


Abstract
Gastrointestinal toxicity, such as late-onset diarrhea, is a significant concern in irinotecan hydrochloride (CPT-11)–containing regimens. Prophylaxis of late-onset diarrhea has been reported with use of Japanese traditional (Kampo) medicine containing baicalin and with the antibiotic cefixime, and this has been explained in terms of inhibition of bacterial deconjugation of SN-38-glucuronide since unconjugated SN-38 (active metabolite of CPT-11) is responsible for the gastrointestinal toxicity. It is also prerequisite for SN-38 to be accumulated in intestinal tissues to exert toxicity. Based on the fact that liver-specific organic anion transporting polypeptide (OATP)1B1, a member of the same family as OATP2B1, is known to be involved in hepatic transport of SN-38, we hypothesized that intestinal transporter OATP2B1 contributes to the accumulation of SN-38 in gastrointestinal tissues, and its inhibition would help prevent associated toxicity. We found that uptake of SN-38 by OATP2B1-expressing Xenopus oocytes was significantly higher than that by control oocytes. OATP2B1-mediated uptake of SN-38 was saturable, pH dependent, and decreased in the presence of baicalin, cefixime, or fruit juices such as apple juice. In vivo gastrointestinal toxicity of SN-38 in mice caused by oral administration for consecutive 5 days was prevented by coingestion of apple juice. Thus, OATP2B1 contributes to the uptake of SN-38 by intestinal tissues, triggering gastrointestinal toxicity. So, in addition to the reported inhibition of bacterial β-glucuronidase by cefixime or baicalin, inhibition of OATP2B1 may also contribute to prevention of gastrointestinal toxicity. Apple juice may be helpful for prophylaxis of late-onset diarrhea observed in CPT-11 therapy without disturbance of the intestinal microflora.
Introduction
Irinotecan hydrochloride (CPT-11) is widely used in cancer chemotherapy. Its active metabolite, SN-38 (7-ethyl-10-hydroxycamptothecin), is formed mainly in the liver. The pharmacologic/toxicologic effect of CPT-11 is ascribed mostly to SN-38, which is approximately a thousand times more potent than the parent compound in inhibiting topoisomerase I (Chabot, 1997). SN-38 is further metabolized to an inactive glucuronic acid conjugate (SN-38G) in the liver (Di Paolo et al., 2006). SN-38G is excreted mainly into bile via transporters such as the breast cancer resistance protein, multidrug resistance–associated protein, and P-glycoprotein (Sugiyama et al., 1998; Di Paolo et al., 2006), and renal clearance accounts for only 30% of total SN-38 clearance (Slatter et al., 2000); however, SN-38G secreted in bile is hydrolyzed to SN-38 by β-glucuronidase in intestinal microflora, and the resulting SN-38 causes gastrointestinal toxicity. Since one of the dose-limiting toxicities of CPT-11 is late-onset diarrhea, it is important to reduce SN-38–induced gastrointestinal toxicity. A Japanese traditional (Kampo) medicine (containing baicalin as an active ingredient) and antibiotics such as cefixime for pediatric patients are clinically effective to ameliorate gastrointestinal toxicity since they inactivate bacterial β-glucuronidase, thereby reducing formation of toxic SN-38 from SN-38G (Furman et al., 2006; McGregor et al., 2012). In addition, we showed that SN-38 is a substrate of hepatic uptake transporter organic anion transporting protein (OATP)1B1 (Nozawa et al., 2005). Also, a genetic variant of OATP1B1 with reduced transport activity is associated with increased incidence of life-threatening toxicities resulting from increased systemic exposure to SN-38 (Xiang et al., 2006). Accordingly, both drug transporters and drug-metabolizing enzymes likely contribute to the pharmacologic and toxicologic actions of CPT-11.
Another OATP family member, OATP2B1, is expressed in several tissues, including the liver and intestine (Tamai et al., 2000). In the small intestine, OATP2B1 is located at the apical membrane of enterocytes and mediates pH-dependent transport of drugs (Kobayashi et al., 2003; Nozawa et al., 2004; Gröer et al., 2013), contributing to intestinal absorption of drugs (Tamai, 2012). It was reported that the plasma concentration of fexofenadine after oral administration was decreased by coingestion with fruit juices, presumably owing to inhibition of intestinal uptake (Dresser et al., 2002). We suggested a significant contribution of OATP2B1 to intestinal absorption of fexofenadine and proposed that this transporter is a site of interaction with fruit juice components (Imanaga et al., 2011; Shirasaka et al., 2013, 2014). OATP1B1 and 1B3 have broad substrate selectivity and mediate hepatic uptake of various drugs (Shitara et al., 2013). OATP2B1 also exhibits broad substrate selectivity, probably because of multiple binding sites (Shirasaka et al., 2012), as in the case of OATP1B1 (Tamai et al., 2001; Noe et al., 2007). Furthermore, SN-38 is transported by OATP1B1, which exhibits similar transport characteristics to OATP2B1 (Nozawa et al., 2005). Accordingly, it is possible that OATP2B1 contributes to the uptake of SN-38 by enterocytes.
The mechanism of CPT-11-induced late-onset diarrhea has not been fully clarified; however, it is believed that SN-38 formed in intestinal lumen is accumulated in intestinal tissues, where it exerts cytotoxicity. Indeed, it has been shown that intestinal cellular accumulation of SN-38 is positively correlated with cellular toxicity (Kobayashi et al., 1999; Ikegami et al., 2002). In addition, SN-38 was reported to be accumulated in Caco-2 cells via a transporter-mediated mechanism that is inhibited by baicalin (Itoh et al., 2005). Furthermore, cefixime, which is used to prevent gastrointestinal toxicity induced by CPT-11 in pediatric cancer patients, may interact with OATP2B1 since β-lactam antibiotics are mostly substrates of OATP transporters (Tamai et al., 2000; Nakakariya et al., 2008). Although baicalin and cefixime are effective to ameliorate gastrointestinal toxicity of SN-38 by inhibiting bacterial β-glucuronidase, it is also possible to explain their beneficial effect in terms of decreased accumulation of SN-38 in intestinal tissues attributable to inhibition of uptake transport, at least in part. In addition, it is known that the activity of OATP2B1 is decreased in the presence of fruit juices such as grapefruit juice, apple juice and orange juice (Shirasaka et al., 2013). In particular, apple juice inhibits OATP2B1 in a competitive and long-lasting manner, suggesting that it may serve as a potent inhibitor of intestinal OATP2B1. Accordingly, if OATP2B1 plays a significant role in intestinal epithelial tissue accumulation of SN-38, fruit juices could be useful to ameliorate gastrointestinal toxicity; this would be useful for patients since they would not have to take additional drugs to avoid gastrointestinal toxicity.
In the present study, we examined the in vitro transport of SN-38 by OATP2B1 and its relevance to gastrointestinal toxicity caused by SN-38. Based on the results, we propose a novel strategy to alleviate late-onset diarrhea in CPT-11 chemotherapy.
Materials and Methods
Materials.
SN-38 was purchased from Abcam (Cambridge, MA). Baicalin and gentamicin were purchased from Wako Pure Chemical Industries Ltd. (Osaka, Japan). Cefixime was gift from Astellas Pharma Inc. (Osaka, Japan). Apple, grapefruit, and orange juices (Tropicana; 100% pure at normal strength) were purchased at a local supermarket in Kanazawa, Japan. All other compounds and general reagents were obtained from Wako Pure Chemical Industries Ltd., Sigma-Aldrich (St. Louis, MO), or Kanto Chemicals Co., Inc. (Tokyo, Japan).
Transport Studies in Xenopus Oocytes Expressing OATP2B1.
Xenopus laevis frogs were purchased from Kato-S-Science (Chiba, Japan). Preparation of oocytes, in vitro synthesis of OATP2B1 cRNA, and uptake experiments were conducted as described previously (Nozawa et al., 2004; Shirasaka et al., 2010). Xenopus oocytes were used for evaluation of SN-38 transport by OATP2B1, and the accumulated SN-38 was quantitated by high-performance liquid chromatography (HPLC). In brief, the construct pGEMHE containing OATP2B1 cDNA was used to synthesize cRNA in vitro using mMESSAGE mMACHINE T7 (Ambion Inc., Austin, TX). For standard experiments, defolliculated oocytes were injected with 50 nl of the cRNA solution (0.5 μg/μl) or water and then incubated for 3 days at 18°C in modified Barth’s solution (MBS, pH 7.4) (88 mM NaCl, 1 mM KCl, 2.4 mM NaHCO3, 0.82 mM MgSO4, 0.33 mM Ca(NO3)2, 0.41 mM CaCl2, and 10 mM HEPES, pH 7.4, adjusted by NaOH) containing 50 μg/ml gentamicin.
For the uptake studies, oocytes expressing OATP2B1 were transferred to a 24-well culture plate and preincubated in MBS, pH 7.4, for 10 minutes at 25°C. To initiate the uptake reaction, the preincubated buffer was completely replaced with uptake buffer [MBS, pH 6.5 adjusted with 2-(N-morpholino)ethanesulfonic acid and NaOH] containing SN-38, and the oocytes were incubated for the designated time at 25°C. Here, before initiation of uptake reaction, SN-38 was dissolved in 50 mM K2HPO4 buffer (pH 9.0) or KH2PO4 buffer (pH 3.0) overnight to prepare carboxylate or lactone form of SN-38, respectively. Our laboratory has reported that a long-lasting inhibition of OATP2B1 was observed when cells were preincubated with apple juice (Shirasaka et al., 2013); however, the assessment of long-lasting inhibition of uptake of SN-38 was beyond the scope of the current work. Accordingly, for the inhibition studies, inhibitors were included only during uptake reaction. In uptake study at pH 8.0, the pH of the uptake buffer was adjusted with 10 mM HEPES and NaOH. Uptake was terminated by washing the oocytes three times with ice-cold MBS. To confirm stable expression of OATP2B1 in Xenopus oocytes, uptake of estrone-3-sulfate (a reference substrate of OATP2B1) was measured in each lot of Xenopus oocytes (data not shown). Only Xenopus oocytes from lots that showed appropriate activity were used for evaluation of SN-38 transport.
Measurement of SN-38 by HPLC.
For quantitation of SN-38 taken up by the oocytes, oocytes were homogenized by sonication in 500 μl of 50 mM KH2PO4 buffer (pH 3). The homogenate was mixed with methanol to make a 50% final concentration, and 50 μl of 10 nM camptothecin (internal standard) was added. After centrifugation at 15,000 rpm for 15 minutes at 4°C, 400 μl of the supernatant was dried under reduced pressure, and the residue was dissolved in mobile phase for HPLC analysis. Total (lactone and carboxylate forms) concentration of SN-38 was determined by HPLC with fluorescence detection. A 20-μl sample was injected into an HPLC system (JASCO LC-2000 plus series; JASCO, Tokyo, Japan). The analytical column was a Mightysil RP-18 GP column (250 mm × 4.6 mm; Kanto Chemical Co.) maintained at 40°C, with a mobile phase consisting of a mixture of 50 mM KH2PO4 (pH 2.5) and acetonitrile at the ratio of 70:30 v/v. The flow rate of the mobile phase was 1.0 ml/min. The excitation and emission wavelengths of the fluorescence detector were set at 375 and 560 nm, respectively.
Animal Study.
All animal experimentation was carried out in accordance with the guidance of the Kanazawa University Institutional Animal Care and Use Committee. Female mice initially weighing 30–50 g (C57BL6, Japan SLC, Hamamatsu, Japan) were housed three to five per cage with free access to water and a commercial animal chow throughout the acclimatization and experimental periods; they were maintained on a 12-hour dark/light cycle in an air-controlled room (temperature, 24.0 ± 1°C; humidity, 55 ± 5%).
The mice were orally given SN-38 dissolved in water at a dose of 10 mg/kg in a volume of 10 ml/kg, once daily for 5 days. In addition, the mice were given apple juice at 100% strength (10 ml/kg) or water every 12 hours through a gastric tube. The mice were sacrificed by cervical dislocation, and the entire intestinal tissue was removed. Intestinal tissue slices of each mouse were prepared using the Swiss-roll technique, and the samples were fixed with 4% paraformaldehyde in phosphate buffer (23 mM NaH2CO4, 77 mM Na2HPO4, pH 7.4) overnight at 4°C. After washing with phosphate buffer, the blocks of tissue were transferred sequentially to 70%, 80%, 90%, 95%, 99.5%, and 100% ethanol solution for about 1 hour each and then placed in a second 100% ethanol solution to ensure that all water was removed. The fixed tissue preparations were embedded in paraffin and cut into serial 4-μm slices. Paraffin sections were deparaffinized with four 3-minute xylene baths, rehydrated in successive 3-minute baths of ethanol (100%, 100%, 95%, 90%, 80%, and 70%), and one of distilled water, and then incubated for 30 seconds in Mayer hematoxylin solution (Merck KGaA, Darmstadt, Germany). The sections were rinsed thoroughly with distilled water, and eosin solution (Muto Pure Chemicals Co. Ltd., Tokyo, Japan) was applied for 10 seconds; then the sections were transferred sequentially to 70%, 80%, 90%, and 100% ethanol solution for about 1 hour each. They were then placed in a second 100% ethanol solution to ensure that all water was removed, followed by rinsing three times in xylene baths for 3 minutes. Finally, the sections were mounted on slides using VectMount (Vector Laboratories Inc., Burlingame, CA). Villus length was measured using a BZ-9000 microscope (BIOREVO; Keyence, Osaka, Japan).
Statistical Analysis.
Uptake is shown as the amount of SN-38 taken up (pmol/oocyte) or the cell-to-medium ratio (μl/oocyte) obtained by dividing the uptake amount by the initial concentration of SN-38 in the uptake medium. OATP2B1-mediated uptake was obtained after subtraction of the uptake by water-injected oocytes from that by OATP2B1 cRNA-injected oocytes. Kinetic parameters were estimated by means of nonlinear least-squares analysis using the KaleidaGraph program (Synergy Software, Reading, PA). The inhibitor concentration (I) giving half-maximum inhibition (IC50) was obtained by applying eq. 1:
 (1)
(1)The Michaelis-Menten constant Km and maximum uptake rate Vmax for uptake of SN-38 by OATP2B1 were obtained by fitting the data to the following eq. 2: (2)where v and C are uptake rate and concentration of SN-38, respectively.
(2)where v and C are uptake rate and concentration of SN-38, respectively.
Data are given as the mean of values obtained in at least three experiments with the S.E.M. Statistical analyses were performed using Student’s t test. A difference between the means was considered significant when the two-tailed value was less than 0.05.
Results
Characterization of OATP2B1-Mediated Uptake of SN-38.
Uptake of SN-38 carboxylate form by Xenopus oocytes injected with OATP2B1 cRNA or water as a control was measured. The time courses of uptake of SN-38 carboxylate form (0.1 μM) over 1 hour at pH 6.5 are shown in Fig. 1. The amount of SN-38 taken up by OATP2B1-expressing oocytes increased time dependently. SN-38 uptake by water-injected oocytes also increased time dependently but was much less than that in the case of OATP2B1-expressing oocytes (P < 0.05). When uptake of SN-38 lactone form was evaluated, higher uptake by OATP2B1-expressing oocytes than that by water-injected oocytes was similarly observed, with typical uptake values at 10 minutes of 1.15 ± 0.13 and 0.624 ± 0.066 μl/oocyte, respectively. These results show that OATP2B1 transports both carboxylate and lactone forms of SN-38 as substrates.

Time courses of uptake of SN-38 by OATP2B1. Uptake of SN-38 (carboxylate form, 0.1 μM) by Xenopus oocytes expressing OATP2B1 (filled circle) or water-injected oocytes (open circle) over 60 minutes was measured at pH 6.5. Each result represents the mean ± S.E.M. (n = 3–8). *A significant difference from the uptake by water-injected oocytes (P < 0.05).
pH Dependence of OATP2B1-Mediated SN-38 Uptake.
It is known that OATP2B1 exhibits higher uptake activity at acidic pH than neutral pH for certain substrates (Kobayashi et al., 2003). In addition, SN-38 has two forms—carboxylate and lactone forms—whose ratio is pH dependent. In solution at pH 3.0, SN-38 is present mostly in lactone form, whereas it is mostly in carboxylate form at pH 9.0 (Fassberg and Stella 1992). Therefore, we examined the preferred form of SN-38 transported by OATP2B1 and the pH dependence of OATP2B1-mediated uptake of SN-38 by preincubating SN-38 either at pH 3.0 and 9.0 overnight and by incubating SN-38 with oocytes either at pH 6.5 or 8.0 during the uptake reaction, respectively. The results are summarized in Table 1. Here, all uptake reactions were conducted for 10 minutes to minimize the shift of equilibrium between carboxylate and lactone forms during the uptake reaction. Uptakes by water-injected control oocytes and OATP2B1-expressing oocytes were both higher when SN-38 had been preincubated at pH 3.0 rather than at pH 9.0. SN-38 uptake also tended to be higher when OATP2B1-expressing oocytes were preincubated at pH 3.0 compared with pH 9.0. These results may indicate that the increased membrane permeability of the lactone form is due to both increased transport activity and increased diffusion rate. In most cases, uptake measured at pH 6.5 was higher than that measured at pH 8.0, demonstrating that OATP2B1 transport activity is pH-dependent. Diffusional transport is also pH dependent, with higher permeability at weakly acid pH. Overall, the results indicate that the lactone and carboxylate forms of SN-38 are both transported by OATP2B1, and acidic intestinal microclimate pH may facilitate cellular uptake of SN-38 in vivo since acidic pH appears to be preferable for both OATP2B1-mediated transport and diffusion.
pH Dependence on SN-38 uptake by OATP2B1 in Xenopus oocytes
For evaluation of lactone and carboxylate forms of SN-38, SN-38 (0.1 μM) was preincubated in phosphate buffer at pH 3.0 or 9.0, respectively, overnight before the uptake measurement. Then uptake by Xenopus oocytes was measured at pH 6.5 or 8.0 for 10 minutes. Uptake by water-injected oocytes, OATP2B1-expressing oocytes, and the OATP2B1-mediated uptake (shown as “Difference” in the table) was obtained subtraction of the former from the latter and are each shown as the mean ± S.E.M. (n = 7–10).
Concentration Dependence of SN-38 Transport by OATP2B1.
The concentration dependence of initial uptake of SN-38 carboxylate form by Xenopus oocytes expressing OATP2B1 was assessed in the range of 0.05 to 5 μM by measuring uptake at 30 minutes. The results are shown in Fig. 2. The uptake at pH 6.5 was saturable, and estimated kinetic parameters Km and Vmax were 0.763 ± 0.292 μM (mean ± S.E.M.), and 0.557 ± 0.070 pmol/30 minutes·oocyte, respectively. Similarly, Km and Vmax of the lactone form of SN-38 were estimated to be 1.26 ± 0.975 μM and 0.549 ± 0.140 pmol/10 minutes per oocyte at pH 6.5, respectively (data not shown). It should be noted that the obtained parameters may not strictly represent the transport characteristics of lactone and carboxylate forms of SN-38 since the ratio of lactone and carboxylate forms would be gradually changed by mutual conversion during uptake studies, depending on the pH of the solution (Fassberg and Stella, 1992).

Concentration dependence of OATP2B1-mediated uptake of SN-38. Uptake of SN-38 (carboxylate form) by OATP2B1-expressing Xenopus oocytes for 30 minutes was measured at pH 6.5. Data were corrected by subtracting the uptake by water-injected oocytes from that by OATP2B1 cRNA-injected oocytes. The line is the best-fit line calculated using the estimated values of Km and Vmax at pH 6.5. Each result represents the mean ± S.E.M. (n = 7–10).
Effects of Cefixime and Baicalin on OATP2B1-Mediated Uptake of SN-38.
The effects of cefixime and baicalin on OATP2B1-mediated uptake of carboxylate and lactone forms of SN-38 are shown in Fig. 3. Both cefixime and baicalin inhibited OATP2B1-mediated uptake of SN-38 in a concentration-dependent manner. The IC50 values of cefixime and baicalin for uptake of SN-38 carboxylate form were 2078 ± 1040 and 97.7 ± 22.3 μM, respectively, and those for lactone form of SN-38 were 1303 ± 389 and 9.41 ± 2.83 μM, respectively. Uptake of the lactone form of SN-38 appears to be more susceptible to OATP2B1 inhibitors than that of the carboxylate form.

Inhibitory effect of cefixime and baicalin on uptake of lactone or carboxylate form of SN-38 by OATP2B1. OATP2B1-mediated uptake of SN-38 (carboxylate form, 0.1 μM) was measured in the absence and presence of baicalin (A) or cefixime (B) for 10 minutes (lactone form: circle) or 30 minute (carboxylate form: triangle) at pH 6.5. Data were corrected by subtracting the uptake by water-injected oocytes from that by OATP2B1 cRNA-injected oocytes and are shown as % of control uptake. Each result represents the mean ± S.E.M. (n = 5–10).
Effect of Fruit Juices on OATP2B1-Mediated Uptake of SN-38.
The effect of fruit juices, including apple juice, grapefruit juice, and orange juice, on OATP2B1-mediated uptake of SN-38 was examined. As shown in Fig. 4A, each fruit juice at 20% of normal strength significantly reduced the uptake of both carboxylate and lactone forms of SN-38 by OATP2B1 (P < 0.05). To further assess the inhibitory characteristics of apple juice, SN-38 uptake was examined at increasing concentrations of apple juice from 0.2% to 40% (Fig. 4B). It caused a concentration-dependent reduction of SN-38 uptake, and IC50 values were 20.3 ± 3.67% and 6.93 ± 2.11% for the carboxylate and lactone forms of SN-38, respectively. OATP2B1-mediated transport of the lactone form of SN-38 is more susceptible to apple juice than that of the carboxylate form.
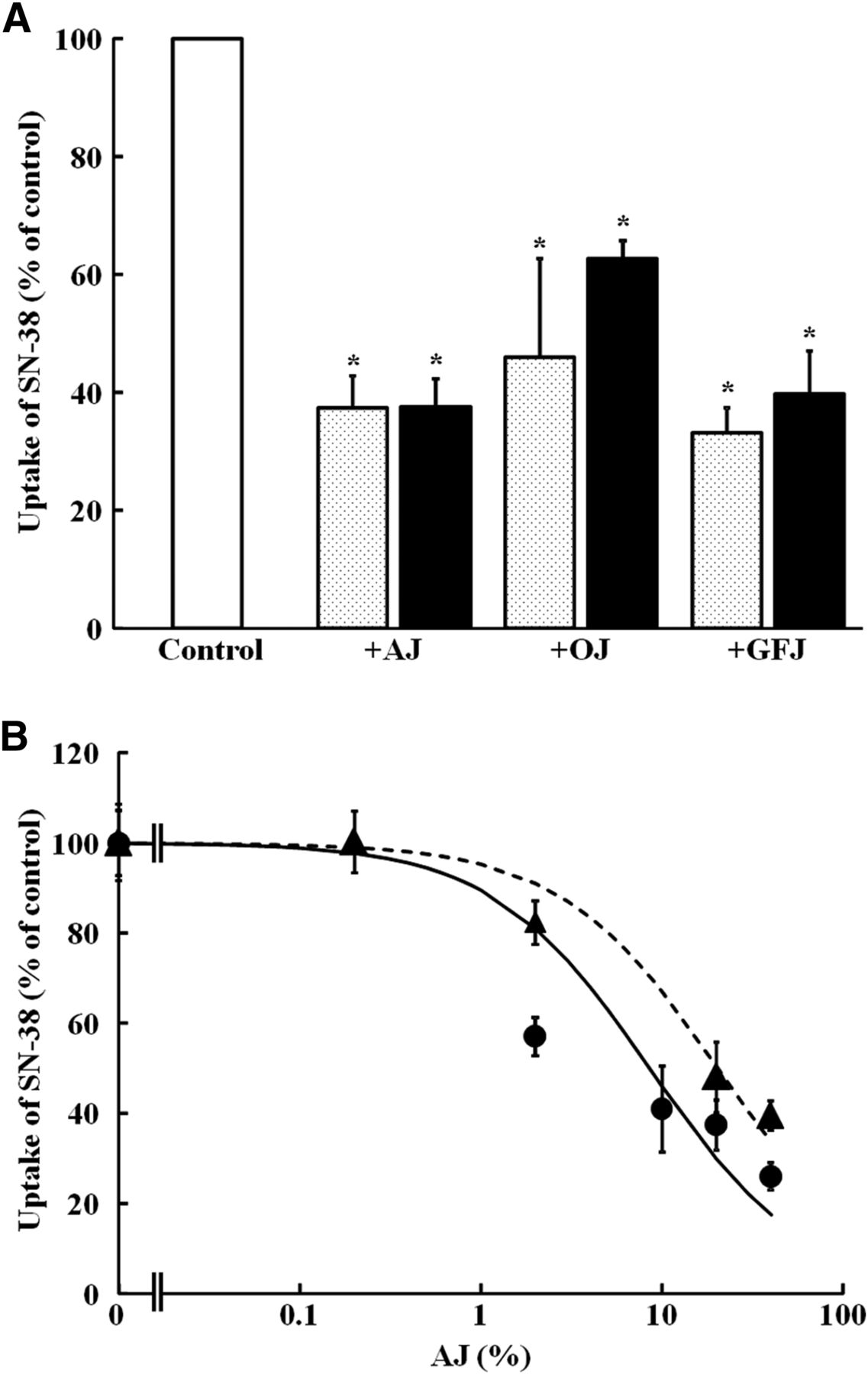
Inhibitory effect of apple juice (AJ), orange juice (OJ), and grapefruit juice (GFJ) on OATP2B1-mediated SN-38 uptake. (A) Uptake of lactone form (dotted column) and carboxylate form (closed column) of SN-38 (0.1 μM) by OATP2B1 was measured in the absence and presence of AJ, OJ, or GFJ at 20% dilution. (B) Concentration dependence of the inhibitory effect of AJ on OATP2B1-mediated uptake of lactone form (circle) or carboxylate form (triangle) of SN-38 (0.1 μM) was examined. Uptakes of lactone or carboxylate forms of SN-38 were measured for 10 minutes or 30 minutes, respectively. Data were corrected by subtracting the uptake by water-injected oocytes from that by OATP2B1 cRNA-injected oocytes. Each result represents the mean ± S.E.M. (n = 5–10).
Effect of Apple Juice on In Vivo Gastrointestinal Toxicity of SN-38 in Mice.
In vivo gastrointestinal toxicity of SN-38 in mice was examined by measuring the change in villus length after oral administration of 10 mg/kg of SN-38 once daily for 5 consecutive days via a stomach tube. Typical morphologic states are shown in Fig. 5A. At day 7, 2 days after termination of administration of SN-38, significant shortening of the intestinal villus was observed compared with the control (P < 0.05), suggesting that this phenomenon can be taken as a measure of late-onset diarrhea (Fig. 5B). SN-38 caused a shortening of villus length throughout the entire small intestine, to a minimum of 0.145 mm. When apple juice was ingested with SN-38, a small but statistically significant protective effect against SN-38-induced villus shortening was observed (Fig. 5B). Since apple juice contains potent OATP2B1 inhibitors, its observed effect can be explained in terms of reduced accumulation of SN-38 in the intestinal epithelial cells.
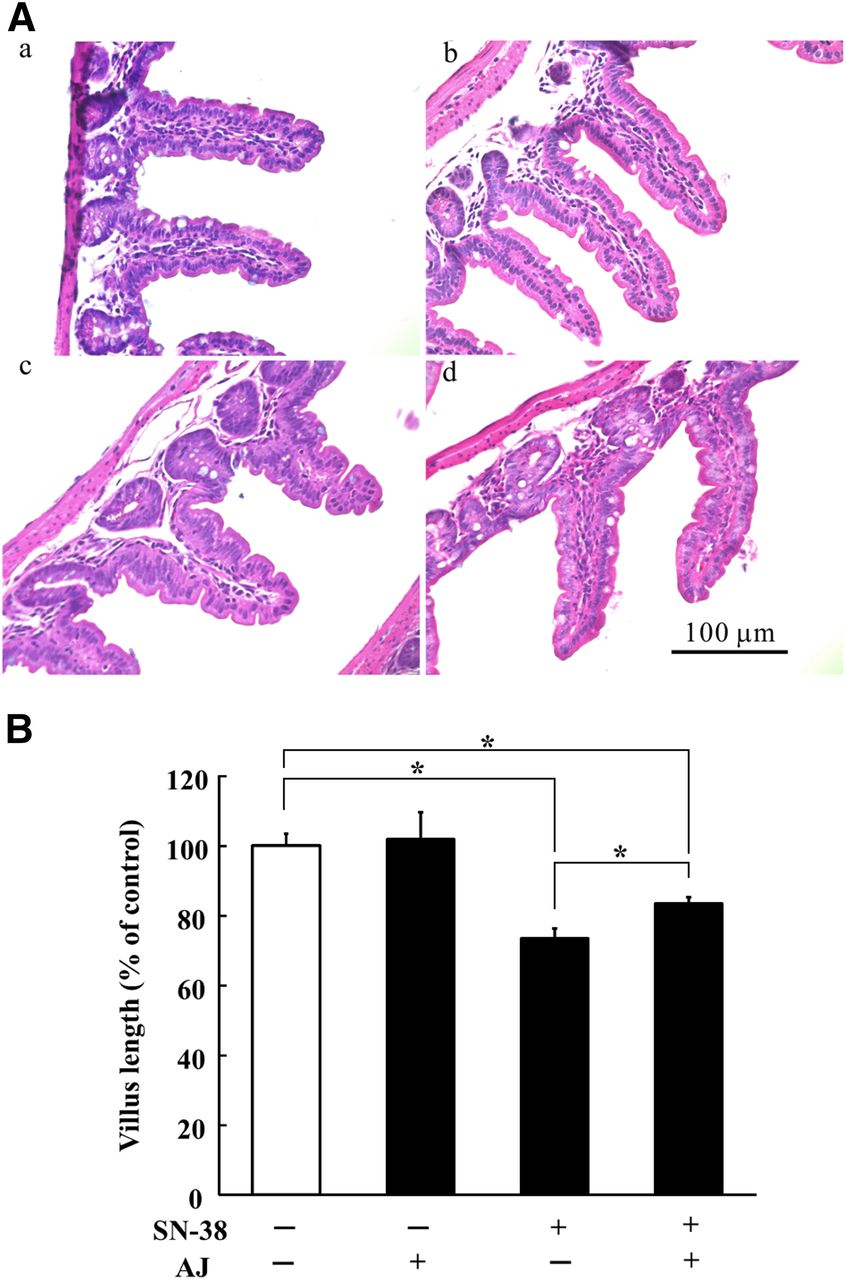
Protective effect of apple juice on SN-38–induced villus enterocyte injury in mice. SN-38 was orally administered at a dose of 10 mg/kg once daily for 5 consecutive days. Apple juice (AJ) at 100% strength (10 ml/kg) or water was orally administered every 12 hours. Two days after termination of SN-38 administration, intestinal tissue samples were prepared and the length of villi was measured under a microscope. (A) H&E staining of paraffin-embedded section of intestinal tissues from mice administered (a) vehicle only (control), (b) AJ only, (c) SN-38 only, and (d) both SN-38 and AJ. Experiments were repeated at least three times, and representative pictures are shown. (B) Lengths of villi of the intestinal tissues, with or without SN-38 or AJ, were measured. The data are shown as % of control length. Each column represents the mean and S.E.M. of 70–84 villi in four mice. *Significant difference between the pairs indicated by bars (P < 0.05).
Discussion
In this study, we examined the mechanism of accumulation of SN-38 in intestinal tissues since toxicity is considered to be dependent on the amount of the compound in the affected tissues. We hypothesized that accumulation of SN-38 in the intestinal tissue is a trigger of gastrointestinal toxicity and also that OATP2B1 is involved in SN-38 uptake from intestinal lumen since OATP2B1 exhibits a broad substrate selectivity and is responsible for intestinal absorption of its substrate drugs.
First, OATP2B1 clearly transported SN-38 when OATP2B1 was expressed in Xenopus oocytes (Fig. 1). SN-38 exists in a pH-dependent equilibrium of lactone and carboxylate forms owing to the labile α-hydroxy-3-lactone ring, which undergoes reversible hydrolysis at a rate that is dependent on pH (Fassberg and Stella, 1992). In addition, since pH affects the apparent transport activity of OATP2B1 (Kobayashi et al., 2003), we examined the effect of pH during preincubation and during the uptake process on the SN-38 uptake (Table 1). The results indicate that both the lactone and carboxylate forms of SN-38 are substrates of OATP2B1; the lactone form exhibited higher uptake at acidic pH, whereas uptake of the carboxylate form did not show clear pH dependence. The lactone form of SN-38 has higher membrane permeability, probably owing to simple diffusion (Kobayashi et al., 1999; Ikegami et al., 2002). Accordingly, the physiologically acidic intestinal luminal pH is favorable for the accumulation of SN-38 since the apparent uptake of SN-38 is the sum of transporter-mediated uptake and diffusion of both carboxylate and lactone forms.
Low Km values at pH 6.5 for uptake of the carboxylate and lactone forms of SN-38 by OATP2B1 (about 1 μM) suggest that SN-38 is a specific substrate of OATP2B1 and can be transported efficiently. Furthermore, these values are comparable with the value obtained for uptake of SN-38 by human colon-derived HT-29 cells (0.38 μM) (Ikegami et al., 2002). Thus, uptake in HT-29 cells may be mediated by OATP2B1. We and others have already demonstrated that SN-38 is a substrate of OATP1B1 (Nozawa et al., 2005) and OATP1B3 (Yamaguchi et al., 2008), which are clinically important liver-specific uptake transporters. Although OATP2B1 is also expressed in the liver (Tamai et al., 2000), OATP2B1 is likely to be pharmacologically more important in the intestine since OATP2B1, but not OATP1B1 or OATP1B3, is expressed in small-intestinal tissues (Tamai, 2012; Shirasaka et al., 2014). In our previous study, we failed to find a significant increase of SN-38 uptake by OATP2B1-expressing Xenopus oocytes (Nozawa et al., 2005). In addition, Fujita et al. did not observe significant transport of SN-38 by HEK293 cells transfected with OATP2B1 gene (Fujita et al., 2014). Although the reason for the discrepant results is not clear, one possible explanation is the differences among the experimental methods used. In our previous study in Xenopus oocytes, we used pcDNA3 plasmid vector to synthesize cRNA of OATP2B1 (Nozawa et al., 2005), whereas pGEMHE was used in the present study. It is known that pGEMHE plasmid vector is useful to achieve abundant expression of the target protein in Xenopus oocytes since the plasmid is able to stabilize the injected cRNA in Xenopus oocytes; consequently, protein synthesis is enhanced (Liman et al., 1992; Debont et al., 1996). The other study used OATP2B1-gene transfected human embryonic kidney cell line 293 (Fujita et al., 2014), a different host cell line from the present study. These differences may account for the fact that a positive result was obtained only in the present study.
Based on the finding that SN-38 is a substrate of intestinally expressed OATP2B1, we hypothesized that OATP2B1-mediated accumulation of SN-38 by enterocytes contributes to the gastrointestinal toxicity of CPT-11. Kampo medicine (containing baicalin) has been used for prevention of SN-38–induced late-onset diarrhea (Mori et al., 2003; Takasuna et al., 2006), whereas for pediatric patients, antibiotics such as cefixime have been used (Furman et al., 2006; McGregor et al., 2012). The effectiveness of these treatments has been explained in terms of inhibition of bacterial de-glucuronidation of biliary excreted SN-38G; however, in the present study, we found that baicalin and cefixime also reduced the OATP2B1-mediated uptake of SN-38 (Fig. 3, A and B). Clinically, Kampo medicine Hangeshashin-to (TJ-14), which contains baicalin, was administered orally at a dose of 7.5 g/day divided three times a day, namely, 2.5 g of TJ-14, which contains about 12.7–38.1 μmol baicalin. If the medicine is taken with a glass of water (200 ml), simple estimation of the intestinal luminal concentration of baicalin suggests a value in the range of 64–160 μM, which is higher than the IC50 for OATP2B1. In the case of cefixime, 8 mg/kg is orally administered once daily to prevent late-onset diarrhea in CPT-11 therapy of pediatric patients. Even if the patient’s body weight is only 10 kg, the calculated dose of cefixime is 80 mg (158 μmol). Since pediatric patients are considered to take the drug orally with a smaller volume of water than adults, the estimated intestinal luminal concentration of cefixime might be 1.5–3 mM, if 50–100 ml of water is taken, which again is higher than the IC50 for OATP2B1. Since these compounds act directly from the intestinal lumen on enterocytes, baicalin and cefixime both have the potential to inhibit OATP2B1-mediated uptake of SN-38 at clinically relevant doses. MIC of cefixime for Escherichia coli is less than 4 μM and the Km of baicalin for β-glucuronidase is about 38 μM (Neu et al., 1984; Ikegami et al., 1995). At the estimated intestinal luminal concentrations of cefixime and baicalin, therefore, they are expected to significantly affect intestinal microflora and bacterial β-glucuronidase activity, respectively, in the clinical setting. Accordingly, inhibition of OATP2B1 may represent an additional mechanism of reduced accumulation of SN-38 in intestinal tissues, reinforcing the effects of the previously proposed mechanisms. These results suggest that inhibition of OATP2B1 in enterocytes would contribute to effective treatment of SN-38–induced late-onset diarrhea.
We next evaluated the effect of fruit juices on OATP2B1-mediated transport of SN-38 since fruit juices interact with OATP2B1 and affect intestinal absorption of substrate drugs (Imanaga et al., 2011; Tamai, 2012; Tamai and Nakanishi, 2013). As shown in Fig. 4A, all the fruit juices tested significantly decreased OATP2B1-mediated uptake of SN-38. It should be noted that apple juice inhibits OATP2B1 in two different ways: reversible and long-lasting inhibitions (Shirasaka et al., 2013), though only reversible inhibition by apple juice was considered in the present in vitro experiments. These results suggest that apple juice would be useful for prophylactic therapy for gastrointestinal toxicity caused by CPT-11. Considering previously reported prolonged inhibition of OATP2B1 by apple juice, the effect of apple juice to alleviate gastrointestinal toxicity induced by CPT-11 may be stronger than expected, but further work is required.
Lastly, a protective effect of apple juice against gastrointestinal toxicity of SN-38 was observed in mice (Fig. 5). SN-38 significantly shortened the length of villi at 2 days after termination of 5-day continuous administration, which is in accordance with previous observations after administration of CPT-11 (Chen et al., 2013). Interestingly, apple juice was able to protect intestinal tissues from SN-38–induced gastrointestinal toxicity, presumably owing to decreased intestinal uptake of SN-38. As discussed, the acidic pH in the gastrointestinal tract is favorable for both diffusional and OATP2B1-mediated uptake of SN-38. These observations are consistent with the reduced gastrointestinal toxicity of CPT-11 resulting from intestinal alkalization with sodium bicarbonate (Ikegami et al., 2002), which decreases the accumulation of SN-38 in the tissues. In other words, the reported reduction of gastrointestinal toxicity by bicarbonate-dependent alkalization of the intestinal lumen can also be explained in terms of reduced transport via both diffusion and OATP2B1. Another issue that should be considered is the possible species difference between mice and humans. Intestinal expression of functional murine Oatp2b1 is not fully understood, but available evidence suggests that Oatp2b1 is expressed in mouse enterocytes and contributes to intestinal absorption of xenobiotics and drugs (Cheng et al., 2005; Iusuf et al., 2012). Accordingly, we consider that Oapt2b1 is functional in mice similarly to OATP2B1 in humans and that consequently the present observations can be extrapolated to humans.
In conclusion, this study provides evidence that OATP2B1 is involved in the uptake of SN-38 by intestinal tissues, a process that triggers gastrointestinal toxicity of CPT-11. Many enzymes and transporters, including carboxylesterase, uridinediphosphate glucuronyltransferase, cytochrome P450 3A4, breast cancer resistance protein, P-glycoprotein, multidrug resistance-associated protein 2, OATP1B1, and OATP1B3, have been shown to influence the pharmacologic and toxicologic actions of CPT-11. Our present findings implicate OATP2B1 as a new transporter molecule contributing to the intestinal disposition of SN-38. Therefore, this work uncovers a new aspect of the complex mechanisms of GI-toxicity of CPT-11 and opens up the possibility that reducing gastrointestinal accumulation of SN-38 by inhibiting OATP2B1-mediated intestinal transport is available as a novel strategy to protect gastrointestinal tissue. Since fruit juices are potent OATP2B1 inhibitors, they may contribute to the prophylaxis of gastrointestinal toxicity induced by CPT-11 and may be especially useful in pediatric patients, in whom the current use of antibiotics for this purpose may disturb the gut microflora. Further evaluation seems worthwhile to confirm efficacy in the clinical setting.
Acknowledgments
Authorship Contributions
Participated in research design: D. Fujita, Y. Saito, I. Tamai.
Conducted experiments: D. Fujita, T. Nakanishi.
Performed data analysis: D. Fujita, I. Tamai.
Wrote or contributed to the writing of the manuscript: Y. Saito, D. Fujita, T. Nakanishi, I. Tamai.
Footnotes
- Received August 6, 2015.
- Accepted October 30, 2015.
D.F. and Y.S. contributed equally to this work.
This work was supported by JSPS KAKENHI Grant Number 26670075 (I.T.).
Abbreviations
- CPT-11
- irinotecan hydrochloride
- HPLC
- high-performance liquid chromatography
- MBS
- modified Barth’s solution
- OATP
- organic anion transporting polypeptide
- SN-38
- 7-ethyl-10-hydroxycamptothecin
- SN-38G
- SN-38 glucuronide
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}