


Abstract
The biotransformation of TNP-470 [O-(chloroacetylcarbamoyl)fumagillol; AGM 1470], a potentin vitro inhibitor of angiogenesis, was investigated in primary cultured hepatocytes isolated from different species, including monkey, dog, and rat, as well as in microsomal fractions of various monkey tissues. Previous metabolic studies by our group using human hepatocytes in primary culture demonstrated that TNP-470 was primarily metabolized to M-IV through an ester cleavage, with subsequent conversion of M-IV to M-II by microsomal epoxide hydrolase. Additional studies using monkey liver microsomes demonstrated that M-II was then glucuronidated by uridine-5′-diphosphoglucuronyl transferase, leading to the formation of M-III. Three other, as yet unidentified, metabolites, labeled M-I, M-V, and M-VI, were also detected. Similarly to findings in human hepatocytes, the predominant extracellular metabolite was M-II in all species studied. Minor interspecies variability was observed in the total amount of drug biotransformed by hepatocytes, but some variability was detected in the metabolic pattern of TNP-470 in monkey hepatocytes, compared with rat or dog hepatocytes. In monkey hepatocytes, as previously observed in human cells, TNP-470 was metabolized to six derivatives, labeled M-I, M-II, M-III, M-IV, M-V, and M-VI, whereas the latter metabolite was not observed in dog or rat extracellular medium. Extrahepatic metabolism of TNP-470 was also studied using monkey intestine, kidney, and lung microsomes, which demonstrated that, under these experimental conditions, TNP-470 was extensively metabolized to four derivatives, i.e. M-I, M-II, M-III, and M-IV, with M-III being detected only in liver samples. These studies suggest that the metabolism of TNP-470 in monkeys appears to be most closely related to that observed in humans. Although the individual quantitative metabolic profiles were quite different in various animal species, only one metabolite, namely M-VI, was not detected in either dog or rat hepatocytes in vitro.
Angiogenesis (also called neovascularization) is the process by which new blood vessels are derived from preexisting vessels. This process has been shown to participate only in physiological states such as ovulation, menstruation, placenta development, and wound healing (1, 2). However, this aberrant growth of new blood vessels also plays a significant role in many pathological states, such as diabetic retinopathy, scleroderma, psoriasis, fractures, rheumatoid arthritis, and inflammation (3-7). Foremost, it is thought that angiogenesis is critical for the development and growth of common solid tumors (4, 5, 8, 9), also allowing them to metastasize (1). In fact, secretion of angiogenic factors by several tumors has been reported to activate angiogenesis (10, 11), and it has been hypothesized that control and inhibition of that neovascularization may provide a powerful and selective therapeutic approach for a large variety of metastatic cancers, leading to the search for effective angiogenesis inhibitors (4, 5).
Recently, it has been reported that TNP-470, a semisynthetic analogue of fumagillin, which is an antibiotic naturally secreted by the fungusAspergillus fumigatus fresenius (6), exhibits potent antitumor and antimetastatic activities (6, 12, 13). Furthermore, TNP-470 has been shown to be 50 times more potent than fumagillin in inhibiting capillary growth and to be less toxic than its parent derivative (6, 12, 14-17). The demonstration of in vitroand in vivo antitumor activity has led to initiation of phase I/IIa clinical trials in cancer patients (18, 19). In addition, TNP-470 has also been reported to inhibit Kaposi’s sarcoma-derived spindle cell proliferation at a concentration as low as 10 pg/ml (20), suggesting that this compound may represent a possible treatment for human immunodeficiency virus-associated Kaposi’s sarcoma. Previousin vitro and in vivo studies by our laboratory have demonstrated rapid and extensive metabolism of TNP-470 with formation of six metabolites, labeled M-I, M-II, M-III, M-IV, M-V, and M-VI (21-23).
This study sought to investigate TNP-470 biotransformation using primary cultured hepatocytes obtained from different species, including rats, dogs, and monkeys. These investigations, which detail the species differences in TNP-470 metabolism, should help in determining the adequacy of animal species used in the in vivopharmacological and toxicology preclinical studies of this novel class of antineoplastic drugs.
Materials and Methods
Chemicals.
[3H]TNP-470 [(3R,4S,5S,6R)-5-methoxy-4-[(2R,3R)-2-methyl-l-3-(3-methyl-2-butenyl)oxiranyl]-1-[6-3H]oxaspiro-[2.5]-oct-6-yl-(chloroacetyl)carbamate] (2.6 Ci/mmol or 6.5 mCi/mg) and authentic standards of unlabeled TNP-470, unlabeled metabolite M-IV (AGM-1883), and unlabeled metabolite M-II (T71712) were synthesized by Takeda Chemical Industries (Osaka, Japan) and were >96% pure, as ascertained by the HPLC method described below. The structure of the compounds was confirmed by proton NMR, 13C NMR, and LC-MS. All chemicals used in these studies were of analytical grade.
Isolation of Rat, Dog, and Monkey Hepatocytes.
Studies were performed using rat hepatocytes in culture, which were freshly isolated from male Sprague-Dawley rats (200–250 g; Harlan Laboratories, Indianapolis, IN). Dog and monkey hepatocytes were prepared from whole livers of Beagle dogs and cynomolgus monkeys. Male Beagle dogs (12–15 kg) and male Cynomolgus monkeys (3–4 kg) were both supplied from Lebeau Ets. (Gambais, France). Animals were killed by anesthetic overdose and hepatocytes were obtained by a two-step collagenase perfusion, as described previously (24). Freshly isolated rat hepatocyte viability, determined by the erythrosin B exclusion test, was approximately 85%.
Cryopreservation and Thawing of Monkey and Dog Hepatocytes.
Monkey hepatocytes were cryopreserved in L15 medium containing 25 g/liter bovine serum albumin, 20 g/liter polyvinylpyrrolidone, 10% dimethylsulfoxide, and 20% fetal calf serum, in 1.8-ml sterile polypropylene vials, as described previously (22). Dog hepatocytes were cryopreserved in the same medium but containing 40% Belzer liquid instead of bovine serum albumin and polyvinylpyrrolidone. Cell freezing was performed with a Nicool ST20 apparatus (Air Liquide, Grenoble, France), which had been previously programmed to optimize temperature drop, and hepatocytes were subsequently stored in liquid nitrogen until used.
Cells were thawed by immersing vials for 2 min in a water bath at 37°C. The suspension was diluted in L15 medium containing 10% fetal calf serum at 37°C and was then centrifuged at 40g to remove cryopreservation medium. Viable cells were separated by a Percoll density gradient, as described previously (25). The cells were then centrifuged and washed in L15 medium containing 10% fetal calf serum, and cell counting and viability determination were performed as described above. After Percoll gradient separation, viability after thawing was 94%.
Seeding and Culture of Rat, Dog, and Monkey Hepatocytes.
Thawed and freshly isolated hepatocytes were resuspended in L15 medium, seeded in six-well plates, and then incubated in a humidified 5% CO2 atmosphere at 37°C, as previously described by our laboratory in previous studies using human hepatocytes (21).
Hepatocyte Incubation Conditions.
Hepatocytes isolated from rats, dogs, and monkeys in L15medium were incubated with 5 μM [3H]TNP-470 (specific activity, 70 dpm/pmol) for specified times between 5 min and 4 hr, at 37°C. Times of incubation in the text represent the length of time between the introduction of radiolabeled TNP-470 into the cultured cells and the addition of acetonitrile to stop the reaction. After addition of an equal volume of acetonitrile, extracellular medium aliquots (100 μl) were analyzed by HPLC, without further processing, as described below.
Preparation of Microsomes.
Monkey samples obtained from different organs such as liver, intestine, kidney, and lungs were initially perfused in situ with Eurocollins or Belzer liquid at 4°C and stored at −80°C until used. Microsomal fractions were prepared by following the protocol described by our group using human microsomes obtained from different organs such as liver, intestine, stomach, and kidney (21). Aliquots of the microsomal suspension (0.25 ml) were stored at −80°C until used. The entire procedure was performed at 0–4°C, and no loss of enzyme activity was observed for up to 1 year after isolation. Protein content was quantitated by the method of Bradford (26), using bovine serum albumin as standard.
All assay mixtures contained 0.1 mg/ml microsomal protein, 100 mM potassium phosphate monobasic, 100 mM NADPH, and 50 μM [3H]TNP-470 (specific activity, 4.44 dpm/pmol), in a final volume of 1 ml. Reaction was initiated by adding microsomal protein, and samples were incubated at 37°C for 15 min and terminated by adding an equal volume of acetonitrile. Proteins were then removed by centrifugation at 14,000g for 5 min in a 5314 Eppendorf microcentrifuge (Brinkman Instruments, Inc., Westbury, NY), and aliquots (100 μl) were analyzed by HPLC. Control incubation conditions at 0 and 15 min were in the absence of either NADPH or microsomal protein. All experiments were carried out in duplicate under conditions leading to linear reaction rates with time, protein content, and cofactor concentration.
Analysis of [3H]TNP-470 and Its Metabolites by HPLC.
Samples were analyzed by HPLC using a Hewlett-Packard model 1050 liquid chromatograph equipped with a manual injector and a fixed-wavelength spectrophotometer (Hewlett-Packard, Palo Alto, CA). Reverse-phase chromatography was performed with a Hypersil ODS 5-μm column (Jones Chromatography, Littleton, CO), and elution was performed at 1 ml/min with a 50 mM KH2PO4 buffer (pH 3) and a 40-min linear gradient of acetonitrile from 0 to 25%, starting at the time of injection, followed by an increase to 50% acetonitrile between 40 and 50 min. Column temperature was maintained at 25°C, and absorbance was monitored at 210 nm. Eluent from the column was directed by way of a low-dead volume connection line into A 500 TR Radiomatic FLO-ONE radiochromatography analyzer (Packard Instrument Co., Meriden, CT). Under the above conditions, retention times for TNP-470 and its metabolites M-IV and M-II were 58, 52, and 44 min, respectively. Four unidentified chromatographic peaks, labeled M-I, M-III, M-V, and M-VI, eluted at 48, 38, 28, and 26 min, respectively. Intraday and interday coefficients of variation were <5% for all analysis. The standard curve for TNP-470 was linear with r2 > 0.99. However, the unavailability of radiolabeled metabolites did not permit the establishment of standard curves for these derivatives.
Isolation and Identification of M-III.
Metabolite M-III was isolated from extracellular medium by HPLC as described above, and then fractions were pooled and lyophilized. The dry residue was then dissolved in 1 ml of water, passed through a 0.45-μm Acro LC13 filter (Gelman Sciences, Ann Arbor, MI), and applied to a C18 Sep-Pak cartridge (Waters, Milford, MA) that had been preconditioned with 1 ml of methanol and 1 ml of water. After sample loading, the cartridge was washed with 3 ml of water and increasing percentages of acetonitrile from 5 to 50%. Metabolite M-III was eluted with a 25% acetonitrile mixture, and a 50-μl sample was chromatographed to ensure purity. The remaining eluent was lyophilized and analyzed by LC-MS.
[3H]TNP-470 (50 μM) was incubated in the presence of 25 mM UDPGA1 in a mixture containing 0.04% (v/v) Triton X-100, 50 mM MgCl2, and 3 mg/ml microsomal protein, in a final volume of 1 ml. Reaction was initiated by adding UDPGA, and samples were incubated for 4 hr at 37°C. Incubations were terminated by addition of 200 μl of acetonitrile, proteins were removed by centrifugation at 15,000g for 4 min in an Eppendorf model 5314 microcentrifuge, and 100-μl aliquots were analyzed by HPLC. Then the purified metabolite M-III was incubated for 2 hr at 37°C in the presence of 5000 units of β-glucuronidase in a 0.2 M sodium acetate (pH 4.5) buffer. After incubation, a 100-μl aliquot was chromatographed by HPLC.
LC-MS Analysis.
Analyses were performed on an API-III triple-quadruple mass spectrophotometer (P.E.-Sciex, Kornhill, Ontario, Canada) equipped with two Macintosh Quadra 950 computers for data analysis. The purified metabolite M-III was injected into a reverse-phase HPLC system with a 10-cm × 2.1-mm Aquapore C8 column (Applied Biosystems), at a flow rate of 0.2 ml/min, with a linear gradient of acetonitrile from 0 to 50%. The column eluate was split 1:1, with 100 μl going into the electrospray interface.
Results and Discussion
Analysis of the Time Course of Disappearance of Extracellular TNP-470 and Appearance of Extracellular Metabolites.
The present study provides a detailed analysis of the metabolism of a novel angiogenesis inhibitor, TNP-470, using cultured hepatocytes isolated from different species, including monkeys, dogs, and rats. These investigations of TNP-470 biotransformation and disposition in intact cells were complemented with studies using subcellular fractions of monkey tissues. This study was undertaken on the basis of an hypothesized metabolic pathway from previous in vitrostudies using human cultured hepatocytes and microsomes (21), which included the conversion of TNP-470 to M-IV through an “ester” cleavage, with further conversion of M-IV to M-II by microsomal epoxide hydrolase. These observations were consistent with the proposed structures of both metabolites, as determined by fast-atom bombardment MS and proton NMR (fig. 1). In addition, the identity of the chromatographic peak corresponding to metabolite M-III was determined by LC-MS as described in Materials and Methods. The LC-MS spectrum of M-III biologically formed in monkey extracellular medium demonstrated a single negative molecular ion (M−H)−, at m/z 518 (fig.2).

Chemical structures of TNP-470 and its metabolites M-IV, M-II, and M-III.
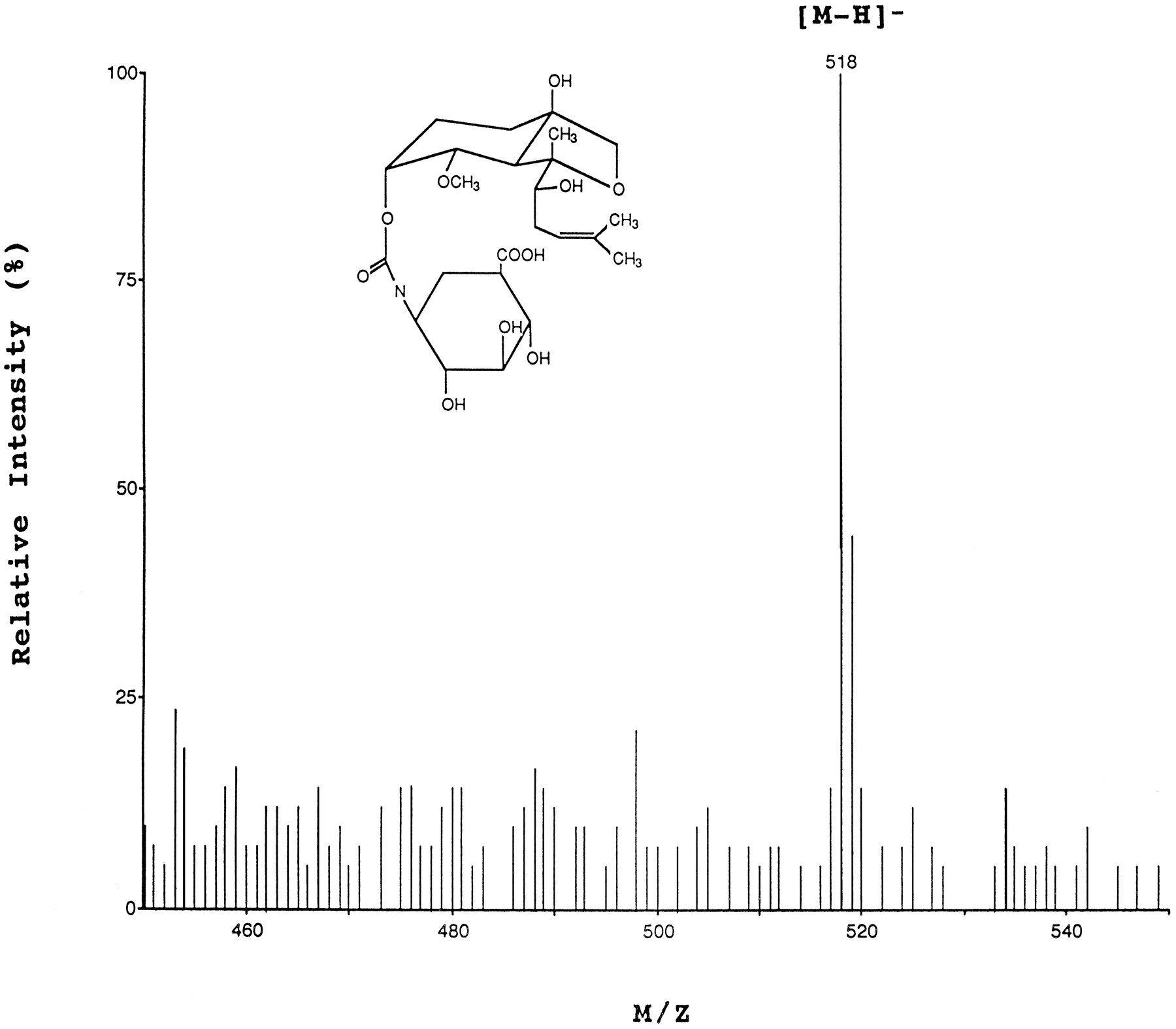
LC-MS spectra, obtained with electrospray interface, of the purified chromatographic peak labeled M-III, isolated from primary cultures of monkey hepatocytes.
Furthermore, enzymatic reactions were performed to identify the structure of this metabolite M-III. [3H]TNP-470 was incubated in the presence of UDPGA and microsomal proteins. The HPLC radiochromatograms of [3H]TNP-470 without or with 25 mM UDPGA are shown in fig. 3, A andB. Similar results were obtained when M-II was incubated with the same concentrations of UDPGA and microsomal proteins. These data were consistent with metabolite M-III being a glucuronide derivative of M-II. Additional information and final identification of this glucuronide derivative were obtained after incubating M-III with β-glucuronidase. Results showed that M-III was converted into M-II by β-glucuronidase, as shown in fig. 3C.

HPLC radiochromatograms of M-II and M-III after a 4-hr incubation of monkey liver microsomes in the absence (A) or presence of 25 mM UDPGA (B) or 5000 units of β-glucuronidase (C).
Data are representative of three different experiments, with duplicate incubations.
Hepatocytes isolated from different species have been routinely and increasingly used over the past few years for pharmacological and toxicological studies (21, 27, 28). Hepatocytes express most of the functional and metabolic activities of intact liver and provide an opportunity to study both the pathways and extents of metabolism of novel drugs at a very early stage in the drug development process. Thein vitro comparisons performed in this study provide information on both rates of reaction and metabolic profiles among the three studied species and confirm our previous studies using cultured human hepatocytes and microsomal fractions, as well as in vivo studies using rhesus monkeys, which demonstrated that TNP-470 was rapidly metabolized to six different metabolites (21, 22).
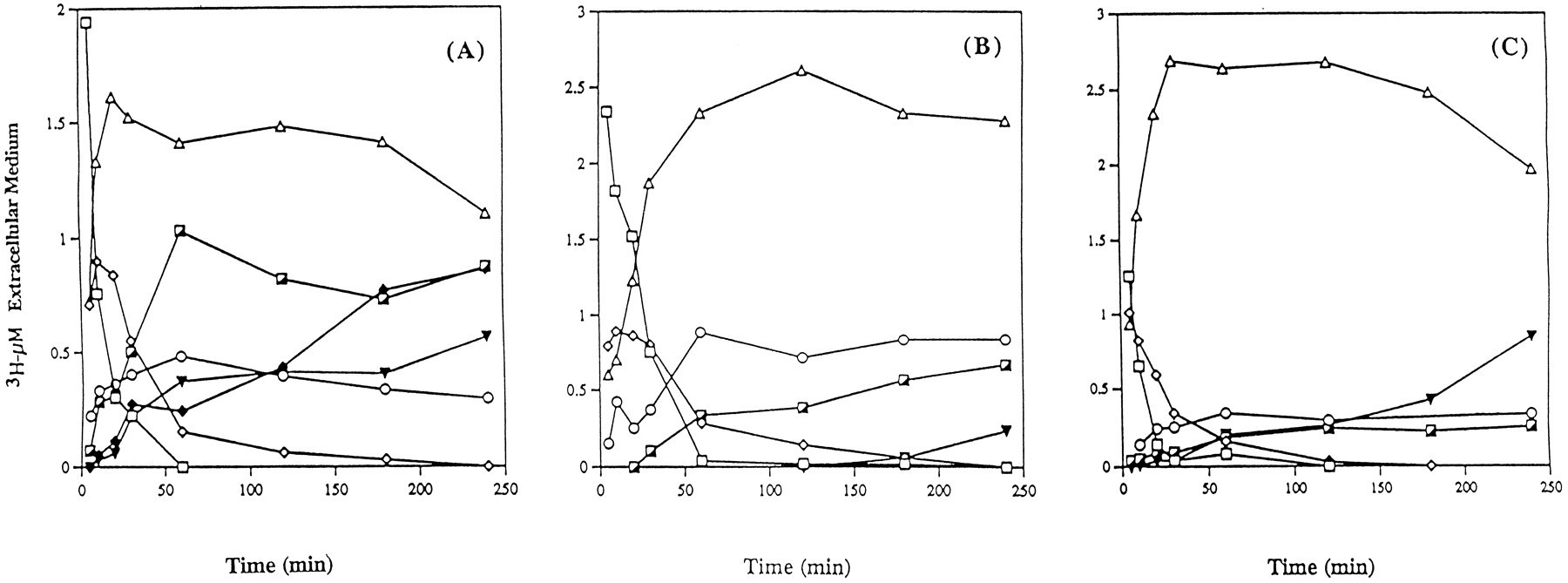
Although no major difference was observed regarding the extracellular profiles of unchanged TNP-470 and its metabolite M-IV among species, a few quantitative and qualitative differences were detected in the other metabolic steps. Fig. 4 illustrates these extracellular kinetics in monkey (fig. 4A), dog (fig. 4B), and rat (fig. 4C). In all species, unchanged TNP-470 disappeared rapidly from the extracellular compartment, reaching one-fifth of its initial level over 10–30 min, and was then undetectable by 1–3 hr. Similarly, its metabolite M-IV, after reaching a maximum level of 1.01, 0.90, and 0.89 μM after a 5-min incubation with rat hepatocytes and a 10-min incubation with dog and monkey hepatocytes, respectively, rapidly decreased to undetectable levels in the same time period. As extracellular TNP-470 and M-IV levels decreased, subsequent metabolites that were synthesized within cells appeared in the extracellular compartment. M-II was by far the predominant metabolite and attained a maximum level of 2.69 μM over 30 min in rat hepatocytes, 2.33 μM over 2 hr in dog hepatocytes, and 1.61 μM over 20 min in monkey hepatocytes. Interestingly, marked quantitative differences were observed among the various species. In all species, three other metabolites were observed. M-I was rapidly detected within the early incubation times and attained maximum levels of 0.88, 0.48, and 0.34 μM over 60 min in dog, monkey, and rat extracellular medium. M-III appeared slowly and reached a maximum of only 0.24 μM over 2 hr in rat hepatocytes, 0.66 μM over 4 hr in dog hepatocytes, and 1.03 μM over 1 hr for monkey hepatocytes. M-V appearance was significantly variable, depending on the species. In rat and monkey hepatocytes, M-V appeared at 10 min and increased over the 4-hr exposure of TNP-470 to the cells, reaching levels of 0.42 and 0.56 μM, respectively. In dog hepatocytes, M-V appeared with a lag in its formation for 3 hr and then reached a level of 0.23 μM at 4 hr. Furthermore, the only qualitative difference among the three species studied was the absence of M-VI in dog and rat hepatocytes, whereas in monkey hepatocytes that metabolite reached a maximum level of 0.86 μM within 4 hr.

Analysis of unchanged TNP-470 (□) and its metabolites M-IV (⋄), M-I (○), M-II (▵), M-III (┌), M-V (▾), and M-VI (♦) in the extracellular medium after exposure of monkey (A), dog (B), and rat (C) hepatocytes to 5 μM [3H]TNP-470.
At the indicated times, portions of the cell suspension were separated by centrifugation, and total radioactivity was assayed by HPLC. All data presented represent the mean of three experiments.
Metabolism of [3H]TNP-470 in Monkey Tissue Microsomes.
Because data obtained in monkeys were in complete agreement with those obtained in human hepatocytes (21), complementary experiments using monkey microsomes isolated from different organs, such as liver, intestine, kidney, and lungs, were performed. This study confirmed that the liver was the major site of TNP-470 metabolism and that the intestine represents another important site for TNP-470 biotransformation, as previously observed in human microsomes (21). Table 1 summarizes major interorgan differences in the metabolism of [3H]TNP-470 in several tissue microsomal fractions obtained from monkeys. As observed previously in human microsomal fractions, M-IV was the predominant metabolite; the highest levels were detected in liver and intestine microsomes, with values being approximately 3-fold higher than those in kidney and lung microsomes. M-I appeared only in liver and kidney microsomes, reaching concentrations of 35.6 and 25.6 nmol/mg of protein, respectively. The maximal concentration of M-II detected was in kidney microsomes, with a value of 126.8 nmol/mg of protein, whereas M-II levels remained very low in liver and intestine samples (9.4 and 5.9 nmol/mg of protein, respectively). Of note, M-III was detectable in liver microsomal fractions, with a level of 28.0 nmol/mg of protein. Under these conditions, the formation of M-V and M-VI was not detected in liver microsomes, probably due to the short incubation period and low protein concentration. In conclusion, the present studies demonstrate that the complex metabolism of TNP-470 in human hepatocytes (21) is paralleled by that observed in monkey hepatocytes, demonstrating that monkey is the closest species to humans to evaluate TNP-470 metabolism and biodisposition. The minor qualitative and quantitative metabolic differences observed with the other species, i.e. dogs and rats, also suggest the adequacy of the in vivopharmacological and toxicological preclinical studies using these animal species.
Amounts of TNP-470 metabolite formed in different monkey tissue microsomes after incubation of 50 μM [3H]TNP-470 (specific activity, 4.4 dpm/pmol) and 10 MM NADPH at 37°C, for 15 min, in the presence of 0.1 mg/ml microsomal protein.
Acknowledgments
We thank Marion Kirk for performing the MS analyses.
Footnotes
-
Send reprint requests to: Jean-Pierre Sommadossi, Department of Pharmacology and Toxicology, University of Alabama at Birmingham, Volker Hall 600, Birmingham, AL 35294-0019.
-
This work was supported in part by United States Public Health Service Grant AI32775 and by an unrestricted grant from TAP Pharmaceuticals, Inc. J.-P.S. is the recipient of a Faculty Research Award from the American Cancer Society. The mass spectrometer was purchased with funds from a National Institutes of Health Instrumentation Grant (S10RR06487) and from this institution. The University of Alabama at Birmingham Comprehensive Cancer Center Mass Spectrometry Shared Facility is supported in part by a National Cancer Institute Core Research Support Grant (P30-CA13148) to the University of Alabama at Birmingham Comprehensive Cancer Center.
- Abbreviation used is::
- UDPGA
- UDP-glucuronic acid
- Received June 28, 1996.
- Accepted September 17, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}