


Abstract
The metabolism of tresperimus, a new immunosuppressive agent, was investigated in vivo and in vitro in rat and in human. Two metabolic pathways were identified at each side of the molecule with two deamination reactions on the spermidine moiety and hydrolysis of the amide bond leading to the liberation of guanidinohexylamine. As the major metabolic pathway of the drug seemed to be the oxidative deamination, the capacity of different amine oxidases to metabolize tresperimus was then tested using in vivo experiments in rat and in vitro studies in rat and human plasma. The increase of tresperimus plasma levels induced by the administration of hydralazine, an irreversible in vivo inhibitor of semicarbazide-sensitive amine oxidase (SSAO), reflected the major involvement of this enzyme in tresperimus metabolism. This result was confirmed in vitro in rat and human plasma by the use of semicarbazide, a specific SSAO inhibitor. As opposed to rat plasma, human plasma may be an interesting in vitro model to study the metabolism of a drug extensively metabolized by SSAO such as tresperimus. Indeed, SSAO activity was significantly higher in human plasma than in rat plasma. The second metabolic pathway of the drug, which only occurred in rat plasma, appeared thus as the major route of tresperimus metabolism in this biological matrix.
Tresperimus is a new immunosuppressive compound inducing a specific tolerance in rat cardiac allograft model after a short term treatment. It is effective in the prevention of murine graft-versus-host-disease (Dutartre et al., 1995; Andoins et al., 1996, 1997). Tresperimus belongs to the polyamine class of compounds with a spermidine moiety and a guanidine functionality. After a bolus intravenous administration in rat, tresperimus is rapidly cleared with a half-life of 0.3 h. None of the hepatic in vitro models usually used in drug metabolism studies, such as liver microsomes or cultured hepatocytes, can metabolize the drug. However, LC/MS1 analyses of the biological fluids of animals or men treated with tresperimus were used to identify the main metabolites of the drug (Fig.1). The major metabolic pathway of the drug seems to be the oxidative deaminations of the spermidine moiety especially in man. Since such reactions are usually mediated by amine oxidases, the first investigations focused on this class of enzymes.
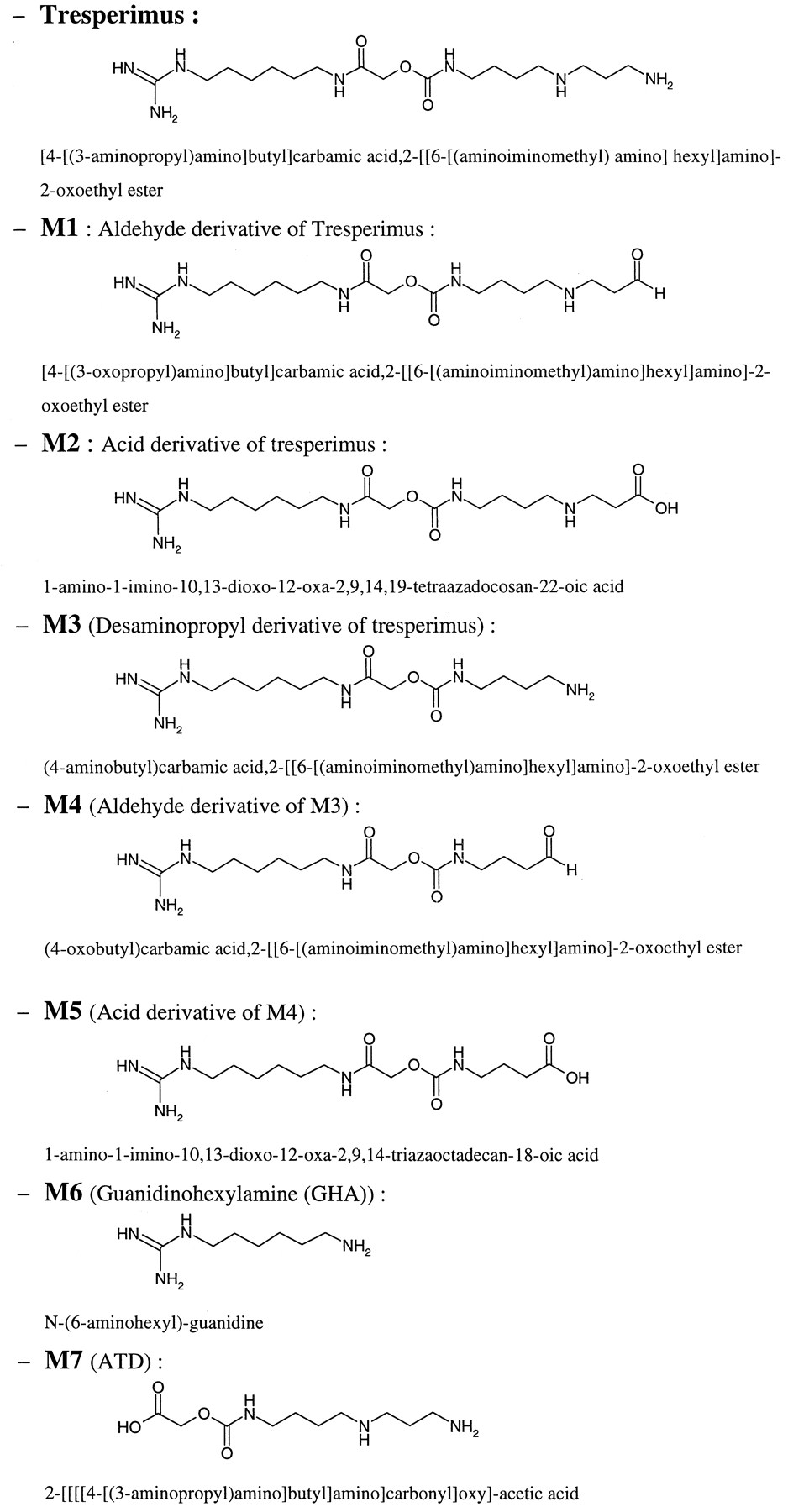
Chemical structure of tresperimus and its proposed metabolites.
The heterogeneous class of amine oxidases can be divided into two types according to the chemical structure of their cofactors. Monoamine oxidases A and B (MAO A and B) and an intracellular form of polyamine oxidase (PAO) contain flavin adenine dinucleotide as prosthetic group, whereas a second group is composed of amine oxidases with an organic cofactor containing one or more carbonyl groups making them sensitive to inhibition by agents such as semicarbazide. After many controversies, this cofactor was identified, first for bovine serum amine oxidase, as topaquinone (Janes et al., 1990). Diamine oxidase, lysyl oxidase, and semicarbazide-sensitive amine oxidase (SSAO) belong to this second group.
These two groups of enzymes also exhibit a different substrate specificity. Indeed, SSAO deaminate only primary amines, in contrast to MAO, which have also the ability to metabolize certain secondary and tertiary amines, or PAO, which act specifically on secondary amino groups (Lyles, 1997). The use of selective inhibitors, of either SSAO or MAO, can help to distinguish between the involvement of these different enzymes in amines metabolism. Indeed, SSAO are largely insensitive to inhibition by 0.1 to 1 mM concentrations of acetylenic aromatic amines such as clorgyline, pargyline, and deprenyl, which produce a complete and irreversible inhibition of both forms of MAO at these concentrations. Conversely, SSAO are sensitive to semicarbazide at concentrations between 0.1 and 1 mM, which have a relatively weak effect on MAO (Lyles, 1997). The antihypertensive agent hydralazine is also a potent irreversible in vivo inhibitor of rat SSAO (Lyles and McDougall, 1989).
In mammals, PAO activity was detected in most tissues as the two MAO forms, the highest activity being in the liver (Seiler et al., 1980). MAO B is also expressed in blood platelets and lymphocytes (Strolin Benedetti et al., 1994). MAO is primarily a mitochondrial enzyme, although some MAO activity has also been found in the microsomal fraction. SSAO has been found in several tissues, with a particularly high activity in blood vessels (Lyles and Singh, 1985; Precious and Lyles, 1988) and in highly vascularized tissues (Lewinsohn, 1984). A soluble form of SSAO has also been identified in blood plasma (McEwen and Cohen, 1963).
MAO and PAO may be considered as endobiotic-metabolizing enzymes; they respectively catalyze deamination of biogenic amines and the interconversion of polyamines (Seiler et al., 1981). On the other hand, the physiological substrates of SSAO are not clearly identified, even if particular attention is given to various endogenous amines such as aminoacetone and methylamine.
The contribution of MAO or PAO to the metabolism of xenobiotics is clearly established (Strolin Benedetti and Tipton, 1998), whereas the involvement of SSAO in drug metabolism has been rarely demonstrated. In fact, few xenobiotics have been identified as substrates of SSAO. Indeed, only allylamine, an industrial toxin (Hysmith and Boor, 1988), 2-propyl-1-aminopentane, a prodrug of valproic acid (Yu and Davis, 1991), and the radioprotective drug WR 105 (Meier and Issels, 1995) undergo a metabolism partly depending on SSAO.
The main goal of this study was to identify the nature of the amine oxidase involved in tresperimus metabolism. First, in vivo experiments in rat were conducted using specific inhibitors of amine oxidases. Since no hepatic in vitro model allows for the study of tresperimus metabolism, the suitability of plasma as in vitro model was tested. Thus, HPLC and LC/MS analyses were used to demonstrate the capacity of rat and human plasma to metabolize the drug. Furthermore, experiments performed with this model underlined the role of SSAO in tresperimus metabolism.
Materials and Methods
Animals and Chemicals.
Male Sprague-Dawley rats (Charles Rivers, Iffa Credo, Cléon, France) weighing 200 to 220 g were used. Human blood was obtained from Bruant Laboratories (Dijon, France). Tresperimus HCl was synthesized at Synkem (Chenôve, France). Metabolites M1, M2, M3, and the internal standards LF 13-0435 and LF 07–0109 were synthesized by Laboratoires Fournier (Dijon, France). Pargyline, hydralazine, semicarbazide, benzylamine, and 5,5-dimethyl-1,3-cyclohexanedione (dimedone) were obtained from Sigma Chemicals (Saint-Quentin Fallavier, France) and Waymouth medium WB 752/1 from Life Technology (Cergy Pontoise, France).
In Vivo Rat Metabolism.
The rats were randomly divided into four groups of 10 animals. Each group received a single 4-mg/kg i.v. administration of tresperimus 1 h after intraperitoneal injection of either pargyline (25 mg/kg), hydralazine (5 mg/kg), or a mixture of pargyline (25 mg/kg) and hydralazine (5 mg/kg), or isotonic saline for the control group. Blood samples were collected at the abdominal aorta into heparinized tubes. Two rats were bled at each of the following times: 0, 0.25, 0.5, 2, and 3 h. Upon completion of blood collection, rats were euthanized. Plasma was then separated by centrifugation at 2400g for 5 min at 4°C. Plasma was dispensed into ice-cooled tubes and acidified with a mixture of citric acid (0.5 M)/NaH2PO4 (0.5 M), 70:30, v/v, added on the basis of 5% (v/v). The samples were then directly used for analysis.
Pharmacokinetics Data Analysis.
The plasma concentration-time data were evaluated using Topfit version 2.0 software. A noncompartmental analysis was used to analyze the plasma concentration-time data. Area under the concentration-time curve was calculated by the linear trapezoidal rule. The elimination rate constant (ke) was estimated from the terminal slope of the log concentration-time curve and the apparent elimination half-life (t1/2) was calculated from ke (i.e.,t1/2 = 0.693/ke).
In Vivo Human Metabolism.
The metabolic profile of tresperimus was investigated by LC/MS in the plasma of healthy volunteers having received 9 mg/kg of tresperimus as a 6-h intravenous infusion.
In Vitro Determination of SSAO Activity in Rat and Human Plasma.
Blood was collected into heparinized tubes and plasma was obtained after centrifugation at 2400g for 5 min at 4°C. The determination of SSAO activity was performed using a sensitive and reproducible method (Van Dijk et al., 1995). Briefly, after 45 min of preincubation with clorgyline at 1 mM to inhibit any MAO activity, plasma was incubated at 37°C with the SSAO substrate benzylamine (2.6 mM for human plasma and 50 μM for rat plasma). The incubation time was 1 h for human plasma and three h for rat plasma. The production of benzaldehyde was linear for these incubation times. After deproteinization, the samples were derivatized with the specific aldehyde reagent dimedone (5,5-dimethyl-1,3-cyclohexanedione) prepared in ammonium acetate (7.8 M) at high temperature (95°C) with a resulting pH of 4.5. The derivatization reaction was based upon the condensation between benzaldehyde, the β-diketone dimedone, and ammonium acetate (Sawicki and Carnes, 1968; Compton and Purdy, 1980). The amount of benzaldehyde formed was linear for these incubation times and quantified by HPLC with fluorometric detection (Mopper and Stahovec, 1983). SSAO activity was expressed in milli-international units per liter, which corresponds to picomoles of benzaldehyde formed per minute per liter of plasma.
In Vitro Metabolism of Tresperimus, M1, and M2.
Tresperimus was incubated at 7.75 μM for 6 h in rat and human plasma. Tresperimus was incubated in rat and human plasma at concentrations close to the Cmax determined in vivo. Concurrently, M1 and M2 were incubated at 10 μM in rat plasma and in Waymouth medium as a control. Waymouth medium was a culture medium composed of a mixture of inorganic salts, vitamins, amino acids, and diverse constituents like glucose or an indicator of pH (phenol red). Waymouth medium was devoid of any reactive proteins. Both compounds were incubated at 37°C for 3 h. Incubations were stopped by cooling the tubes in ice and adding 5% (v/v) of the acidic reagent. The samples were analyzed by LC/MS.
Identification of the Enzyme Involved in the Oxidative Deamination of Tresperimus.
A pool of plasma was freshly prepared from at least 15 human healthy volunteers or 8 rats per experiment. Plasma was preincubated for 45 min at 37°C with either pargyline (1 mM), hydralazine (100 μM), semicarbazide (100 μM), or phosphate-buffered saline buffer as control. The reaction was then initiated by adding tresperimus at a final concentration of 2.6 μM in human plasma and 13 μM in rat plasma. The samples were incubated for 2 h at 37°C. The reaction was stopped by cooling the tubes in ice and adding 5% (v/v) of the acidic reagent. The samples were stored at −80°C pending analysis.
Quantitative Determination of Tresperimus in Plasma.
A HPLC method was used to determine the plasma concentrations of tresperimus. This method is a modification of that used to measure polyamines (Simpson et al., 1982). One milliliter of plasma sample was submitted to a liquid-solid extraction on a C18Bond Elut column before HPLC analysis. After addition of the internal standard (LF 13-0435), tresperimus was eluted with methanol containing 0.5% (v/v) of trifluoroacetic acid. The eluate was evaporated at about 40°C under a gentle stream of nitrogen. The dry residue was then dissolved in 150 μl of hydrochloric acid 0.1 N, and 75 μl were injected into the HPLC system. The HPLC analysis was achieved by using a Superspher 100 C18 column, protected by a Superspher 100 RP 18 guard column (Merck, Nogent sur Marne, France). Column temperature was maintained at 40°C. The mobile phase was composed of reagent A, a mixture of ion pair solution and methanol (75:25, v/v), and reagent B, a mixture of ion pair solution, methanol, and propan-2-ol (65:25:10, v/v/v). The ion-pair solution was 1-heptane sulfonic acid (5 mM) in potassium dihydrogen phosphate (100 mM, pH 2.5). A gradient elution at a flow rate of 1.2 ml/min was used with 0 to 35% reagent B from 0 to 10 min, 35% reagent B from 10 to 17 min, 35 to 0% B from 17 to 17.25 min, and 0% B from 17.25 to 22.5 min. After a post-column derivatization of tresperimus based upon the reaction of the primary amino group of the drug withO-pthaldialdehyde in the presence of 2-mercaptoethanol, the highly fluorescent compound produced was detected by fluorescence (excitation 340 nm, emission 440 nm). The post-column derivatization was performed with a solution in borate buffer (0.75 M) at pH 10.5 containing 0.5 g/l of O-pthaldialdehyde and 500 μl of 2-mercaptoethanol per liter, dispensed at a rate of 0.5 ml/min.
Metabolites Identification.
The LC/MS analysis was used to establish the in vivo metabolic pattern of tresperimus and to measure the effects of different amine oxidase inhibitors on the oxidative deamination of tresperimus. The structural analysis of metabolites in rat and human plasma was performed by electrospray-linked LC/MS (TSQ 7000, Finnigan MAT, San Jose, CA) equipped with HPLC (HP 1100, Hewlett Packard, Waldbronn, Germany). The HPLC analyses were performed at 40°C on an Inertsil ODS 2 column (Interchim, Montluçon, France) for human plasma and a Supelco ABZ column (Supelco, Saint-Quentin Fallavier, France) for rat plasma. The mobile phase was a mixture of water and acetonitrile containing 0.05% (v/v) trifluoroacetic acid. A gradient elution (0–30% acetonitrile) was used with a flow rate of 0.4 ml/min. For the inhibition studies, the detection of the main plasma metabolites of tresperimus was performed in single ion monitoring with atmospheric pressure chemical ionization. The ion monitoring on protonated molecular MH+ was selected for each known metabolite, namely tresperimus, M1, and M3. The results were expressed as the ratios of the peak areas of the metabolite to internal standard. The peak areas were measured in arbitrary units of mass, but the coefficient of response in mass for each metabolite was unknown. Consequently, such a way of expressing the results only allowed us to compare the quantity of the same metabolite in different samples but not to accurately quantify the metabolites.
Results
In Vivo Metabolic Pattern of Tresperimus in Rat and Human Plasma.
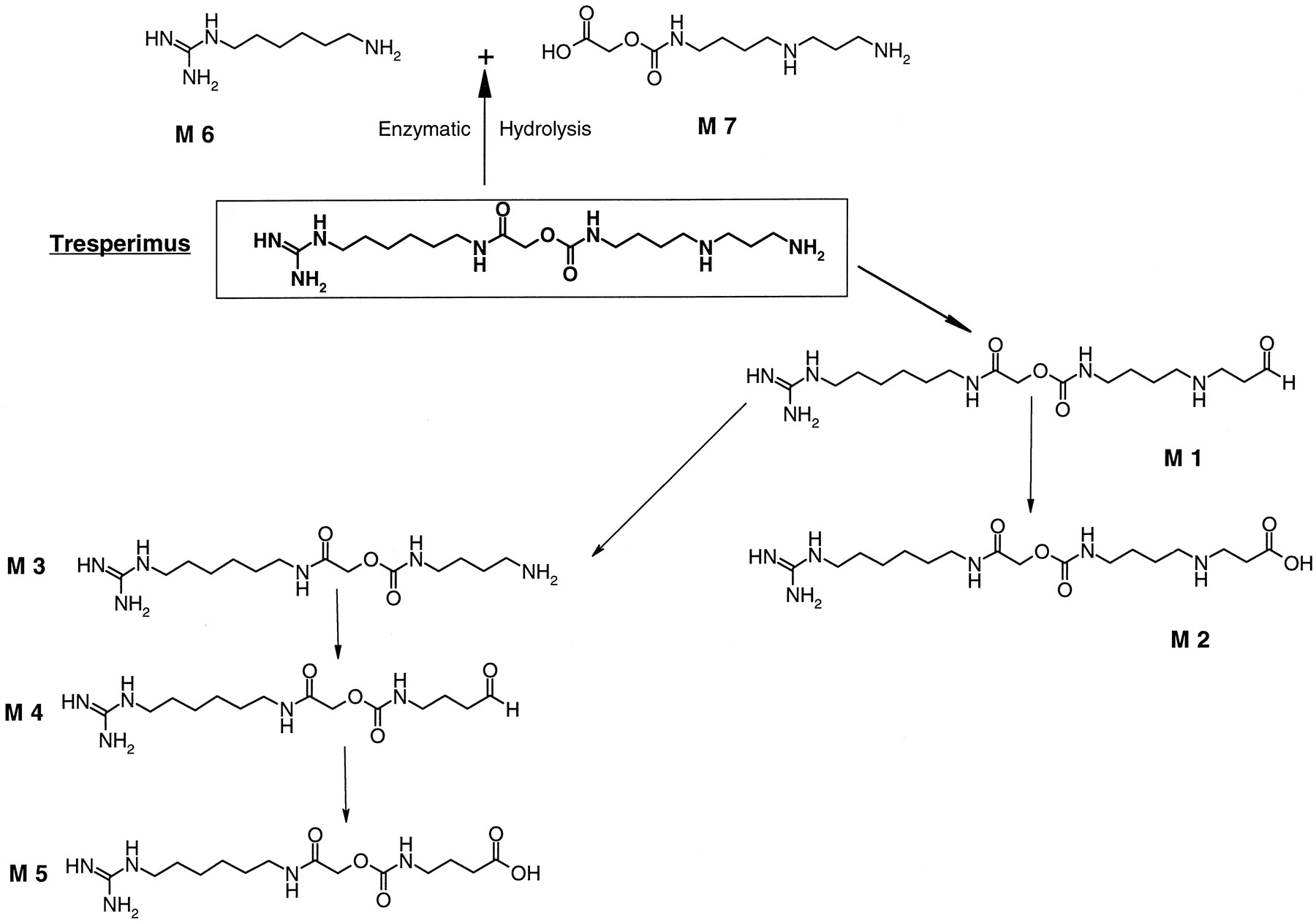
The tresperimus metabolic pattern in rat and human plasma after in vivo administration is presented in Fig. 2. The same metabolites were identified in the plasma of both species. The formed metabolites allowed us to localize two sites of metabolization on the molecule: at the primary amino terminal and at the middle on the amide bond. At the spermidine moiety, the aldehyde (M1) and acid (M2) derivatives of tresperimus were identified. Similar to tresperimus, the oxidized metabolites of M3 were identified with its aldehyde (M4) and its acid (M5) derivatives. At the middle of the molecule, M6 is liberated by hydrolysis of the amide bond.

Possible metabolic scheme of tresperimus in humans and in rat.
Male Sprague-Dawley rats (n = 2) received a single intravenous administration of tresperimus at 4 mg/kg. Plasma was analyzed in LC/MS 0.5 h after the drug administration. Tresperimus (9 mg/kg) was administered to healthy subjects as a 6-h intravenous infusion. Plasma was analyzed 1 h after the end of infusion.
Pharmacokinetics of Tresperimus in Rat Plasma after Inhibition of Different Classes of Amine Oxidases.
Tresperimus was extensively metabolized in vivo in rat. To identify the amine oxidase involved in tresperimus oxidative deamination, rats were first treated with the SSAO inhibitor hydralazine and the MAO inhibitor pargyline. The drug was then administered and its pharmacokinetic profile evaluated. As described in Fig.3, both groups of rats treated with hydralazine, in absence or in presence of pargyline, presented plasma pharmacokinetics different from that of the control. As shown in Table1, hydralazine caused significant increases of the area under the curve (AUC) of the drug. Indeed, the AUC of the drug was 1.75 times higher than in control, andt1/2 was increased by 2.33. These effects were stronger when pargyline was administered simultaneously with hydralazine. In contrast, the administration of pargyline had only a weak effect, with an AUC only 1.17 times higher than that of the control.

Plasma pharmacokinetics of tresperimus after single i.v. injection (4 mg/kg) in rat previously treated or not (control) with amine oxidase inhibitors.
Plasma pharmacokinetics of the unchanged drug after single i.v. administration of tresperimus (4 mg/kg), 1 h after intraperitoneal administration of amine oxidase inhibitors, 5 mg/kg hydralazine, 25 mg/kg pargyline, a mixture of hydralazine + pargyline, and physiological serum for the control (10 animals per group, 2 different animals at each collection time). Values are means of the tresperimus plasma concentration. The bars represent the minimal and maximal concentrations.
Pharmacokinetic parameters of tresperimus after i.v. injection (4 mg/kg) in rat previously treated or not (control) with amine oxidase inhibitors
Determination of SSAO Activity and in Vitro Metabolic Pattern of Tresperimus in Rat and Human Plasma.
SSAO seemed to be predominantly involved in tresperimus oxidative deamination in rat. SSAO being present in plasma, the suitability of plasma to study in vitro tresperimus metabolism, especially in man, was then investigated. Rat and human plasma SSAO activities were first determined with benzylamine as substrate to evaluate the metabolic capacity of these biological matrices. The measured SSAO plasma activity was 631 ± 241 mIU/l in 20 human healthy volunteers (50 ± 5 years old). This plasma activity was only 24 ± 5.5 mIU/l in eight CD Sprague-Dawley male rats (7 weeks old). The SSAO activity in rat plasma was 26 times lower than the SSAO activity in human plasma.
As shown in Table 2, after in vitro incubation of tresperimus in rat or human plasma, all the metabolites resulting from oxidative reactions on the spermidine moiety were present except the acids. Despite a tresperimus disappearance higher in rat plasma than in human plasma, M1, M3, and M4 were clearly formed at lower concentrations in rat than in human. These rates of tresperimus oxidation in rat and human plasma seemed consistent with the different benzylamine deaminating activities assayed in these biological matrices. M6 was only detected in rat plasma.
Metabolic profile of tresperimus after incubation in rat and human plasma for 6 h at 37°C
In Vitro Incubation of M1 and M2.
Tresperimus is deaminated into the corresponding aldehyde, M1, which is then oxidized in M2 (Fig. 1). These two metabolites could be at the origin of the M3 liberation. Such reaction could result from a nonenzymatic mechanism rather than from an enzymatic activity. To explore the mechanism of M3 formation and to exclude any enzymatic intervention, M1 and M2 were incubated in Waymouth medium and in rat plasma.
The results reported in Table 3 seemed to confirm the M3 formation by a nonenzymatic mechanism from M1. Indeed, M3 appeared after M1 incubation in Waymouth medium as well as in rat plasma, whereas M3 was not formed during M2 incubation in Waymouth medium as well as in rat plasma. Furthermore, M1 was not spontaneously oxidized into the corresponding acid in both incubation media tested. The formation of M6 occurred only when M1 or M2 was incubated in rat plasma.
Metabolic profile of M1 and M2 after 3-h incubation at 37°C in rat plasma or in Waymouth medium
Effect of Specific Amine Oxidase Inhibitors on the Oxidative Deamination of Tresperimus in Rat and Human Plasma.
Tresperimus oxidative deamination occurs in vitro in rat and human plasma, and these biological matrices can be used to identify the amine oxidase responsible for this reaction. The tresperimus deamination could be evaluated by the disappearance of the unchanged product as well as by the formation of M1 and M3. According to Table4, the rates of tresperimus metabolism determined by HPLC and by LC/MS analyses were very close. The tresperimus deamination was almost suppressed in human plasma by semicarbazide or hydralazine at 100 μM. No effect of the amine oxidase inhibitors on the rate of tresperimus metabolism was observed in rat plasma. Concurrently, as shown in Fig.4, LC/MS analysis demonstrated an inhibition of M1 and M3 formation after preincubation with semicarbazide or hydralazine, in human plasma as well as in rat plasma. The inhibition was almost complete with semicarbazide in plasma of both species. On the other hand, hydralazine inhibited the tresperimus oxidation in human plasma by 98% but only by 65% in rat plasma. After preincubation with pargyline, the formation of these two metabolites in rat as well as in human plasma was decreased by about 25% compared with the control.
Effect of inhibitors of amine oxidases on the disappearance of tresperimus in rat and human plasma

Effect of inhibitors of amine oxidase on the oxidative deamination of the tresperimus in rat and human plasma.
The aldehyde derivative of tresperimus (M1) and desaminopropyl tresperimus (M3) were evaluated by LC/MS analysis as described underMaterials and Methods. Values are means ± S.D. (n = 3) of the percentage of metabolites formed (expressed as the sum of the relative quantities of M1 and M3 compared with a control without inhibitor). ■, rat plasma; ▥, human plasma.
Discussion
After in vivo administration of tresperimus, no significant difference between rat and human was observed in the metabolic pattern of the drug. Two metabolic pathways were identified at the primary amino terminal and at the middle, on the amide bond of the molecule (Fig. 2).
First, the spermidine moiety of the drug underwent oxidative deamination reactions. The primary amino terminals of tresperimus and M3, the desaminopropyl tresperimus, were deaminated leading to oxidized metabolites M1 and M4 identified in rat and in human plasma after intravenous administration. The identification of M1 and M4 was unequivocally confirmed by LC/MS/MS studies, and the NMR analysis demonstrated that the aldehyde derivatives of tresperimus and its M4 metabolite were present as hydrated forms. As shown by the incubation of M1 in Waymouth medium, this metabolite could be at the origin of the M3 formation by a nonenzymatic process.
The second metabolic pathway of the drug took place on the amide bond, close to the guanidine extremity, which was hydrolyzed, leading to the formation of M6. After in vivo administration of tresperimus, the acid derivative of M6 was also detected in rat and in human urine (data not shown). During the incubation of tresperimus and its oxidized metabolites in Waymouth medium, M6 was never detected. These results suggested that M6 is not spontaneously formed in physiological conditions and that the hydrolysis of the amide bond of tresperimus in rat plasma might be enzymatic. The formation of M6 was never observed in human plasma in vitro, but was observed in vivo in plasma of healthy volunteers.
The oxidative deamination seemed to take a major part in the degradation of the drug in rat, since the administration of hydralazine significantly increased tresperimus plasma concentrations in rat. Hydralazine is a potent irreversible in vivo inhibitor of rat SSAO but not MAO. On the other hand, pargyline, which is an irreversible inhibitor of MAO, does not modify the tresperimus plasma concentrations. These results suggest the major involvement of SSAO in the tresperimus biotransformation on the primary amine, which would be the main site of tresperimus metabolism. In vitro experiments with specific SSAO inhibitors, in human and in rat plasma, confirm the in vivo results. Semicarbazide almost completely inhibited the oxidative deamination of the drug in rat and human plasma (Fig. 4). However, in rat plasma, semicarbazide did not modify the drug disappearance despite a strong inhibition of the tresperimus oxidative deamination. These findings suggest that, in rat plasma, the contribution of the oxidative deamination is a minor pathway compared with the hydrolysis of the amide bound leading to the liberation of M6. This is confirmed by the lower formation of tresperimus-oxidized metabolites in rat plasma than in human plasma (Table 2). This result is in accordance with SSAO benzylamine oxidase activity being significantly lower in rat plasma than in human plasma. A similar inhibition of tresperimus metabolism was obtained with hydralazine in human plasma. Hydralazine is a potent time-dependent inhibitor of SSAO, but in vitro hydralazine can also reversibly inhibit the two MAO forms (Lyles et al., 1983). As no effect of pargyline on tresperimus metabolism was detected in human plasma, the hydralazine effect seems to be due to the inhibition of plasma SSAO. SSAO in rat plasma could be less sensitive to hydralazine than human plasma SSAO. Such interspecies differences in sensitivity to SSAO inhibitors have already been reported for bovine serum amine oxidase and human serum SSAO (Yu et al., 1994).
The small increase of tresperimus plasma concentrations after in vivo administration of pargyline might reflect a minor involvement of MAO in tresperimus metabolism. The increase in AUC was greatly potentiated by the coadministration of MAO and SSAO inhibitors. A MAO activity could act upon primary and secondary amine. The deamination of secondary amine by MAO could lead to the formation of M3 or M4. However, the involvement of a PAO activity should not be excluded as pargyline is a specific inhibitor of MAO but might also have a weak inhibitory effect on PAO at high concentrations (Hölttä, 1977). A PAO activity could take place in the intracellular compartment after theN-acetylation of the primary amine of the substrate, leaving the internal nitrogen to be attacked by PAO (Bolkenius and Seiler, 1981). Therefore, the terminal primary amine of tresperimus was acetylated. This N-acetylated derivative of tresperimus was administered in vivo. No metabolites resulting from the oxidation of the spermidine moiety of this compound and especially M3 were detected in urine (data not shown). The involvement of PAO thus appeared to be very unlikely. Furthermore, the weak effect of pargyline could also result from in vivo metabolism of this monoamine oxidase inhibitor. Indeed, pargyline undergoes an extensive hepatic metabolism with a minor metabolic pathway leading to the formation of propargylamine (Weli and Lindeke, 1985; DeMaster et al., 1986), a potent SSAO inhibitor (Precious et al., 1988). The formation of a low amount of propargylamine could thus be indirectly responsible for the slight inhibitory effect of pargyline on tresperimus metabolism in vivo. Pargyline seemed to decrease the in vitro formation of the metabolites M1 and M3 in rat and in human plasma. This effect was very weak and was only revealed by the semiquantification of M1 and M3 performed by LC/MS analysis. LC/MS analysis is very sensitive and directly dependent on the mass coefficient response of each metabolite. Thus, M1 and M3 could have a high coefficient of response facilitating the determination of an inhibition of the oxidative deamination. However, the involvement of MAO should be very weak, since the presence of pargyline in human plasma has no effect on the rate of tresperimus metabolism (Table 4).
Furthermore, a significant increase of tresperimus plasma concentrations was observed in one patient with graft-versus-host disease, treated simultaneously with isoniazid and tresperimus. Isoniazid, an antitubercular drug, inhibits plasma and tissue-bound SSAO with a relatively weak effect upon MAO (Blaschko, 1962; Lewinsohn et al., 1978). This clinical result confirms the major role of SSAO in the biotransformation of tresperimus in humans.
As no hepatic metabolism of tresperimus was observed in vitro, plasma provided an interesting tool to study tresperimus metabolism at physiologically relevant drug concentrations. However, rat and human plasma exhibited marked interspecies differences. Indeed, rat plasma allowed a correct qualitative prediction of in vivo tresperimus metabolism, since both metabolic pathways of the drug were present in this biological matrix, unlike human plasma. However, the acid derivatives of tresperimus and its M3 metabolite were not produced in the plasma of both species. As shown by incubation of M1 in physiological conditions, the conversion of this product into the corresponding acid did not result from a chemical oxidation. Consequently, this oxidation reaction resulted from an enzymatic activity and most likely from an aldehyde dehydrogenase. Approximately 99% of the aldehyde dehydrogenase activity in blood is in the intracellular fraction of the erythrocytes, and this activity is not detected in plasma (Helander and Tottmar, 1986). Therefore, the oxidation of the aldehyde is unlikely to occur in plasma. However, from a quantitative point of view, in vitro metabolism of tresperimus in rat plasma did not seem to reproduce the in vivo situation, as the oxidative deamination of tresperimus occurred to a much lesser extent than the liberation of M6. On the other hand, human plasma could provide an interesting tool to study the metabolism of drugs highly deaminated by SSAO, such as tresperimus. Since tresperimus is abundantly deaminated in human plasma, this in vitro model could contribute to a better understanding of tresperimus metabolism by the determination of its enzymatic kinetic parameters or the assessment of drug interaction potential.
In rat, the discrepancy between in vivo and in vitro results suggests that soluble enzyme is probably not able to account for this metabolism. The SSAO activity in plasma is lower than the enzymatic activity leading to the formation of the metabolite M6. Conversely, smooth vascular cells are described as one of the tissues with the highest specific SSAO activity (Lewinsohn, 1984). Therefore, the involvement in tresperimus metabolism of membrane-bound SSAO, especially in smooth vascular cells, should be investigated.
The oxidative deamination of the primary amine of tresperimus, leading to the formation of M1, seems to play a major role in tresperimus metabolism. This reaction is predominantly catalyzed by SSAO in humans as well as in rat. However, a minor MAO contribution should not be excluded. Some metabolites of tresperimus, like M3 or M6, seemed to undergo an oxidative deamination, as shown in the proposed metabolic pattern of tresperimus (Fig. 5). From their structure, these metabolites could be potential substrates of amine oxidases, as well as SSAO or MAO. If so, tresperimus would not only be one of the first xenobiotics found to be extensively metabolized by SSAO, but also a drug for which amine oxidases could be involved in several steps of its metabolic pathways.
Footnotes
-
Send reprint requests to: Romuald Lainé, Head of Drug Metabolism, Laboratoires Fournier S.A., 50 route de Dijon, 21121 Daix, France. E-mail: r.laine{at}fournier.fr
- Abbreviations used are::
- LC/MS
- liquid chromatography/mass spectrometry
- AUC
- area under the curve
- MAO
- monoamine oxidase
- PAO
- polyamine oxidase
- SSAO
- semicarbazide-sensitive amine oxidase
- HPLC
- high-performance liquid chromatography
- Received July 27, 2000.
- Accepted January 30, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}