., 1999). Sample was added to a reaction mixture (100 μl) containing 500 μM Cyd or antitumor dCyd analogs in potassium phosphate buffer (pH 7.4). The mixture was incubated at 37°C for 30 min, followed by HPLC analysis. The mixture was incubated at 95°C for 5 min to inactivate CDA. The mixture was then centrifuged at 15,000 rpm for 10 min, and the supernatant was applied to a reverse-phase chromatography column (CAPCELL PAK C18 UG120, ϕ4.6 × 250 mm; Shiseido, Tokyo, Japan). The elution was conducted at 0.5 ml/min using methanol-water (10:90), and Cyd and Urd were monitored at 254 nm. The retention time was approximately 7 min for Cyd and 9 min for Urd.
Establishment of CDA-Overexpressing Cancer Cell Line.
To establish a stable cell line overexpressing human CDA, the full coding sequence of human CDA cDNA was amplified by PCR using the sense primer 5′-GTCTAGGAGAGGAGCCGCAATCGTGTCT-3′ and antisense primer 5′-AAGTTGCTCGGAACAGGATAGAACC-3′. The CDA cDNA was cloned into a pcDNA4-myc-His A vector (Invitrogen) to construct pcDNA4/CDA. This plasmid was transfected into TMK-1 cells using FuGENE 6 transfection reagent (Roche Diagnostics, Indianapolis, IN) according to the manufacturer's instructions. The transfectants were isolated by culture in medium containing 40 μg/ml Zeocin, and a Zeocin-resistant clone, designated TMK-1/CDA, was established. The pcDNA4 empty vector alone was also transfected into TMK-1 cells to generate control clones, designated TMK-1/mock. CDA protein expression and CDA activity were examined by Western blotting and HPLC analysis.
Western Blot Analysis for Protein Expression of Human CDA.
Cell lysates were prepared in radioimmunoprecipitation assay buffer [50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, and 0.1% SDS]. Protein samples and serum samples were mixed with a loading buffer [50 mM Tris-HCl (pH 6.5), 10% glycerol, 2% SDS, 0.1% bromphenol blue, and 40 mM dithiothreitol]. After completion of electrophoresis using a 4 to 20% gradient SDS-polyacrylamide gel (e-PAGEL; ATTO, Tokyo, Japan), proteins were transferred to a polyvinylidene difluoride membrane filter (Immobilon; Millipore Corporation, Billerica, MA). The membrane was blocked in Tris-buffered saline-Tween containing 5% skim milk for 1 h and then probed with anti-human CDA monoclonal antibody for 2 h at room temperature. Horseradish peroxidase-conjugated anti-mouse IgG antibody and enhanced chemiluminescence detection reagent (GE Healthcare, Little Chalfont, Buckinghamshire, UK) were used for the detection of blots.
In Vitro Chemosensitivity.
In vitro cytotoxicity was examined using a modified tetrazolium-based semiautomatic colorimetric assay with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reagent (Sigma-Aldrich, St. Louis, MO) as described previously (Carmichael et al., 1987; Tanaka et al., 1992). Cancer cells were plated in a 96-well plate at a density of 5 × 103 cells/well. dCyd analogs were dissolved and diluted with distilled water for injection (Otsuka Pharmaceutical, Tokyo, Japan). Serial dilutions of dCyd analogs were added in each well, and the plate was incubated at 37°C for 72 h. After incubation, 25 μl of MTT solution (5 mg/ml in phosphate-buffered saline) was added in the culture medium, and the plate was incubated at 37°C for an additional 4 h. The cytotoxicity was calculated by applying the following formula: percent of cell growth inhibition = [1 − (T/C)] × 100, where C is the mean A540 of the control group and T is that of the treated group. The IC50 value was measured graphically from the dose-response curve with at least three drug concentration points.
In Vivo Chemosensitivity in Xenograft Model.
TMK-1/CDA or TMK-1/pcDNA4 cells (2 × 107) were subcutaneously transplanted into 6-week-old female ICRnu/nu nude mice on day 0. CNDAC was intraperitoneally administered at a dose of 7 mg/kg (5 day/week × 2) beginning on day 6. Tumor size was measured and tumor volume was calculated as tumor volume (cubic millimeters) = [length (millimeters) × width2 (square millimeters)]/2. The tumor volume was graphically plotted. After 28 days of tumor transplantation, blood samples were collected and analyzed for human CDA protein expression by Western blot analysis.
Statistical Analysis.
Data were analyzed with standard statistical methods. Student's t test was applied to determine the statistical significance of differences. Statistical probability was expressed as p < 0.05 and p < 0.01.
Results
Kinetic Parameters of rhCDA for Cyd and dCyd Analogs.
First, we determined the Km values of rhCDA for various nucleosides. The Km value for Cyd and dCyd was approximately 0.2 mM, which was lower than that for other dCyd analogs. Among the dCyd analogs, Km values were observed in the following order: Ara-C (0.299 mM) < dFdC (0.594 mM) < CNDAC (5.14 mM) (Table 1). Vmax/Km values, which indicate the efficiency of reactions, for the natural substrates Cyd and dCyd were higher than those of dCyd analogs. In addition, dFdC (0.0167) and Ara-C (0.0185) were found to have nearly the same reactivity; in comparison, the reaction efficiency of CNDAC (0.0014) to rhCDA was more than 10 times lower than those of Ara-C and dFdC.
Comparison of kinetic parameters of rhCDA for Cyd and 2′-dCyd analogs
To assess the kinetic parameters, we used a 100-μl reaction mixture containing 0.25 μg of recombinant human CDA and 0.25 to 3.75 mM Cyd or antitumor dCyd analogs in 20 mM potassium phosphate buffer (pH 7.4). The mixture was incubated at 37°C for 30 min, followed by HPLC analysis.
CDA Activity and Protein Expression in Transfected Cells.
To determine the significance of CDA as a factor regulating chemosensitivity, we introduced a human CDA expression vector into TMK-1 cells to generate cell lines stably overexpressing CDA. The expression of CDA protein in the transfected cells was analyzed by Western blot analysis, and we observed almost no CDA protein expression in TMK-1/mock cells (Fig. 2A). Conversely, strong CDA protein expression was observed in the cell lysates and conditioned medium of TMK-1/CDA cells transfected with the CDA expression vector. In addition, we investigated the time course of secretion of CDA protein in TMK-1/CDA cells. The CDA protein was found to be released into conditioned medium, and the amount of secreted CDA protein gradually increased from 4 to 24 h (Fig. 2B). Compared with TMK-1/mock cells, CDA activity was distinctly enhanced in TMK-1/CDA cells (Fig. 2C); therefore, we were able to confirm that the cells overexpressed active CDA. The CDA activity of lysate from TMK-1/CDA cells was 16.6 pmol/min (Fig. 2D).

Characterization of CDA-overexpressing cells. cDNA was cloned into the pcDNA4-myc-His A vector to construct pcDNA4/CDA. This plasmid was transfected into TMK-1 cells. The transfectant was designated TMK-1/CDA. Likewise, the pcDNA4 empty vector was transfected, and transfectants were designated TMK-1/mock. A, 10 μg of cell lysate and 15 μl of concentrated medium were electrophoresed on a 4 to 20% SDS-polyacrylamide gel. Conditioned medium was concentrated 10-fold using a gel filtration spin column. Western blot analysis of CDA protein was conducted using anti-human CDA monoclonal antibody. B, cells were plated at a density of 1 × 106 cells/35-mm dish. After 24 h, the medium was exchanged for serum-free medium, and the cells were incubated for 0, 4, 12, or 24 h. Ten micrograms of cell lysate, and 15 μl of concentrated medium were used for Western blot analysis. C and D, CDA activity of cell lysate was measured by high-performance liquid chromatography. The reaction mixture (100 μl) containing 10 μg/ml cell lysate and 0.5 mM Cyd in potassium phosphate buffer was incubated at 37°C for 30 min. CDA activity was determined by the amount of deaminated products (Urd) per min. The values represent means ± S.D.
Expression of CDA in Human Gastric Cancer Cell Lines.
We examined the expression of CDA protein in human gastric cancer NUGC-3, MKN-45, TMK-1, and NKPS cells by Western blot analysis, and CDA protein in cell lysate was detected in every cell line. Furthermore, MKN-45 and NKPS cells also released CDA into their culture medium in vitro as observed in TMK-1/CDA cells (Fig. 3).

Protein expression of CDA in human gastric cancer cell lines. Cells were plated at a density of 1 × 106 cells/6-cm dish. After 24 h, the medium was exchanged for serum-free medium, and the cells were incubated for 24 h. Conditioned medium was concentrated 150-fold using a gel filtration spin column. Ten micrograms of cell lysate and 15 μl of concentrated medium were used for Western blot analysis.
In Vitro Chemosensitivity of CDA-Overexpressing Cells.
We examined the cytotoxicity of dCyd analogs on TMK-1/CDA and on TMK-1/mock cells. Compared with TMK-1/mock cells, TMK-1/CDA cells showed a substantially reduced susceptibility to Ara-C, dFdC, and CNDAC (Fig. 4). The IC50 value for Ara-C was 108 times higher than that in TMK-1/mock cells, the IC50 value for dFdC was 63 times higher, and the IC50 value for CNDAC increased to 101 times higher. Resistance to antitumor dCyd analogs was confirmed in CDA-overexpressing cells (Table 2).
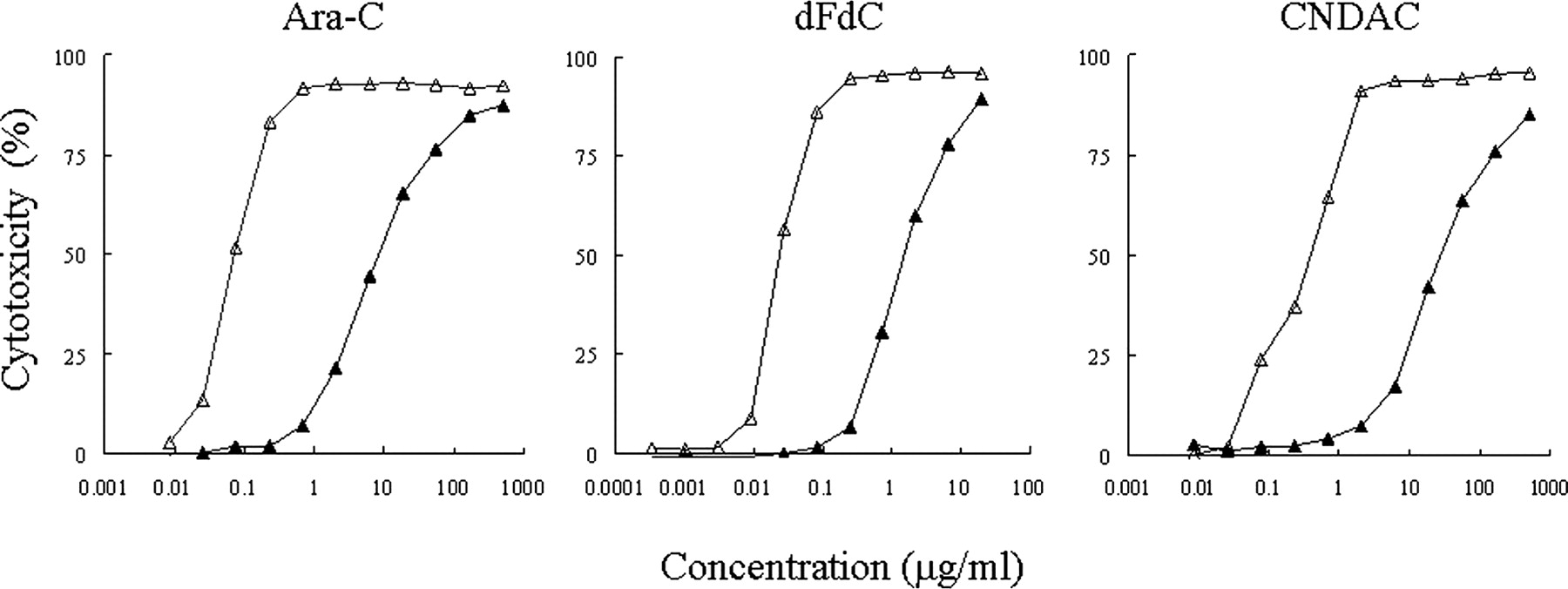
Chemosensitivity of TMK-1/CDA cells to dCyd analogs in vitro. Cells (5 × 103) were seeded onto 96-well plates with simultaneous addition of serial dilutions of nucleoside analog in triplicate, and cells were incubated at 37°C for 72 h. Cell viability was determined by MTT assay. TMK-1/CDA cells (▴) overexpressed CDA protein. TMK-1/mock cells (▵) were transfected with control vector.
Comparison of sensitivity of cell lines to 2′-dCyd analogs
Chemosensitivity of CDA-Overexpressing TMK-1 Cells in a Xenograft Model.
We investigated the chemosensitivity of TMK-1/CDA and TMK-1/mock cells to CNDAC in vivo using the xenograft model. CNDAC was administered intraperitoneally at a dose of 7 mg/kg (5 days/week × 2) beginning 6 days after transplantation. Figure 5 shows the tumor growth curve. CNDAC showed significant antitumor activity against TMK-1/mock cells. In contrast, no significant difference in tumor growth was observed between control and CNDAC-treated TMK-1/CDA cells. To confirm the presence of human CDA protein in the serum of mice transplanted with TMK-1/CDA cells, we performed Western blot analysis using a monoclonal antibody against human CDA. The transplanted TMK-1/CDA cells produced human CDA protein even 28 days after transplantation, but human CDA protein was not detectable in mice transplanted with TMK-1/mock cells (Fig. 6). Thus, the human CDA produced was found to be released into the serum of mice.

Chemosensitivity of TMK-1 cells overexpressing CDA to CNDAC in nude mice. TMK-1/mock cells (A) or TMK-1/CDA cells (B) (2 × 107 cells) were transplanted subcutaneously into nude mice. Treatment was started on day 6 after tumor cell transplantation. Mice received intraperitoneal injections of 7 mg/kg CNDAC, 5 days/week for 2 weeks. Tumor size was measured every other day. The values represent means ± S.E. *, p < 0.05; **, p < 0.01.

Western blot analysis of human CDA in the serum of nude mice transplanted with TMK-1/mock and TMK-1/CDA tumor cells. After 4 weeks of transplantation, blood samples were collected. Serum separated from blood (10 μl) was electrophoresed on a 4 to 20% SDS-polyacrylamide gel, and human CDA protein was detected using anti-human CDA monoclonal antibody. The molecular weight of the CDA protein is indicated at the right with an arrow. Blood samples from two mice in each group were used for the analysis.
Discussion
During the 1970s, several clinical studies were performed on the combined use of a cytidine deaminase inhibitor, tetrahydrouridine (THU), and Ara-C in patients with acute leukemia. It was reported that THU increased serum Ara-C levels, and the response rates achieved with the combination were comparable to those achieved with high-dose Ara-C. 5-Fluoro-2′-deoxycytidine (FdCyd) is a DNA methyltransferase inhibitor and is rapidly converted to its uracil form 5-fluoro-2′-deoxyuridine by CDA in vivo. A phase I study on FdCyd and THU was recently conducted in patients with advanced solid tumors to determine the maximum tolerated dose of FdCyd and to determine the methylation status of p16 and other genes relevant to neoplasias in the National Cancer Institute. CDA is considered not only a predictive marker of chemosensitivity to antitumor dCyd analogs but also a useful target for biochemical modulation of these analogs.
In this study, we established a stable human CDA-expressing cell line, conducted a comparative examination of its chemosensitivity to various antitumor dCyd analogs, and examined the relationship between CDA expression and chemosensitivity. First, we examined the substrate reactivity of antitumor dCyd analogs to rhCDA and found that dFdC and Ara-C had nearly the same reactivity, whereas the reactivity of CNDAC was more than 10 times lower. After adjustment of Vmax/Km = 1 for dFdC, its value was 0.90 for Ara-C and 0.08 for CNDAC. We then examined the cytotoxicity of 2′-dCyd analogs on cancer cells using TMK-1/CDA cells in vitro. CDA overexpression led to reduced susceptibility to antitumor dCyd analogs. However, no clear correlation was observed between the increased IC50 value and the degree of substrate reactivity to rhCDA. Ara-C and dFdC demonstrated comparable degrees of substrate reactivity, but the rate of decrease in the cytotoxicity against CDA-overexpressing cell lines was more pronounced with Ara-C. Although the reactivity of CNDAC to CDA was more than 10 times lower than that of other dCyd analogs, the rate of decrease in the cytotoxicity was relatively greater, as summarized in Table 2. The low rate of decrease in the cytotoxicity of dFdC is thought to be due its faster initial uptake into cells than that of Ara-C. Because it has a high affinity and substrate reactivity to dCK (Heinemann et al., 1988), it enters the cell rapidly, becomes phosphorylated, and is converted into gemcitabine triphosphate; thus, it is less exposed to intra- and extracellular CDA than Ara-C. However, the velocity of the uptake of [3H]CNDAC into tumor cells is less than one-tenth that of [3H]Ara-C (data not shown), and, furthermore, the substrate reactivity of CNDAC to dCK is also as low as one-twentieth (Uchida et al., 1999). Thus, CNDAC can be metabolized by CDA on many occasions inside and outside the cells because of its low efficiency for membrane transport and monophosphorylation, even if it has low substrate reactivity to CDA. These results suggest that the intracellular transport capability of nucleosidic transporters and the activities of both dCK and CDA are important factors that regulate susceptibility to 2′-dCyd analogs and that their reciprocal balance determines chemosensitivity.
Next, to determine the effect of the antitumor activity of 2′-dCyd analogs in vivo, we conducted a comparative study of the efficacy of CNDAC using a nude mouse xenograft model transplanted subcutaneously with TMK-1/CDA or TMK-1/mock cells. An apparent resistance to the antitumor effect of CNDAC was observed in the cells overexpressing CDA. Leukemic cells have been reported to release CDA into their culture medium in vitro, which subsequently influences their chemosensitivity to Ara-C (Kawai et al., 1994). In addition, in the treatment of leukemia, Ara-C is thought to be rapidly metabolized and inactivated by CDA on the basis of its presence in the liver and blood. In this study, CDA-overexpressing cells and some gastric cancer cell lines also released CDA into their culture medium in vitro. We further confirmed that human CDA protein was present in the serum of nude mice transplanted with TMK-1/CDA cells, indicating that the CDA released by tumor cells flows into the blood. As mentioned earlier, CNDAC has a low substrate reactivity to CDA. However, transport of CNDAC into cells is relatively slow, and thus in that environment, it is thought to be easily exposed to extracellular CDA, such as that in blood and extracellular spaces surrounding cancer tissue. In this study, the fact that TMK-1/CDA cells in vivo showed resistance to the antitumor effect of CNDAC may be not only due to the influence of intracellular CDA but also due to extracellularly secreted CDA.
The above-mentioned results indicate that CDA is an important determinant regulating sensitivity to antitumor dCyd analogs both in vitro and in vivo, which have a low substrate reactivity to CDA, and even for CNDAC. In addition, CDA was found to be released extracellularly in some solid tumors, and secreted CDA flows into the blood, suggesting that not only the intracellular but also the extracellular presence of CDA may influence the antitumor activity of 2′-dCyd analogs. Therefore, our findings warrant further investigation of CDA in a tumor tissue or in the blood of cancer patients to predict the susceptibility to dCyd analogs.
Footnotes
This research was supported in part by the Japan Society for the Promotion of Science [Grant-in-Aid for Young Scientists (B), 2004, 15790071 and Grant-in-Aid for Scientific Research (C), 2005–2006, 17590118].
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.110.034397.
-
ABBREVIATIONS:
- Ara-C
- cytarabine
- dFdC
- gemcitabine
- dCyd
- deoxycytidine
- CNDAC
- 2′-C-cyano-2′-deoxy-1-β-d-arabinofuranosylcytosine
- dCK
- deoxycytidine kinase
- CDA
- cytidine deaminase
- Urd
- uridine
- dUrd
- deoxyuridine
- Cyd
- cytidine
- rh
- recombinant human
- HPLC
- high-performance liquid chromatography
- MTT
- 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- THU
- tetrahydrouridine
- FdCyd
- 5-fluoro-2′-deoxycytidine.
- Received May 11, 2010.
- Accepted June 29, 2010.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}