


Abstract
Δ9-Tetrahydrocannabinol, cannabidiol (CBD), and cannabinol are the three major cannabinoids contained in marijuana, which are devoid of nitrogen atoms in their structures. In this study, we investigated the inhibitory effects of the major phytocannabinoids on the catalytic activity of human CYP2D6. These major cannabinoids inhibited the 3-[2-(N,N-diethyl-N-methylammonium)ethyl]-7-methoxy-4-methylcoumarin (AMMC) and dextromethorphan O-demethylase activities of recombinant CYP2D6 and pooled human liver microsomes in a concentration-dependent manner (IC50 = 4.01–24.9 μM), indicating the strongest inhibitory potency of CBD. However, these cannabinoids showed no or weak metabolism-dependent inhibition. CBD competitively inhibited the CYP2D6 activities with the apparent Ki values of 1.16 to 2.69 μM. To clarify the structural requirement for CBD-mediated CYP2D6 inhibition, effects of CBD-related compounds on the AMMC O-demethylase activity of recombinant CYP2D6 were examined. Olivetol (IC50 = 7.21 μM) inhibited CYP2D6 activity as potently as CBD did (IC50 = 6.52 μM), whereas d-limonene did not show any inhibitory effect. Pentylbenzene failed to inhibit CYP2D6 activity. Furthermore, neither monomethyl nor dimethyl ethers of CBD inhibited the activity. Cannabidivarin having a propyl side chain inhibited CYP2D6 activity; its inhibitory effect (IC50 = 10.2 μM) was less potent than that of CBD. On the other hand, orcinol and resorcinol showed lack of inhibition. The inhibitory effect of CBD on CYP2D6 activity was more potent than those of 16 compounds without nitrogen atoms tested, such as progesterone. These results indicated that CBD caused potent direct CYP2D6 inhibition, in which two phenolic hydroxyl groups and the pentyl side chain of CBD may play important roles.
Introduction
Marijuana is the most widely abused drug in the world. Marijuana use is a growing public health concern because of dependence, association with polysubstance use, potential harmful effects on brain function and several peripheral systems, and various health consequences (Hall and Degenhardt, 2009). Epidemiological studies reveal that many abusers of marijuana concurrently use other recreational drugs, such as methamphetamine and 3,4-methylenedioxymethamphetamine (MDMA) (Wu et al., 2006; Grov et al., 2009). These facts prompted us to investigate how marijuana and coadministered drugs interact one another.
Marijuana leaves contain at least 70 cannabinoids, which consist of carbon, hydrogen, and oxygen atoms (ElSohly and Slade, 2005). Major cannabinoids in marijuana are Δ9-tetrahydrocannabinol (Δ9-THC), cannabidiol (CBD), and cannabinol (CBN) (Fig. 1). The average contents of Δ9-THC, CBD, and CBN in dried plant preparations of marijuana confiscated from 1993 to 2008 in the United States are 4.5, 0.4, and 0.3%, respectively, although these contents vary widely (Mehmedic et al., 2010). In marijuana resin, which is commonly referred to as hashish, the mean contents of Δ9-THC, CBD, and CBN are 14.1, 2.5, and 1.9%, respectively (Mehmedic et al., 2010). Δ9-THC is a primary psychoactive component of marijuana, possessing various pharmacological effects such as catalepsy, hypothermia, anti-inflammation, and antinociception (Pertwee, 2008). CBD is not psychoactive but has several pharmacological effects including drug-induced sleep prolongation and antiepileptic, anxiolytic, and antiemetic actions (Mechoulam et al., 2002). CBN is thought to exert minimal pharmacological effects in the central nervous system.

Structures of three major phytocannabinoids.
Δ9-THC, CBD, and CBN have been reported to be extensively metabolized by human liver microsomes (HLMs) (Bornheim et al., 1992; Watanabe et al., 1995, 2006, 2007; Jiang et al., 2011). It has been demonstrated that CYP2C and CYP3A enzymes are predominantly involved in the primary metabolism of the phytocannabinoids in the liver microsomes. Δ9-THC and CBN are mainly oxidized at the 11-position by CYP2C9 and at the 8-position by CYP3A4 (Bornheim et al., 1992; Watanabe et al., 1995, 2007), whereas CBD is primarily oxidized at the 7-position by CYP2C19 and at the 6-position by CYP3A4 (Jiang et al., 2011). On the other hand, these major phytocannabinoids have been shown to inhibit cytochrome P450 (P450)-mediated drug oxidations (Jaeger et al., 1996; Yamaori et al., 2010, 2011a,b).
Some recreational drugs that are likely to be used concurrently with marijuana are substrates for P450 enzymes. In particular, stimulants (e.g., methamphetamine and MDMA) and designer drugs including 5-methoxy-N,N-diisopropyltryptamine are known to be mainly metabolized by CYP2D6 (Zhou et al., 2009). CYP2D6 is expressed predominantly in the liver, although the expression level is low (∼2% of total P450 content) (Shimada et al., 1994). CYP2D6 is responsible for the metabolism of a wide variety of drugs clinically used, such as dextromethorphan, debrisoquine, bufuralol, promethazine, and codeine (Zhou et al., 2009). These findings indicate that CYP2D6 is one of the most important P450 enzymes involved in hepatic drug metabolism. In addition, these findings imply the possibility of frequent occurrence of CYP2D6-targeted drug interactions. Thus, it is toxicologically important to clarify possible interactions of marijuana or its components, including cannabinoids, with CYP2D6 substrates. However, the inhibitory effects of the cannabinoids on CYP2D6 activity remain to be elucidated.
In this study, we examined the potency and mechanism of CYP2D6 inhibition by the three major phytocannabinoids (Δ9-THC, CBD, and CBN). We report herein that CBD is a potent competitive inhibitor for CYP2D6. Furthermore, our study suggests that two phenolic hydroxyl groups and the pentyl side chain of CBD may have structurally important roles in CYP2D6 inhibition.
Materials and Methods
Materials.
Δ9-THC, CBD, and CBN were isolated from cannabis leaves using the method previously reported (Aramaki et al., 1968). CBD-2′-monomethyl ether (CBDM) and CBD-2′,6′-dimethyl ether (CBDD) were prepared as described previously (Gohda et al., 1990). Purities of these cannabinoids were determined to be greater than 97% by gas chromatography, except for CBDD, or which the purity was 93% (Watanabe et al., 2005). Cannabidivarin (CBDV) was generously provided from Dr. Yukihiro Shoyama at Nagasaki International University (Sasebo, Japan). 3-[2-(N,N-Diethyl-N-methylammonium)ethyl]-7-methoxy-4-methylcoumarin (AMMC), 3-[2-(N,N-diethylamino)ethyl]-7-hydroxy-4-methylcoumarin (AHMC), microsomes from baculovirus-infected insect cells expressing CYP2D6 with NADPH-P450 reductase (Supersomes), and pooled HLMs were purchased from BD Gentest (Woburn, MA). Dextromethorphan hydrobromide monohydrate, dextrorphan-d-tartrate, resorcinol, acenaphthylene, benz[a]anthracene, benzo[a]pyrene, 1-methylnaphthalene, 2-methylnaphthalene, phenanthrene, and quinidine sulfate dihydrate were obtained from Wako Pure Chemicals (Osaka, Japan). Olivetol, d-limonene, pentylbenzene, progesterone, allopregnanolone, 17α-hydroxyprogesterone, 5α-dihydroprogesterone, acenaphthene, anthracene, fluoranthene, naphthalene, and pyrene were purchased from Sigma-Aldrich (St. Louis, MO). Orcinol and fluorene were obtained from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan). NADP, glucose 6-phosphate, and glucose 6-phosphate dehydrogenase were obtained from Oriental Yeast Co. Ltd. (Tokyo, Japan). Other chemicals and solvents used were of the highest quality that was commercially available.
Enzyme Assays.
AMMC O-demethylase activity was determined by using 96-well microtiter plates according to the instruction of BD Gentest. In brief, an incubation mixture consisted of recombinant CYP2D6 (2 pmol), AMMC, an NADPH-generating system (8.2 μM NADP, 0.41 mM glucose 6-phosphate, 0.41 mM magnesium chloride, and 1 unit/ml glucose 6-phosphate dehydrogenase), and 100 mM potassium phosphate buffer (pH 7.4) in a final volume of 200 μl. After prewarming at 37°C for 5 min, reactions were initiated by the addition of the NADPH-generating system. Fluorescence derived from formation of an O-demethylated metabolite of AMMC was recorded every 3 min for 30 min using FLUOstar OPTIMA (BMG Labtech, Offenburg, Germany) with excitation and emission filters at 390 and 460 nm, respectively. AHMC was used as an external standard for quantification of the O-demethylated metabolite of AMMC, i.e., 3-[2-(N,N-diethyl-N-methylammonium)ethyl]-7-hydroxy-4-methylcoumarin.
The activity of dextromethorphan O-demethylation was measured as reported previously (Yu and Haining, 2001) with minor modifications. Recombinant CYP2D6 (0.25 pmol) and HLMs (12.5 μg of protein) were used as enzyme sources. An incubation mixture consisted of an enzyme source, dextromethorphan, an NADPH-generating system (500 μM NADP, 10 mM glucose 6-phosphate, 10 mM magnesium chloride, and 1 unit/ml glucose 6-phosphate dehydrogenase), and 100 mM potassium phosphate buffer (pH 7.4) in a final volume of 200 μl. Incubations were performed at 37°C for 10 min and were terminated by adding 10 μl of 70% (w/v) perchloric acid. After removal of protein by centrifugation, 50 μl of the supernatant was subjected to a high-performance liquid chromatography (D7500 integrator, L-7100 pump, L-7200 autosampler, L-7300 column oven, and L-7485 fluorescence detector; Hitachi, Tokyo, Japan) equipped with a Mightysil RP-18 GP column (4.6 × 250 mm, 5 μm; Kanto Chemical, Tokyo, Japan). The mobile phase was the mixture of acetonitrile/methanol: 10 mM potassium phosphate buffer adjusted to pH 3.5 with phosphoric acid (200:160:630). Elution was performed at a flow rate of 1.0 ml/min. The formation of dextrorphan was monitored at an excitation of 280 nm and an emission of 310 nm.
To determine the kinetic parameters for AMMC and dextromethorphan O-demethylations by recombinant CYP2D6 and/or HLMs, AMMC at 0.24 to 15 μM or dextromethorphan at 0.15 to 24 μM were incubated with each enzyme source under the same conditions as mentioned above. In preliminary experiments, these reaction conditions were confirmed to ensure linear initial rates for the formation of an AMMC metabolite or dextrorphan. Data points were fitted to the Michaelis-Menten equation by nonlinear regression analysis with GraphPad Prism 5.02 (GraphPad Software Inc., San Diego, CA).
Inhibition Studies.
Recombinant CYP2D6 and HLMs were incubated with AMMC or dextromethorphan in the presence of test compounds including cannabinoids (up to 100 μM) under the same manner as described under Enzyme Assays. With the exception of quinidine that was dissolved in water, all compounds used were dissolved in dimethyl sulfoxide (DMSO) and added to the incubation mixture at a final DMSO concentration of ≤1%. DMSO (≤1%) in incubation mixtures caused less than 6% inhibition of AMMC and dextromethorphan O-demethylase activities under the current conditions. The IC50 values were calculated by nonlinear regression analysis with GraphPad Prism 5.02 (GraphPad Software Inc.) using the dose-response with variable slope function.
The effects of three or four different inhibitor concentrations on AMMC and dextromethorphan O-demethylations were examined at five substrate concentrations to characterize the enzyme kinetics for the inhibition of CYP2D6 by CBD and its structurally related compounds. The apparent Ki values and the mode of inhibition were determined by nonlinear regression analysis using eq. 1 for competitive inhibition with GraphPad Prism 5.02 (GraphPad Software Inc.):
 where v, Vmax, [S], Km, and [I] are the velocity, maximal velocity, substrate concentration, Michaelis constant, and inhibitor concentration, respectively. Akaike's information criterion was used as a measure of goodness of fit. The mode of inhibition was verified by visual inspection of Lineweaver-Burk plots of the enzyme kinetic data.
where v, Vmax, [S], Km, and [I] are the velocity, maximal velocity, substrate concentration, Michaelis constant, and inhibitor concentration, respectively. Akaike's information criterion was used as a measure of goodness of fit. The mode of inhibition was verified by visual inspection of Lineweaver-Burk plots of the enzyme kinetic data.
To identify potential metabolism-dependent inhibition of CYP2D6 by cannabinoids, the inhibition experiments were performed as described below. The preincubation mixture contained recombinant CYP2D6 (2 pmol), each major cannabinoid (2.5–50 μM), an NADPH-generating system (8.2 μM NADP, 0.41 mM glucose 6-phosphate, 0.41 mM magnesium chloride, and 1 unit/ml glucose 6-phosphate dehydrogenase), and 100 mM potassium phosphate buffer (pH 7.4) in a final volume of 180 μl. After prewarming at 37°C for 5 min, reactions were initiated by the addition of the NADPH-generating system. After a 20-min preincubation, 20 μl of AMMC solution was added to the preincubation mixture (final substrate concentration, 0.6 μM). Incubations were conducted under the same manner as described under Enzyme Assays for AMMC O-demethylase activity.
Prediction of In Vivo Drug Interactions for CYP2D6.
An estimate of in vivo inhibition potency was determined by the methods of Obach et al. (2006). The maximal unbound hepatic input concentration, Cmax, u, inlet, was determined using the following equation:

In eq. 2, Cmax is defined as the maximal systemic concentration, fu is the fraction unbound in the blood, D is the oral dose, Fa is the fraction of the oral dose absorbed, ka is the first-order absorption rate constant, and Qh is the hepatic blood flow. In the case of marijuana smoking, absorption from the gastrointestinal tract is not taken into account. Thus, the Cmax, u, inlet for inhaled cannabinoids is equal to fu · Cmax. The value of fu for cannabinoids is at most 0.05 because 95 to 99% of plasma THC is bound to plasma proteins, mainly lipoproteins (Grotenhermen, 2003). A ratio of the area under the curve (AUC) with inhibitor to control AUC could be estimated using the following equation:

In eq. 3, AUCinhibited is the area under the curve for a given substrate probe in the presence of an inhibitor, and AUCcontrol is the area under the curve for the same probe substrate without inhibitor. The fraction of metabolism of the probe substrate by CYP2D6 and the magnitude of the potency of the inhibitor are represented by fm(CYP2D6) and Ki, respectively. The value of unity was used for the fm(CYP2D6) of dextromethorphan/dextrorphan urinary ratio (Obach et al., 2006). Thus, a ratio of AUCinhibited to AUCcontrol of dextromethorphan is equal to 1 + [I]in vivo/Ki.
Results
Inhibition of CYP2D6 Activity by Major Phytocannabinoids.
To clarify the enzymatic characteristics of recombinant CYP2D6 and pooled HLMs used in this study toward AMMC and dextromethorphan O-demethylase activities, kinetic analysis was conducted with these enzyme sources. All of the reactions examined followed the Michaelis-Menten kinetics based on the Eadie-Hofstee plots (data not shown). The apparent Km values were 0.555 μM for AMMC O-demethylation by recombinant CYP2D6, 0.551 μM for dextromethorphan O-demethylation by recombinant CYP2D6, and 3.62 μM for dextromethorphan O-demethylation by HLMs.
Thus, the effects of Δ9-THC, CBD, and CBN on CYP2D6-mediated activity were investigated at substrate concentrations of 0.6 μM for recombinant CYP2D6 and 4 μM for pooled HLMs. All of the cannabinoids tested inhibited AMMC and dextromethorphan O-demethylase activities of recombinant CYP2D6 and HLMs in a concentration-dependent manner (Fig. 2). Among the cannabinoids tested, CBD most potently inhibited the CYP2D6 activities, showing the IC50 values of 4.01 to 6.52 μM (Table 1). In addition, the inhibitory effect of quinidine, a CYP2D6-selective inhibitor, on CYP2D6 activities was examined. The AMMC O-demethylase activity of recombinant CYP2D6 was strongly inhibited by quinidine with the IC50 value of 3.33 nM, and the dextromethorphan O-demethylase activity of recombinant CYP2D6 and HLMs was efficiently inhibited by quinidine with the IC50 values of 1.86 and 20.3 nM, respectively (data not shown). The inhibitory effects of quinidine on CYP2D6-mediated activity in this study were comparable to previous findings (Chauret et al., 2001; Madeira et al., 2004).

Effects of the major phytocannabinoids on AMMC and the dextromethorphan O-demethylase activities of CYP2D6 and HLMs. Recombinant CYP2D6 was incubated with 0.6 μM AMMC (A) or 0.6 μM dextromethorphan (B) in the presence of various amounts of Δ9-THC, CBD, and CBN. C, HLMs were incubated with 4 μM dextromethorphan in the presence of various amounts of Δ9-THC, CBD, and CBN. Each point is the mean of two determinations.
IC50 values of the major phytocannabinoids for inhibition of AMMC and dextromethorphan O-demethylase activities of recombinant CYP2D6 and HLMs
Kinetic Analysis for Inhibition of CYP2D6 Activity by CBD.
Kinetic analysis for the inhibition was performed to characterize the mode of inhibition of CYP2D6 activity by CBD. The cannabinoid competitively inhibited the CYP2D6 activities (Fig. 3). The apparent Ki values were 1.16 μM for AMMC O-demethylation by recombinant CYP2D6, 2.69 μM for dextromethorphan O-demethylation by recombinant CYP2D6, and 2.42 μM for dextromethorphan O-demethylation by HLMs.
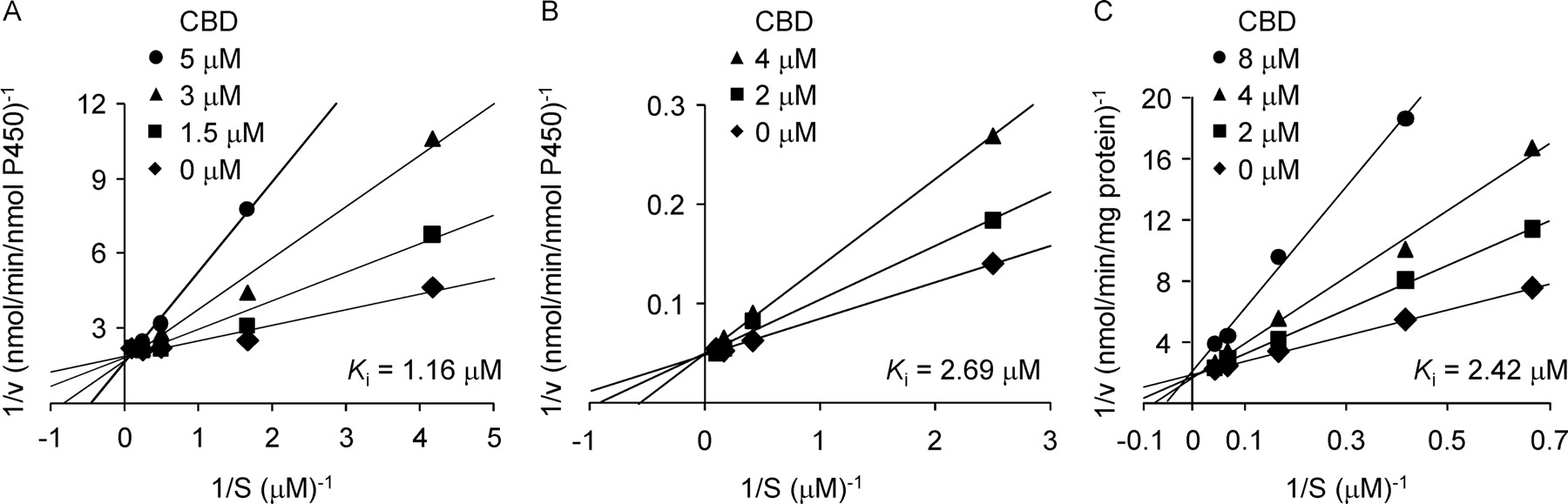
Lineweaver-Burk plots for the inhibition of CYP2D6 and HLMs by CBD. Recombinant CYP2D6 was incubated with AMMC (A) or dextromethorphan (B) in the presence or absence of CBD. C, HLMs were incubated with dextromethorphan in the presence or absence of CBD. Each point is the mean of duplicate determinations.
Structural Requirements for Inhibition of CYP2D6 Activity by CBD.
The inhibitory effects of CBD-related compounds on CYP2D6 activity were investigated to clarify structural requirements for the CYP2D6 inhibition by CBD (Fig. 4). In these inhibition studies, the activity of AMMC O-demethylation by recombinant CYP2D6 was used as a marker of activity. CBD consists of the pentylresorcinol and terpene moieties, as shown in Fig. 1. To determine which moiety of CBD is important in the CYP2D6 inhibition, the effects of olivetol and d-limonene were examined. Olivetol, which corresponds to the pentylresorcinol moiety of CBD (Fig. 4A), inhibited the AMMC O-demethylase activity of recombinant CYP2D6 (Fig. 4B): its inhibitory effect (IC50 = 7.21 μM) was comparable to that of CBD (Table 1). On the other hand, d-limonene, which corresponds to the terpene moiety of CBD (Fig. 4A), showed no inhibitory effect (Fig. 4B).
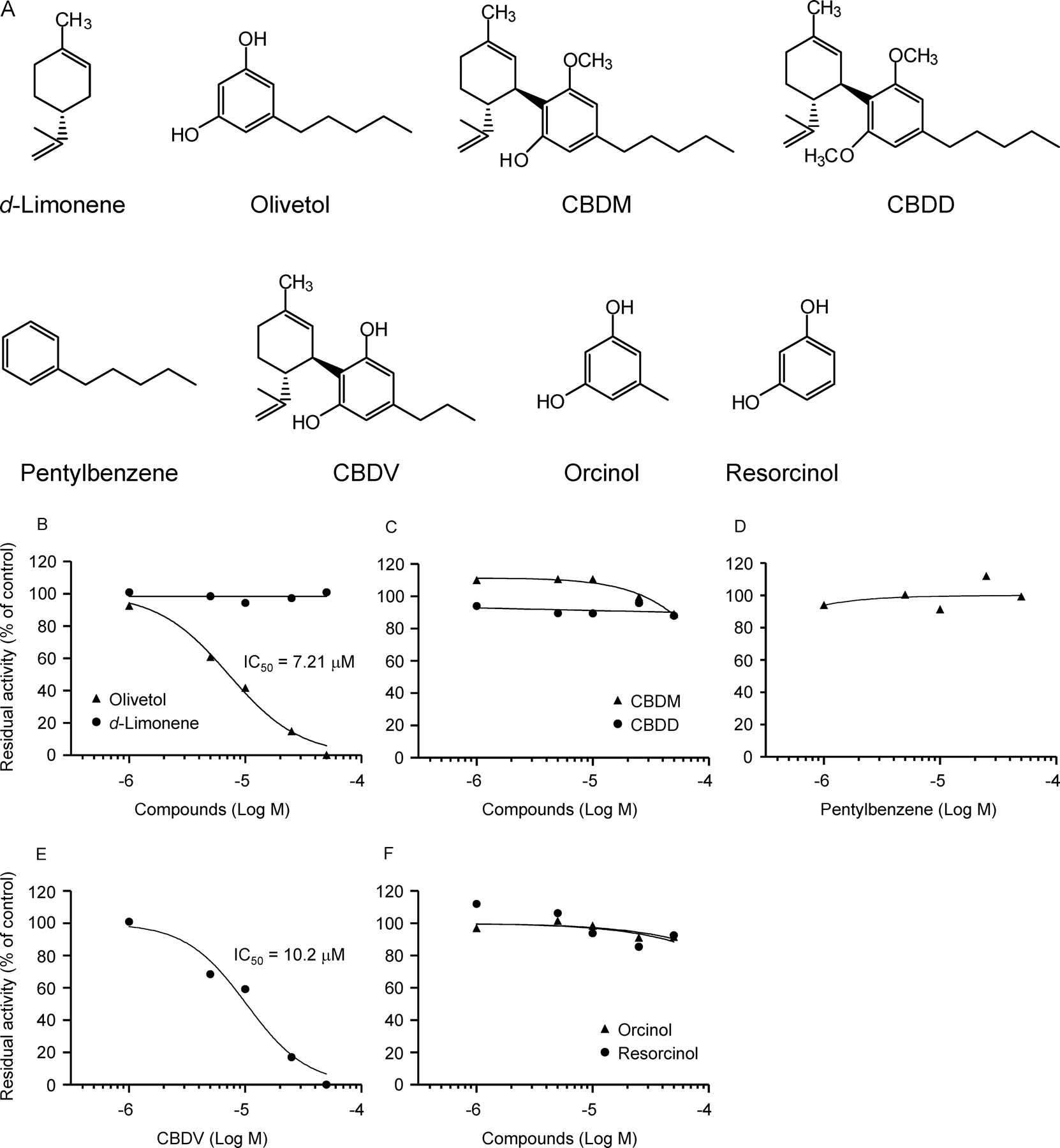
Effects of CBD-related compounds on the AMMC O-demethylase activity of CYP2D6. A, structures of CBD-related compounds. B to F, recombinant CYP2D6 was incubated with 0.6 μM AMMC in the presence of various amounts of CBD-related compounds: olivetol and d-limonene (B), CBDM and CBDD (C), pentylbenzene (D), CBDV (E), and orcinol and resorcinol (F). Each point is the mean of two determinations.
To determine whether the two free phenolic hydroxyl groups of CBD have a role in the inhibition of CYP2D6, the inhibitory effects of two methylated derivatives of CBD were investigated. The CYP2D6 activity was not inhibited by CBDM or CBDD (Fig. 4C), which are monomethylated and dimethylated derivatives of CBD, respectively (Fig. 4A). In addition, pentylbenzene, which is free of phenolic hydroxyl groups (Fig. 4A), did not inhibit the activity of CYP2D6 at all (Fig. 4D).
CBD and olivetol have a pentyl side chain. To elucidate the importance of the pentyl group of CBD in the inhibition against CYP2D6 activity, inhibition studies were conducted with CBDV, resorcinol, and orcinol. CBDV, possessing a shorter side chain (i.e., propyl group; Fig. 4A), inhibited the CYP2D6 activity to some extent (Fig. 4E), although its inhibitory effect (IC50 = 10.2 μM) was less potent than that of CBD (Table 1). On the other hand, resorcinol and its derivative orcinol, having a methyl group at the 5-position (Fig. 4A), did not exert any inhibitory effect (Fig. 4F).
Kinetic analysis for the inhibition was conducted to characterize the mode of inhibition of CYP2D6 activity by olivetol and CBDV. These compounds competitively inhibited the CYP2D6 activity (Fig. 5). The Ki values of olivetol and CBDV were 2.87 and 5.80 μM, respectively.

Lineweaver-Burk plots for inhibition of CYP2D6 by olivetol and CBDV. Recombinant CYP2D6 was incubated with AMMC in the presence or absence of olivetol and CBDV. Each point is the mean of duplicate determinations.
Effects of Endogenous Steroids and Marijuana Smoke Containing Polycyclic Aromatic Hydrocarbons on CYP2D6 Activity.
Endogenous steroids, polycyclic aromatic hydrocarbons (PAH), and cannabinoids do not have any basic nitrogen atom, the presence of which is a common characteristic of most CYP2D6 inhibitors. Nevertheless, some steroids have been reported to inhibit CYP2D6 activity (Hiroi et al., 2001). Thus, the inhibitory effects of endogenous steroids and the major PAH contained in marijuana smoke (Moir et al., 2008) on CYP2D6 activity were examined and compared with that of CBD. Progesterone inhibited the AMMC and dextromethorphan O-demethylase activities of recombinant CYP2D6 (Fig. 6), showing the IC50 values of 68.8 and 94.3 μM, respectively (Table 2). Allopregnanolone inhibited the AMMC O-demethylase activity of recombinant CYP2D6 in a concentration-dependent manner (IC50 = 44.6 μM) and in a competitive type (Ki = 35.2 μM). The other steroids tested, 17α-hydroxyprogesterone and 5α-dihydroprogesterone, exerted no inhibitory effect on the CYP2D6 activity under the current conditions. On the other hand, benz[a]anthracene, benzo[a]pyrene, and fluoranthene inhibited CYP2D6 activity in a concentration-dependent manner, exhibiting 37.9, 38.9, and 45.2%, respectively, inhibition at 100 μM. The other PAH examined—acenaphthene, acenaphthylene, anthracene, fluorene, 1-methylnaphthalene, 2-methylnaphthalene, naphthalene, phenanthrene, and pyrene—did not show significant inhibition (Fig. 6). These results revealed that, among the compounds tested without nitrogen atoms, CBD most potently inhibited CYP2D6 activity.

Effects of steroids and marijuana smoke containing PAH on CYP2D6 activities. A, recombinant CYP2D6 was incubated with 0.6 μM AMMC in the presence of various amounts of progesterone (P), allopregnanolone, 17α-hydroxy-P, and 5α-dihydro-P. B, recombinant CYP2D6 was incubated with 0.6 μM AMMC in the presence of various amounts of acenaphthene, acenaphthylene, fluorene, 1-methylnaphthalene, 2-methylnaphthalene, and naphthalene. C, recombinant CYP2D6 was incubated with 0.6 μM dextromethorphan in the presence of various amounts of P. D, recombinant CYP2D6 was incubated with 0.6 μM dextromethorphan in the presence of various amounts of anthracene, benz[a]anthracene, benzo[a]pyrene, fluoranthene, phenanthrene, and pyrene. Each point is the mean of two determinations.
Summary of kinetic parameters for inhibition of CYP2D6 activity by steroids and representative PAH contained in marijuana smoke
Metabolism-Dependent Inhibition of CYP2D6 Activity by Major Phytocannabinoids.
An effect of preincubation on inhibition by Δ9-THC, CBD, and CBN was investigated to determine whether these cannabinoids inhibit CYP2D6-mediated oxidation in a metabolism-dependent manner. A 20-min preincubation of Δ9-THC and CBN in the presence of NADPH did not increase the inhibition of the CYP2D6 activity (Table 3). The preincubation of CBD slightly potentiated the CYP2D6 inhibition.
Effect of preincubation on inhibition of CYP2D6 activity by major phytocannabinoids
All determinations were performed in duplicate.
Discussion
In this study, we demonstrated that Δ9-THC, CBD, and CBN, the three major cannabinoids in marijuana, inhibited the catalytic activity of human CYP2D6. Among the cannabinoids tested, CBD most potently inhibited CYP2D6 activity. However, the metabolism-dependent inhibition of CYP2D6 by CBD was less effective. We have recently reported that CYP2D6 is capable of metabolizing CBD, although the enzyme is not a major one responsible for CBD metabolism (Jiang et al., 2011). These results suggest that CBD itself may inhibit the activity of CYP2D6.
The structure of CBD contains the pentylresorcinol and terpene moieties. The fact that the inhibitory effect of olivetol on CYP2D6 activity was equal to that of CBD suggests that the pentylresorcinol moiety may play a critical role in the CBD-mediated CYP2D6 inhibition. The structure of the resorcinol moiety contains two phenolic hydroxyl groups and a phenyl ring, which are likely to interact with amino acids within the active site of CYP2D6. Then, we focused on the role of the phenolic hydroxyl groups of CBD in CYP2D6 inhibition. The lack of both phenolic hydroxyl groups in olivetol completely eliminated the inhibitory effect on CYP2D6 activity. Furthermore, the methylation of either hydroxyl group in CBD failed to inhibit CYP2D6 activity. A hydroxyl group at the 2′-position in CBD is equivalent to the hydroxyl group at the 6′-position because there is free rotation of the phenyl ring about the chemical bond at the 1′-position (Fig. 1). These results suggest that both free phenolic hydroxyl groups in the resorcinol moiety of CBD may be required for CYP2D6 inhibition.
Next, we focused on a role of the pentyl side chain of CBD in CYP2D6 inhibition. The substitution of the pentyl group of CBD to a propyl group attenuated the inhibitory effect on CYP2D6 activity. In addition, the side chain shortening to a methyl group resulted in the loss of CYP2D6 inhibition. In our recent study, recombinant CYP2D6 has been shown to oxidize CBD at the 6-, 7-, 4″-, and 5″-positions (Jiang et al., 2011). This finding suggests two possible orientations in which the terpene moiety or pentyl side chain of CBD are positioned near the heme in the CYP2D6 active site when the cannabinoid enters into the active site. When the terpene moiety in CBD is located close to the heme, the pentyl side chain in the cannabinoid is thought to be fixed within the proposed hydrophobic region corresponding to a toe area of the foot-shaped cavity in CYP2D6 (Rowland et al., 2006). On the other hand, when the pentyl side chain of CBD is positioned near the heme, the 4″- or 5″-position of the side chain is located above the heme, and the proximal portion of the side chain is considered to interact with the proposed hydrophobic region defined by hydrophobic residues, such as a Met residue at site 374 and Leu residues at sites 213 and 484 (de Graaf et al., 2007; Kotsuma et al., 2008). In both cases, the side-chain shortening of CBD may lead to unstable binding of the cannabinoid within the active site in CYP2D6 because of attenuation of the hydrophobic interactions. These results suggest that the pentyl side chain of CBD also plays an important role in CYP2D6 inhibition.
Certain structural characteristics common to typical CYP2D6 inhibitors are known (Strobl et al., 1993). One of these is to contain at least one basic nitrogen atom in structure. However, several CYP2D6 inhibitors do not have this characteristic. Some steroids such as progesterone and allopregnanolone are atypical inhibitors of CYP2D6 because they have no nitrogen atom in their structures (Hiroi et al., 2001). The three major phytocannabinoids (Δ9-THC, CBD, and CBN) and some PAH (benz[a]anthracene, benzo[a]pyrene, and fluoranthene), which inhibited CYP2D6 activity in this study, have no nitrogen atom in their structures. Therefore, these compounds would be categorized as atypical CYP2D6 inhibitors. The most potent inhibition reported so far of CYP2D6 by compounds without a nitrogen atom was observed in progesterone, with a Ki value of approximately 33 μM (Hiroi et al., 2001). In this study, CYP2D6-mediated activity was most potently inhibited by CBD among the major phytocannabinoids, marijuana smoke containing PAH, and endogenous steroids examined. The Ki values of CBD were 1.16 to 2.69 μM, indicating that the inhibitory effect of CBD on CYP2D6 activity is much stronger than that of progesterone. Compared with typical CYP2D6 inhibitors, the inhibitory effect of CBD on recombinant CYP2D6-mediated activity is comparable to those of (S)-nolfluoxetine (Ki = 0.84 μM), (S)-fluoxetine (Ki = 1.27 μM) (Shen et al., 2007), desipramine (Ki = 1.0 μM), and imipramine (Ki = 1.4 μM) (Niwa et al., 2008). The inhibitory effect of CBD on HLM-mediated CYP2D6 activity is as potent as those of (S,R)-fluoxetine (Ki = 1.6 μM), desipramine (Ki = 1.7 μM) (Ball et al., 1997), and 3,4-methylenedioxyamphetamine (Ki = 1.8 μM) (Wu et al., 1997). In addition, the potency of CBD against CYP2D6 inhibition in HLMs is slightly lower than that of MDMA (Ki = 0.6 μM), but it is much higher than those of (+)-methamphetamine and (−)-amphetamine (Ki ≈ 25 μM) (Wu et al., 1997). These results indicate that CBD is a potent inhibitor for CYP2D6.
Marijuana is commonly consumed by smoking. A clinical study has shown that a peak level of plasma concentration of Δ9-THC is 162 ng/ml (0.516 μM) after smoking a single marijuana cigarette containing 34 mg Δ9-THC, the content of which is 3.55% (Huestis et al., 1992). The content of CBD is higher in marijuana resin (hashish) than in a dried plant preparation of marijuana (Mehmedic et al., 2010). The plasma level of CBD after smoking a placebo marijuana cigarette spiked with 20 mg of the cannabinoid has been reported to be 114 ng/ml (0.363 μM) (Ohlsson et al., 1984). This plasma profile has been shown to be similar to that of Δ9-THC administered under a similar experimental design (Ohlsson et al., 1980). When the in vivo inhibition potency of CBD against CYP2D6 is determined from the plasma concentration of CBD described above and the Ki values of CBD for dextromethorphan O-demethylation (≈2.5 μM) by the methods of Obach et al. (2006), the ratio of AUCinhibited to AUCcontrol is estimated to be 1.01. Thus, CBD is not anticipated to exhibit in vivo inhibition of CYP2D6. However, we cannot exclude the possibility that the in vivo inhibition potency of CBD is underestimated because there is an example that the predicted ratios of AUCinhibited with fluoxetine to the AUCcontrol of CYP2D6 substrates are 1.1 to 1.5, albeit the observed ratios are 3.3 to 7.4 (McGinnity et al., 2008). To our knowledge, there is no report on the in vivo drug interactions of cannabinoids with CYP2D6 substrates such as methamphetamine and MDMA. Because marijuana and these recreational drugs are regulated as illegal drugs in most countries, it is thought to be difficult to obtain information about drug interactions among these illegal drugs. Therefore, further investigations are required to clarify in vivo CYP2D6-targeted drug interactions with marijuana or cannabinoids.
In conclusion, we demonstrated that CBD, devoid of a nitrogen atom, is characterized as a potent competitive inhibitor of CYP2D6. Our results suggest that two free phenolic hydroxyl groups in the resorcinol moiety and the pentyl side chain of CBD may be required for CYP2D6 inhibition.
Authorship Contributions
Participated in research design: Yamaori, Okamoto, Yamamoto, and Watanabe.
Conducted experiments: Yamaori and Okamoto.
Performed data analysis: Yamaori and Okamoto.
Wrote or contributed to the writing of the manuscript: Yamaori, Yamamoto, and Watanabe.
Acknowledgments
We thank Dr. Yukihiro Shoyama (Nagasaki International University, Sasebo, Japan) for generously providing CBDV.
Footnotes
This work was supported in part by the Ministry of Education, Culture, Sports, Science, and Technology of Japan [Grant-in-Aid for Scientific Research (C) 20590127 and Grant-in-Aid for Young Scientists (B) 23790159]; and the Academic Frontier Project for Private Universities from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (2005–2009).
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.111.041384.
-
ABBREVIATIONS:
- MDMA
- 3,4-methylenedioxymethamphetamine
- AMMC
- 3-[2-(N,N-diethyl-N-methylammonium)ethyl]-7-methoxy-4-methyl-coumarin
- AHMC
- 3-[2-(N,N-diethylamino)ethyl]-7-hydroxy-4-methylcoumarin
- CBD
- cannabidiol
- P450
- cytochrome P450
- CBDM
- CBD-2′-monomethyl ether
- CBDD
- CBD-2′,6′-dimethyl ether
- CBDV
- cannabidivarin
- CBN
- cannabinol
- DMSO
- dimethyl sulfoxide
- HLMs
- human liver microsomes
- PAH
- polycyclic aromatic hydrocarbon
- Δ9-THC
- Δ9-tetrahydrocannabinol
- AUC
- area under the curve.
- Received June 26, 2011.
- Accepted August 5, 2011.
- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}