


Abstract
Eletriptan (Relpax) is a novel 5-hydroxytryptamine (serotonin)1D/1B agonist currently in development for the acute treatment of migraine. The aim of this work was to evaluate the relative induction potency of eletriptan in vitro compared with well characterized cytochrome P-450 (CYP) inducers with primary cultures of human hepatocytes and to relate this to the situation in vivo. Eletriptan was a weak inducer of CYP3A4 protein and cyclosporin A oxidation in four of the six cultures used, whereas rifampicin was a potent inducer in all cultures. Induction was concentration dependent and not detectable at eletriptan concentrations of 5 μM and lower. The amplitude of the increase in CYP3A4 protein and activity by 25 μM eletriptan was significantly lower, with a mean of 19 (P = .0015) and 26% (P = .0002), respectively, of that observed in response to 25 μM rifampicin. CYP2A6, a protein with minor pharmacological implication, also was induced by eletriptan and rifampicin in two cultures but was not detected in the others. The levels of other CYP proteins, including CYP1A2, CYP2C9, CYP2C19, CYP2D6, and CYP2E1, were not affected by eletriptan. Because the maximum blood concentration of eletriptan in humans after a therapeutic dose (maximum 80 mg) is 0.5 μM, the in vitro model would predict no clinically significant induction of CYP3A4 protein in vivo. This has been confirmed subsequently in a clinical study, with 6β-hydroxycortisol/cortisol ratios as marker of CYP3A4 activity. Eletriptan is therefore not an inducer of CYP3A4 at clinical doses.
Eletriptan (Relpax) {(R)-3-(1-methyl-2-pyrrolidinylmethyl)-5-[2-(phenylsulphonyl)ethyl]-1H-indole}, is a new selective agonist at the 5-hydroxytryptamine (serotonin)1D/1B receptor (Gupta et al., 1996). The compound has been shown to be clinically effective in the acute treatment of migraine and is well tolerated (Farkkila, 1996; Jackson, 1996). Migraine affects ∼18% of all women compared with 6% of men (Stewart et al., 1992; Rasmussen and Olesen, 1994; Hopkins, 1996) and prevalence is highest between the ages of 25 and 55 years (Hopkins, 1996). This results in a migraine prevalence of 25% in premenopausal women, many of whom will use the oral contraceptive. Therefore, it is important to predict whether eletriptan is an inducer of the metabolism of oral contraceptives because such an effect, if large enough, could result in a clinically significant reduction in the effectiveness of the oral contraceptive.
The primary route of metabolism for the major synthetic estrogen oral contraceptive (ethinylestradiol) is cytochrome P-450 (CYP)1dependent, and CYP2C9 and CYP3A4 have been shown to be involved (Guengerich, 1988; Ball et al., 1990). CYP3A4 is the main form of CYP expressed in the adult human liver (Maurel, 1996a). This enzyme has been shown to metabolize >60% of drugs under current use and, in addition, is inducible by a number of structurally unrelated compounds (Maurel, 1996a). It is well documented that inducers of CYP3A4, including many antiepileptic drugs and rifampicin (RIF), interact with oral contraceptive (Guengerich, 1988). In a review of oral contraceptive drug interactions, the coadministration of these compounds has been associated with contraceptive failure in several cases (Shenfield, 1986). Therefore, the relative induction potency of eletriptan compared with a well documented inducer of CYP activity, such as RIF, would give an indication of possible clinical interactions between eletriptan and oral contraceptive.
Primary cultures of human hepatocytes have been shown to represent an invaluable in vitro model for predicting the metabolic pathway and adverse effects of drugs in humans (Maurel, 1996b). Previous reports from our and other laboratories (Diaz et al., 1990; Morel et al., 1990;Pichard et al., 1990, 1992; Schuetz et al., 1993; Curi-Pedrosa et al., 1994; Li et al., 1995; Chang et al., 1997) have focused on the use of these cultures for the screening of drugs that could be enzyme inducers. CYP1As, CYP2A6, CYP2Cs, and CYP3A4 were found to be inducible in these cultures and the agreement with clinical data in humans was fair (Morel et al., 1990; Pichard et al., 1990, 1992;McDonnell et al., 1992; Rost et al., 1992; Maurel, 1996b). The aim of this work was to determine whether eletriptan is an inducer of CYP3A4 protein in human hepatocytes in primary culture and to extrapolate the data obtained to the situation in humans in vivo. This combined approach provides a guide as to the clinical consequences of coadministration of eletriptan and CYP3A4 substrates.
Materials and Methods
Human Liver Samples.
Human liver samples were obtained from eight patients, seven undergoing liver lobectomy for colon tumor metastasis (FT103, a 56-year-old female; FT110, a 45-year-old female; FT112, a 70-year-old female; FT113, a 61-year-old male; FT127, a 52-year-old female; FT142, a 47-year-old male; and FT151, a 61-year-old female) and one patient (FT105, a 57-year-old male) who became an organ donor after a massive digestive hemorrhage. In this last case, the liver was refused for transplantation because clinical investigations revealed the presence of pancreatic neoplasia. The use of these samples for scientific research was authorized by the French National Ethics Committee.
Primary Cultures of Human Hepatocytes.
Primary cultures of human hepatocytes were prepared as described in previous articles (Diaz et al., 1990; Pichard et al., 1990). The viability of cells before plating was determined with the trypan blue exclusion test and comprised between 72 and 83%. Eight million cells in 8 ml of culture medium were placed into 100-mm plastic dishes precoated with collagen. The culture medium consisted of a 1:1 mixture of Ham F12 and Williams' E, supplemented as described (Isom and Georgoff, 1984). The culture medium was supplemented with 5% calf serum during the first 4 h after plating to favor the attachment of cells. Then, the medium was changed and subsequently renewed every 24 h in the absence of serum. Cultures were maintained at 37°C in a humid atmosphere of air and 5% carbon dioxide. For the treatment of cells, prototypical inducers, including RIF, 2,3,7,8-tetrachlorodibenzo(p)dioxin (TCDD), and eletriptan were diluted in dimethyl sulfoxide (DMSO) and added to the culture medium at the following concentrations: 25 μM (RIF), 1 nM (TCDD), and 1 to 50 μM (eletriptan), depending on the experiments. In all cases, the concentration of DMSO was 0.1%, and control cultures received only DMSO at the same concentration. The treatments lasted for 96 h and were renewed every 24 h as the culture medium was changed.
Hepatocyte Microsome Preparation.
Microsomes were prepared from hepatocyte cultures by differential centrifugation and stored as described previously (Diaz et al., 1990;Pichard et al., 1990). Protein concentration was determined by the bicinchoninic acid method, according to the protocol provided by the manufacturer (Pierce Chemical Co., Rockford, IL). BSA (Pierce Chemical Co.) was used as the standard.
Measurement of Monooxygenase Activities.
Five hundred micrograms of microsomes was resuspended in 500 μl of 0.1 M potassium phosphate buffer, pH 7.4, in the presence of adequate concentrations of the various substrates (ethoxyresorufin, coumarin, and cyclosporine). After a 3-min incubation at 37°C, the reaction was initiated by the addition of 1 mM NADPH and quenched 5 to 30 min later (under conditions of linear kinetics). Oxidation of the substrates was monitored according to previously published procedures (Diaz et al., 1990; Pichard et al., 1990; Dalet-Beluche et al., 1992). Monooxygenase activities were expressed as nanomoles of metabolite produced per minute, per milligram of protein.
Immunoquantitation of CYP Proteins.
CYP proteins, including forms 1A2, 2A6, 2C9, 2C19, 2D6, 2E1, and 3A4 were quantitated by immunoblot with specific polyclonal antibodies, as described previously (Diaz et al., 1990; Pichard et al., 1990;Dalet-Beluche et al., 1992). Routinely, 20 to 100 μg of liver microsomes prepared from the culture after treatments were submitted to electrophoresis on SDS 10% polyacrylamide gel before transferring to nitrocellulose. Blots were developed with specific anti-CYP antibodies and the enhanced chemiluminescence (ECL) method (Amersham Corp., Amersham, UK). The relative amount of CYP was estimated from densitometric analysis of the blot with a scanner (Shimadzu, Tokyo, Japan).
RNase Protection Assay.
A 1014-base pair CYP3A4 cDNA fragment (93–1107) was amplified by polymerase chain reaction with two oligonucleotides, 5′ sense: TGGAAACCTGGCTTCTCCTG and 3′ antisense: GGTGGGTGGTGCCTTATTGGG (Beaune et al., 1986), and subcloned in plasmid pT3T7a18. The plasmid was linearized with EcoRI, which cuts into the insert at position 905, and the CYP3A4 antisense RNA probe was synthesized with the T3 RNA polymerase. The native probe was 250 nucleotides in length and the protected probe was 202 nucleotides in length (905–1107). Total RNA (100–150 μg) extracted from treated cells was hybridized with radiolabeled CYP3A4 antisense RNA probe (300,000 cpm) and analyzed by the RNase protection assay as described (Melton et al., 1984;Muntanet-Relat et al., 1995). The protected CYP3A4 RNA probe was revealed by autoradiography. The quality control of RNA samples was assessed by Northern blot, with a human glyceraldehyde 6-phosphate dehydrogenase (GAPDH) cDNA probe.
In Vivo Study.
Ethical approval was obtained for the study and all subjects gave written informed consent. Sixteen healthy male Japanese subjects ranging in age from 20 to 45 years were included into the study as two groups of eight. The study was a randomized, double blind, placebo-controlled, multiple-dose design with eletriptan doses of 40 mg t.i.d. (i.e., 120 mg/day), or 80 mg b.i.d. (i.e., 160 mg/day) given over 7 days. Urine was collected for 24 h before dosing on day 1, and then for 24 h postdose, on days 1 and 7. Urine was assayed for cortisol and 6β-hydroxycortisol by HPLC-mass spectrometry (Ged et al., 1989) or HPLC-UV. Urinary 6β-hydroxycortisol was chromatographed with an HPLC column-switching (heart-cutting) technique (Abel et al., 1992). Initial separation from urine constituents was achieved with a Kromasil 5C1 column (10 cm × 4.6 mm) with a mobile phase of ammonium phosphate (0.02 M, pH 6.0)/methanol (83:17 v/v) at a flow rate of 1.5 ml/min. The fraction of eluent containing 6β-hydroxycortisol was trapped in a sample loop and switched to a less polar Kromasil 5C18 column (25 cm × 4.6 mm), and eluted with a mobile phase of water/acetonitrile (85:15 v/v) at a flow rate of 1 ml/min. Final detection was by UV absorbance at 250 nm.
Statistical Analysis.
Because of the limited number of cultures analyzed, and because the levels of CYP3A4 protein and mRNA as well as the rate of cyclosporin A oxidation are widely variable from one culture to another, the data were normalized with respect to the maximum value attained in each culture (taken as 100) (i.e., in most of cases, in response to RIF). Under these conditions, the data were normally distributed. The effect of eletriptan was then compared with that of RIF with the pairedt test. Data were expressed as relative levels of CYP3A4 protein and mRNA or relative rate of cyclosporin A oxidation, as means ± S.D. Statistical analyses were performed with the MacIntosh Stat View program (Abacus Concepts, Berkeley, CA).
A general linear model was used for log-transformed 6β-hydroxycortisol/cortisol, for each group, comparing days 1, 7, and predose. These analyses allowed for variation due to subject and day. Differences between means, S.E. values associated with these differences, and the 95% CI for the differences were presented for 6β-hydroxycortisol/cortisol. The ratio between antilogged treatment means and the corresponding antilogged confidence intervals also was presented.
Results
Effect of Eletriptan on Expression of CYP3A4 Protein and on Rate of Cyclosporine A Oxidation in Human Hepatocytes.
Human hepatocytes were maintained in culture for 96 h in the absence of treatment or in the presence of eletriptan or of prototypical inducers of CYP3A or CYP1A, RIF, or TCDD, respectively. At the end of treatments, the level of CYP3A4 protein and the rate of cyclosporine A oxidation, a CYP3A4-specific activity (Pichard et al., 1990; Maurel, 1996a), were measured in microsomes by immunoblot and HPLC, respectively.
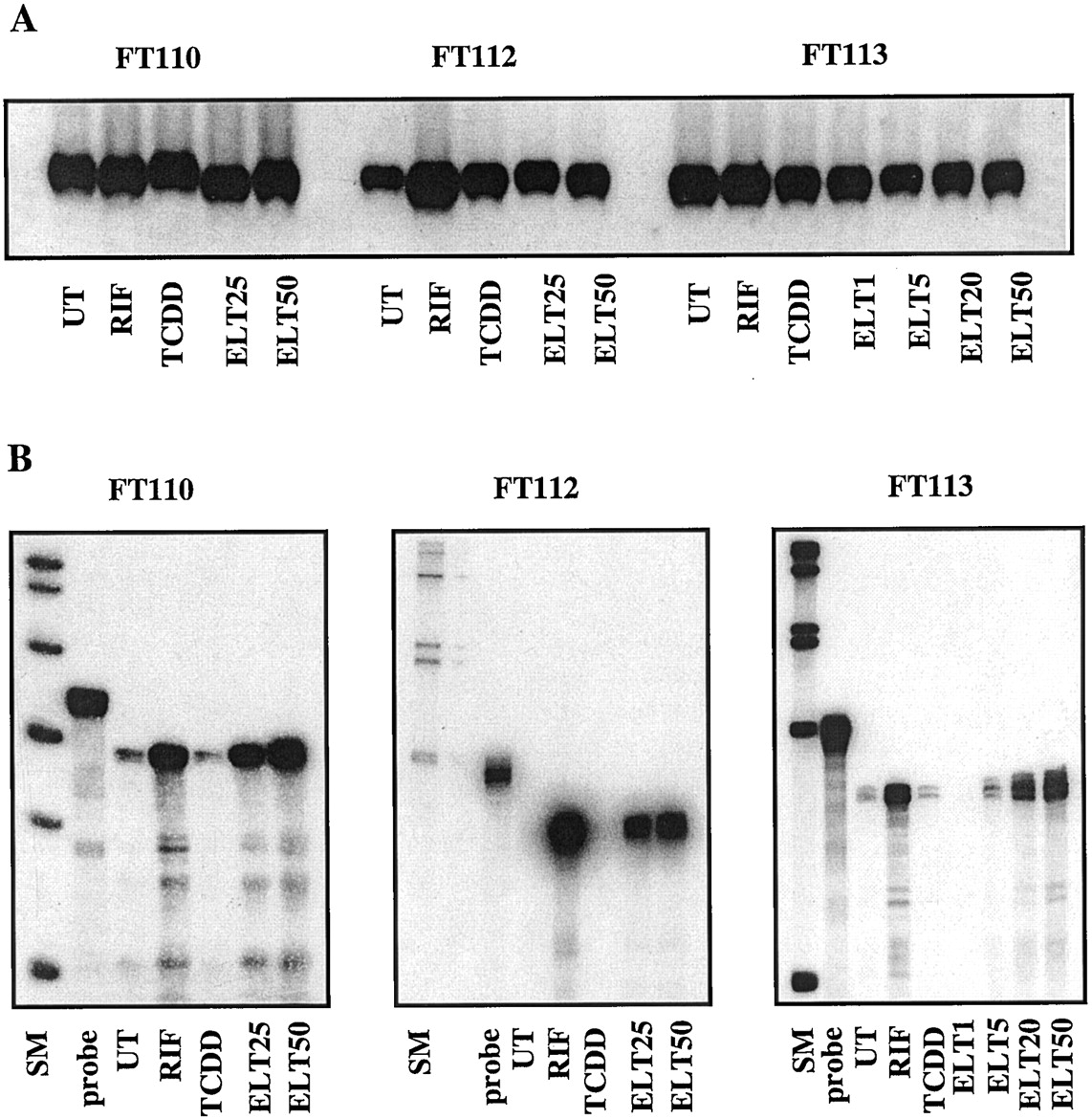
As reported in Fig. 1, CYP3A4 protein was clearly induced in all cultures by 25 μM RIF, although there was approximately an 8-fold variability of the accumulation of this CYP protein from one culture to another, as observed previously (Pichard et al., 1990, 1992; Curi-Pedrosa et al., 1994; Chang et al., 1997). TCDD was not an inducer, as expected. CYP3A4 protein was induced in response to eletriptan (25 or 50 μM) in four cultures, including FT103, FT105, FT112, and FT127, but not in cultures FT110 and FT113. These observations were clearly confirmed when the rate of cyclosporine A oxidation was measured in the same microsomes, as reported in Fig.2. Both the protein and rate of cyclosporine A oxidation exhibited a concentration-dependent increase in cultures FT105, FT112, and FT127. Closer examination of the concentration dependence (1–50 μM) with culture FT127 revealed that induction of CYP3A4 protein and cyclosporine A oxidation by eletriptan becomes detectable at concentrations >5 μM and increases between 25 and 50 μM. The reason why the rate of cyclosporine A oxidation in microsomes from culture FT127 is very low compared with the other cultures is unknown. The data obtained with the six different cultures tested in this work are summarized in Table1. The level of CYP3A4 protein and the rate of cyclosporine A oxidation were statistically significantly lower (P < .02 and P < .001, respectively) in cells treated with eletriptan at 25 and 50 μM than in cells treated with 25 μM RIF.

Quantification of CYP3A4 protein in microsomes prepared from primary hepatocyte cultures after various treatments.
Hepatocytes from patients FT103, FT105, FT110, FT112, FT113, and FT127 were cultured for 96 h in the absence (UT) or in the presence of 1 nM TCDD; 25 μM RIF; or 1, 5, 25, or 50 μM eletriptan (ELT). Microsomes were extracted and 100 μg was assayed by immunoblot with anti-CYP3A6-specific antibodies and revealed by the ECL method (Amersham Corp.). Authentic standard (STD) was a sample of microsomes (50 μg) from a human lymphoblastoid cell line transfected with a human CYP3A4 cDNA (Gentest, Woburn, MA).
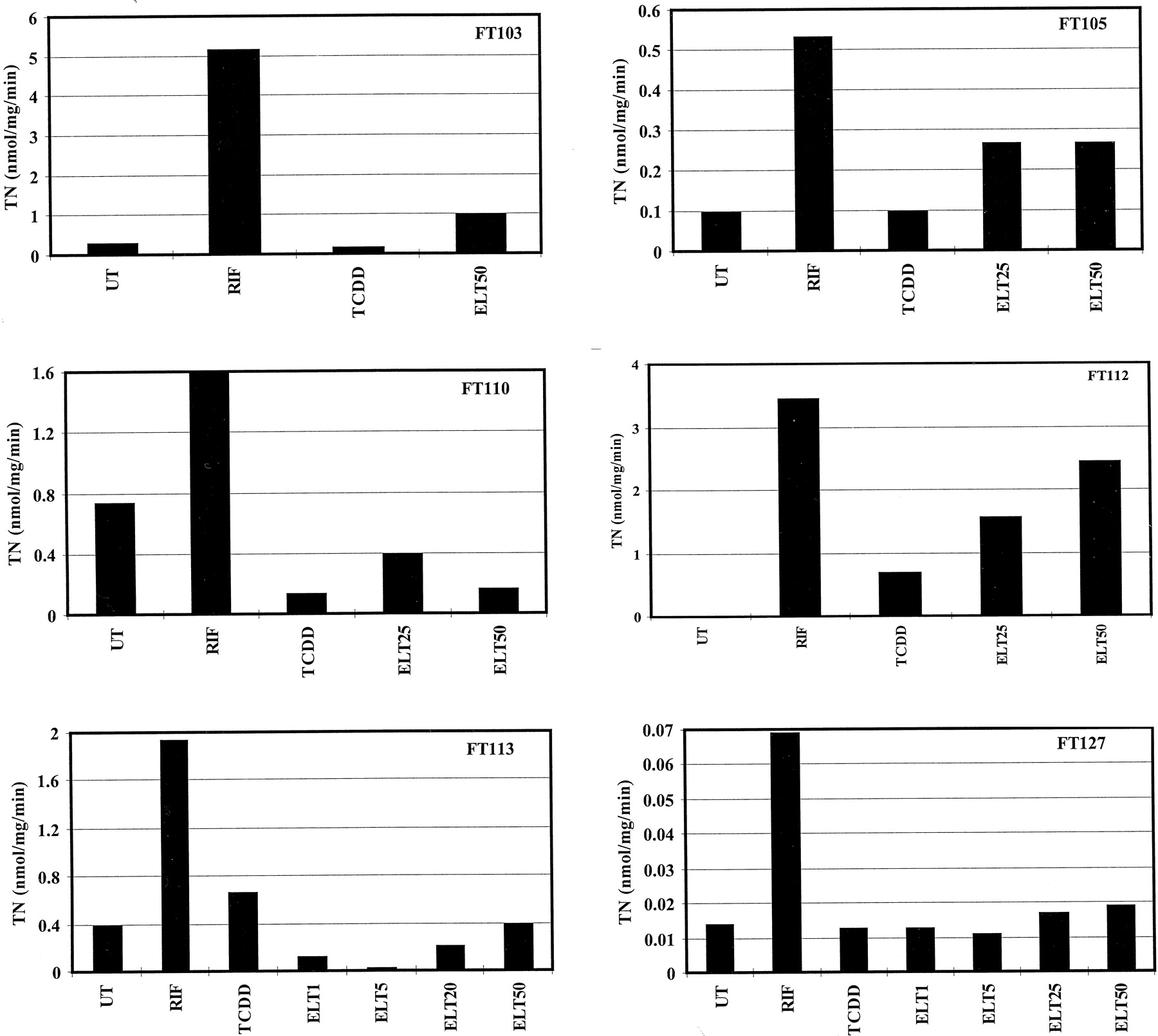
Rate of cyclosporine A oxidation in microsomes prepared from primary hepatocyte cultures after various treatments.
Hepatocytes from patients FT103, FT105, FT110, FT112, FT113, and FT127 were cultured for 96 h in the absence (UT) or in the presence of 1 nM TCDD; 25 μM RIF; or 1, 5, 25, or 50 μM eletriptan (ELT). Microsomes were extracted and assayed for the rate of cyclosporine A oxidation by HPLC.
Relative levels of CYP3A4 protein and mRNA, and rates of cyclosporine A oxidation in microsomes prepared from six different human hepatocyte cultures
Effect of Eletriptan on Expression of CYP3A4 mRNA in Human Hepatocytes.
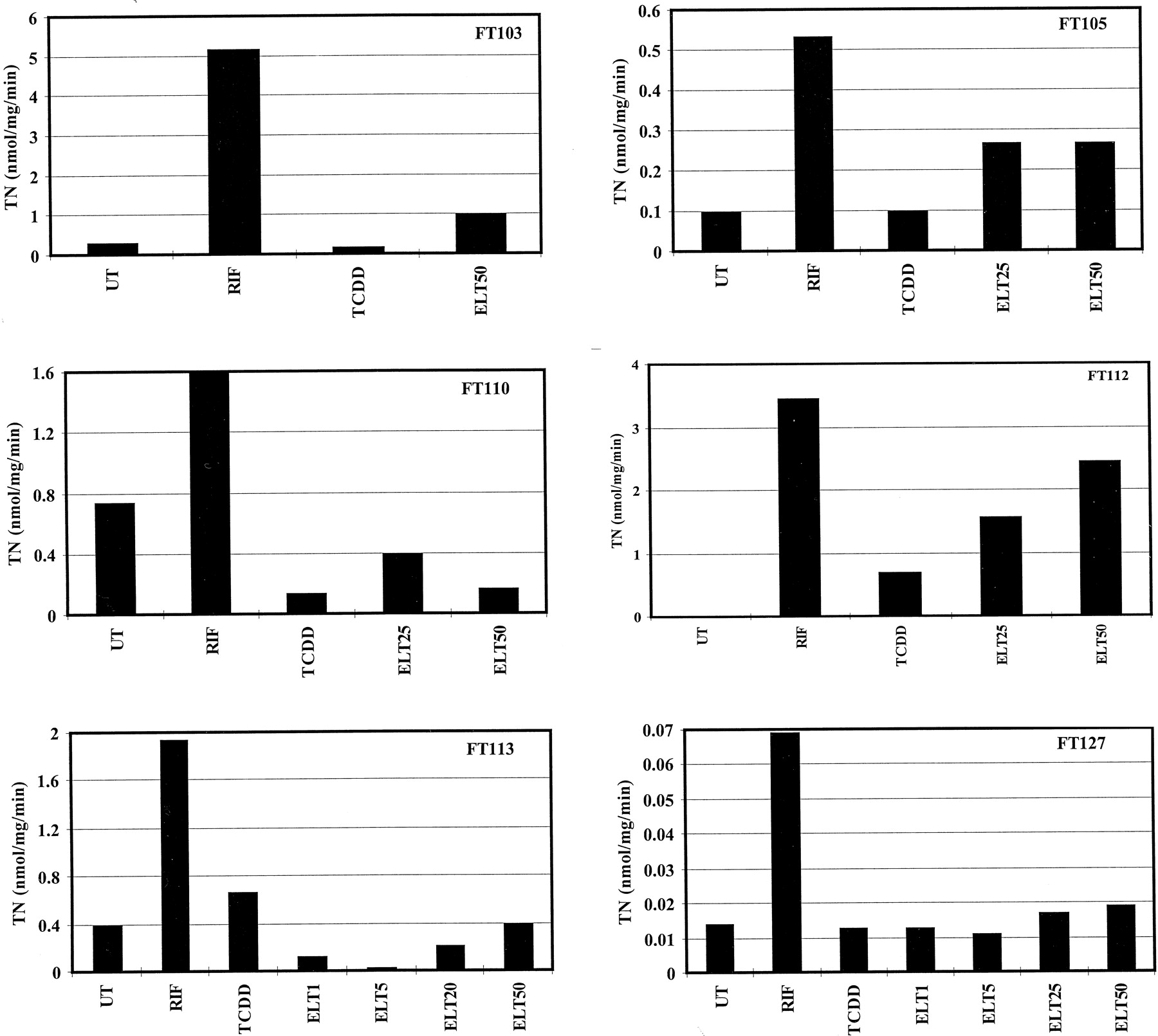
Next, the effect of eletriptan on the expression of CYP3A4 mRNA was investigated with the RNase protection assay (Melton et al., 1984;Muntanet-Relat et al., 1995) in cultures FT112 (in which the protein was induced) and FT110 and FT113 (where it was not). Again, RIF and TCDD were used for comparison. In preliminary experiments, the quality of RNA samples prepared from the cultures after 96 h of treatment was assessed by Northern blot with a human GAPDH cDNA probe. The results reported in Fig. 3A show that GAPDH mRNA was constitutively expressed in all cultures and its level was not affected significantly by any of the treatments. CYP3A4 mRNA was clearly induced in all cultures treated by RIF but not by TCDD (as expected) (Fig. 3B). Interestingly, CYP3A4 mRNA was induced by eletriptan not only in culture FT112 (in which the protein was induced) but also in cultures FT110 and FT113 (where it was not). As observed with the protein and the rate of cyclosporin A oxidation (Figs. 1 and2), the level of CYP3A4 mRNA was concentration dependent, induction becoming detectable at concentrations >5 μM and the level continuing to increase in the range of 25 to 50 μM. It also appears that in the three cultures tested, the maximum level of CYP3A4 mRNA in cells treated with eletriptan was similar to, or even greater (e.g., in cultures FT110 and FT113; Table 1) than that observed in cells treated with RIF, in contrast to the observations made at the protein and activity levels. However, because induction in mRNA without an induction of protein or activity will not result in a clinical response, more emphasis was placed on effects at the protein and activity levels.

Quantification of GAPDH and CYP3A4 mRNAs in human hepatocytes in primary culture after various treatments.
Hepatocytes from patients FT110, FT112, and FT113 were cultured for 96 h in the absence (UT) or in the presence of 1 nM TCDD; 25 μM RIF; or 1, 5, 25, or 50 μM eletriptan (ELT). Total RNA was extracted and quantified by UV-spectrum. One hundred micrograms of RNA was assayed to quantify the GAPDH mRNA by Northern blot (A) and CYP3A4 mRNA by RNase protection (B). SM, standard markers: QX174 digested byHaeIII for RNase protection FT110, and PM2 digested byHindIII for RNase protections FT112 and FT113; probe, native undigested cDNA probe; UT, untreated cells; RIF, cells treated with 25 μM RIF; TCDD, cells treated with 1 nM TCDD; ELT1, ELT5, ELT25, and ELT50, cells treated with 1, 5, 25, or 50 μM eletriptan, respectively. The protected probe migrates slightly ahead of the native probe because of the removal of a fragment of plasmid DNA in the native probe during digestion by RNases.
Effect of Eletriptan on Rate of Degradation of CYP3A4 Protein in Human Hepatocytes.
The disagreement between the levels of CYP3A4 mRNA and protein in response to eletriptan in hepatocyte cultures could result from an increased rate of degradation of the protein by this molecule. To test this possibility, hepatocytes (culture FT142) were preinduced for 96 h with 10 μM RIF. At this time, referred to as time zero in this experiment, the cells were extensively washed with fresh culture medium to remove the inducer and the culture was allowed to continue for 72 h in the absence of treatment, in the presence of 10 μM RIF, or in the presence of 25 μM eletriptan. After 24, 48, and 72 h, the levels of CYP3A4 protein were measured in microsomes prepared from the cells by immunoblot. Data are reported in Table2. In this culture, control experiments revealed that the levels of CYP3A4 mRNA were not significantly different in response to RIF and eletriptan, whereas the level of CYP3A4 protein in eletriptan-induced cells was <2% of that measured in RIF-induced cells (see legend, Table 2). During the washout of RIF, the levels of CYP3A4 protein were not significantly different in eletriptan-treated cells and in untreated cells, suggesting that this molecule does not accelerate the rate of decay of CYP3A4 protein after removal of the inducer. A similar conclusion was reached after analyzing data obtained with another culture (culture FT151, data not shown).
Effect of eletriptan on the rate of decay of CYP3A4
Effect of Eletriptan on Expression of CYP1 and CYP2 Proteins and Some Related Activities in Human Hepatocytes.
The effect of eletriptan on immunodetectable levels of CYP1A2, CYP2A6, CYP2C9, CYP2C19, CYP2D6, and CYP2E1, as well as on some related monooxygenase activities, including ethoxyresorufinO-deethylation (CYP1A) and coumarin 7-hydroxylation (CYP2A6), was investigated in parallel in the different cultures. In all cultures tested, CYP1A2 and ethoxyresorufinO-deethylation (CYP1A1 and CYP1A2)were induced by TCDD but neither by RIF (as expected) nor by eletriptan (CYP1A1/1A2, data not shown). In addition, the levels of proteins CYP2C9, CYP2C19, CYP2D6, and CYP2E1 were not affected by eletriptan (data not shown). CYP2A6 was induced by RIF, as reported previously (Dalet-Beluche et al., 1992), as well as by eletriptan at 25 and 50 μM in cultures FT103 and FT110 (Fig. 4). This protein was not detectable in the other cultures. The rate of coumarin 7-hydroxylation, a CYP2A6-dependent activity, also was enhanced in culture FT103, as expected, but was not detectable in culture FT110 (RIF or eletriptan treated), nor was activity measurable in the other cultures in which CYP2A6 protein was not detected. This is not the first time we observe, in some cultures, an increased expression of CYP2A6 without associated coumarin hydroxylase activity. We have no explanation for this observation, although it could be related to some genetic defect.

Quantification of CYP2A6 protein and rate of coumarin 7-hydroxylation in microsomes prepared from primary hepatocyte cultures after various treatments.
Hepatocytes from patients FT103 and FT110 were cultured for 96 h in the absence (UT) or in the presence of 1 nM TCDD, 25 μM RIF, or 25 or 50 μM eletriptan (ELT). Microsomes were extracted and 100 μg was assayed by immunoblot (top and middle blots) with anti-CYP2A-specific antibodies and revealed by the ECL method (Amersham Corp.). Authentic standard (STD) was a sample of microsomes (50 μg) from a human lymphoblastoid cell line transfected with a human CYP2A6 cDNA (Gentest). ∗, nonspecific bands. Bottom, microsomes were extracted and assayed for the rate of coumarin 7-hydroxylation.
Effect of Eletriptan on 6β-Hydroxycortisol/Cortisol Excretion.
Previous work showed that the ratio of urinary concentrations of 6β-hydroxycortisol/cortisol was a good marker of CYP3A4 induction in vivo (Park 1981; Ged et al., 1989). Therefore, this method was used herein to evaluate the effect of eletriptan on the status of hepatic CYP3A4 in healthy male volunteers. After both 40 mg t.i.d. and 80 mg b.i.d., the adjusted mean ratio of 6β-hydroxycortisol/cortisol at day 7 compared with the predose was not significantly altered (Table3). Furthermore, after both 40 and 80 mg, the ratios postdose on day 1 (6.9 and 3.75, respectively) were slightly lower than those observed at predose on day 1 (7.54 and 7.88, respectively), but neither comparison was statistically significant.
6β-Hydroxycortisol/cortisol ratios after dosing with eletriptan
Discussion
The present finding that, at high concentrations in vitro, eletriptan is a weak inducer of CYP3A4 protein in at least four of the six cultures tested, raises two questions. First, is this induction likely to occur in humans in vivo? And second, if yes, is it likely to be clinically significant? According to previous observations, there is no reason to conclude that induction of a CYP protein in response to a drug, as assessed herein, is restricted to hepatocytes in primary culture and should not occur in vivo (Diaz et al. 1990; Pichard et al., 1990, 1992; McDonnell et al., 1992; Rost et al., 1992; Maurel, 1996b;Chang et al., 1997). However, the actual concentration to which hepatocytes are being exposed in vivo may be very different from the situation in vitro. This is a critical point in what constitutes an inducer in vivo. In the test cultures, the lowest concentration at which eletriptan produces a detectable induction of CYP3A4 protein is between 5 and 25 μM, as reported in Table 1 and Figs. 1 to 3. This is an order of magnitude greater than the actual plasmaCmax of eletriptan after one oral dose of 80 mg (∼0.5 μM) in normal healthy volunteers (Milton et al., 1997). Therefore, this compound is not likely to have any significant clinical affect on the level and activity of CYP3A4 protein in vivo. Eletriptan is unlikely to accumulate in the liver and produce intracellular concentrations similar to those obtained in hepatocyte culture (i.e., >5 μM, the lowest concentration producing CYP3A4 mRNA induction) because the half-life of this compound is ∼4.5 h and its pharmacokinetics is essentially linear. Moreover, the intended dosing regimen for the treatment of acute migraine will result in only intermittent exposure to eletriptan.
Although the concentration of RIF routinely used herein was 25 μM compared with peak plasma concentrations of 1 to 4 μM for a standard 600-mg dose of RIF, recent investigations have shown that the levels of CYP3A4 protein (as well as related activities) and mRNA reach a maximum in response to 1 to 10 μM of this inducer in cultures (Chang et al., 1997; Greuet et al., 1997). The data presented in Table 1 and Figs. 1to 3 suggest, therefore, that eletriptan is a much less potent inducer of CYP3A4 mRNA and protein than RIF. Indeed, the dose of eletriptan required to induce CYP3A4 mRNA, protein, and cyclosporin A oxidation appears to be ∼10 times greater than that of RIF (a dose of 25 to 50 μM eletriptan is required for the maximum induction), and also the amplitude of induction observed in response to this molecule (Table 1) is significantly lower than that observed in response to RIF. The discordance observed between CYP3A4 mRNA and protein expression (i.e., induction of CYP3A4 mRNA in two cultures where CYP3A4 protein was not induced) in response to eletriptan was shown not to result from an increased rate of degradation of the protein in the presence to eletriptan (Table 2). Currently, we have no further explanation for this observation. The in vitro model would therefore predict that unlike the potent CYP3A4 inducer RIF, which clinically achieves plasma concentrations known to maximally induce CYP3A4 in vitro, therapeutic doses of eletriptan will not induce CYP3A4 in vivo. This contention has been upheld by the 6β-hydroxycortisol/cortisol data obtained after an eletriptan multiple-dose study in human volunteers. Urinary excretion of 6β-hydroxycortisol is well recognized as an in vivo marker of CYP3A4 activity (Park, 1981; Ged et al., 1989), with several known CYP3A4 inducers such as RIF and phenytoin increasing 6β-hydroxycortisol levels (Roots et al., 1979). In one study of 14 patients, a 3-fold increase in the 6β-hydroxycortisol/cortisol ratio was recorded after 5 days of RIF treatment, with a range of 1.5- to 18-fold (Ged et al., 1989). In the present study, no increase in 6β-hydroxycortisol/cortisol ratio was observed after 7 days of eletriptan dosing (with the highest clinical dose, 80 mg), which suggests that eletriptan does not induce CYP3A4 activity in vivo and supports the in vitro findings. Eletriptan is therefore not expected to increase the metabolism of oral contraceptives and hence decrease their circulating levels. This study therefore precludes the need to perform an oral contraceptive interaction study.
At present, there is no direct data on the extent to which intestinal CYP3A4 is involved in the inactivation of ethynylestradiol. However, grapefruit juice has been reported to increase the oral bioavailability of ethynylestradiol, suggesting a degree of intestinal CYP3A4 metabolism (Weber et al., 1996). Whether inducibility of CYPP3A4 in the intestine parallels its inducibility in the liver is not known, although RIF seems to be an inducer of intestinal CYP3A4 (Kolars et al., 1992), as in the liver.
Eletriptan does not affect the expression of other CYP proteins that are involved in the metabolism of many drugs, including CYP1A2, CYP2C9, CYP2C19, CYP2D6, and CYP2E1. The possibility that eletriptan can induce CYP2A6 as does RIF (Dalet-Beluche et al., 1992) has to be considered. However, this was observed in only two of the six cultures tested, and the pharmacological consequences of this effect are not likely to be important due to the limited contribution of CYP2A6 to the metabolism of drugs (Waxman, 1996). Indeed, only coumarin and fadrozole have been shown to be metabolized primarily by CYP2A6 so far, and although the contribution of this enzyme to the activation of oxazaphosphorines, such as cyclophosphamide and ifosphamide, also has been reported, it is now considered to be minor in regard to that of CYP2B6 and CYP3A4, respectively (Chang et al., 1997). Interestingly, induction of CYP2A6 by eletriptan was observed in culture FT110 in which CYP3A4 protein was not induced. It appears, therefore, that although these two genes respond to the same inducers, CYP2A6 and CYP3A4 protein expression are not coregulated.
In summary, eletriptan is only a relatively weak inducer of CYP activity in vitro, and at clinical doses does not induce cortisol metabolism in vivo. The data therefore suggest that eletriptan is extremely unlikely to produce clinically significant induction of CYP proteins in humans in vivo.
Footnotes
-
Send reprint requests to: Dr. Patrick Maurel, Institut National de la Santé et de la Recherche Médicale U128, Centre National de la Recherche Scientifique IFR24, 1919 Route de Mende, 34293 Montpellier (Cedex 05), France. E-mail:maurel{at}u128.crbm.cnrs-mop.fr
- Abbreviations used are::
- CYP
- cytochrome P-450
- RIF
- rifampicin
- TCDD
- 2,3,7,8-tetrachlorodibenzo(p)dioxin
- DMSO
- dimethyl sulfoxide
- ECL
- enhanced chemiluminescence
- GAPDH
- glyceraldehyde 6-phosphate dehydrogenase
- Received July 9, 1999.
- Accepted September 27, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}