


Abstract
Deltamethrin (DLM) is a relatively potent and widely used pyrethroid insecticide. Inefficient detoxification has been proposed to be the primary reason for the greater sensitivity of immature rats to the acute neurotoxicity of DLM. The objective of this study was to test this hypothesis by characterizing the age dependence of DLM metabolism in vitro, as well as toxic signs and blood levels of the neurotoxic parent compound following administration of 10 mg DLM/kg p.o. in glycerol formal. Metabolism was quantified in vitro by monitoring the disappearance of the parent compound from plasma [via carboxylesterases (CaEs)] and liver microsomes [via CaEs and cytochromes P450 (P450s)] obtained from 10-, 21-, and 40-day-old male Sprague-Dawley rats. Mean (±S.E.) intrinsic clearances (Vmax/Km) in these respective age groups by liver P450s (4.99 ± 0.32, 16.99 ± 1.85, and 38.45 ± 7.03) and by liver CaEs (0.34 ± 0.05, 1.77 ± 0.38, and 2.53 ± 0.19) and plasma CaEs (0.39 ± 0.06, 0.80 ± 0.09, and 2.28 ± 0.56) increased significantly (p ≤ 0.05) with age, because of progressive increases in Vmax. Intrinsic clearance of DLM by plasma CaEs and liver P450s reached adult levels by 40 days, but clearance by liver CaEs did not. Hepatic P450s played the predominant role in DLM biotransformation in young and adult rats. The incidence and severity of neurotoxic effects varied inversely with age. Correspondingly, blood DLM areas under the concentration versus time curve (AUCs) and Cmax values progressively decreased with increasing age. Internal exposure to DLM (blood AUCs) was closely correlated with toxic signs (salivation and tremors). The present study provides evidence that the limited metabolic capacity of immature rats contributes to elevated systemic exposure and ensuing neurotoxic effects of DLM.
Pyrethroid insecticides are widely used in forestry, agriculture, veterinary and human medicine, and household pest control (ATSDR, 2003; Soderlund et al., 2002). Pyrethroids did account for ∼25% of the worldwide insecticide market (Casida and Quistad, 1998). However, use of pyrethroids in the United States has substantially increased since 1998, because of the decreased use of organophosphorus pesticides. Exposures to pyrethroids have been documented in several potentially sensitive subpopulations, including pregnant women, infants, and children (Whyatt et al., 2002; Yanez et al., 2002; Berkowitz et al., 2003; Heudorf et al., 2004). Heudorf et al. (2004) measured pyrethroid metabolites in the urine of 1177 persons without apparent exposure in Germany.
Pyrethroids are divided into two categories, types I (which do not contain a cyano group; tremor and parathesias are the major signs of poisoning) and II (which contain a cyano group; choreoathetosis and salivation are the major signs of poisoning), based on their chemical structure and toxicity (Glickman and Casida, 1982). DLM, a type II pyrethroid, is a relatively potent neurotoxicant. Major signs of acute poisoning include salivation, hyperexcitability, choreoathetosis, and seizures. Interaction with neuronal voltage-sensitive sodium channels is the primary mode of action of DLM (Soderlund et al., 2002). The parent compound is believed to be the proximate toxicant, as demonstrated by correlation of brain DLM levels with the onset of symptoms of poisoning in rats (Rickard and Brodie, 1985). Intracerebral injection of DLM produced signs of neurotoxicity within a minute or less in mice (Lawrence and Casida, 1982).
Immature animals are more sensitive to the acute neurotoxicity of pyrethroids than adults. The susceptibility of rat pups to cypermethrin (type II) and permethrin (type I) lethality was found to be inversely related to age (Cantalamessa, 1993). Cantalamessa, (1993) speculated that incomplete development of detoxification enzymes was probably responsible for the age dependence, although his experiments with monooxygenase and esterase inhibitor pretreatments were inconclusive. Sheets et al. (1994) also noted that preweanling pups were much more susceptible to acute DLM toxicity than were adults; oral LD50 values for 11-, 21-, and 72-day-old rats were reported to be 5, 11, and 81 mg DLM/kg. At the time of death, comparable brain DLM concentrations were present in weanling and adult rats that received 12 and 80 mg DLM/kg p.o., respectively. Sheets (2000) proposed that the greater sensitivity of rat pups to lethal doses of type II pyrethroids was due to limited metabolic capacity, although no data on the age dependence of biotransformation of pyrethroids were located.
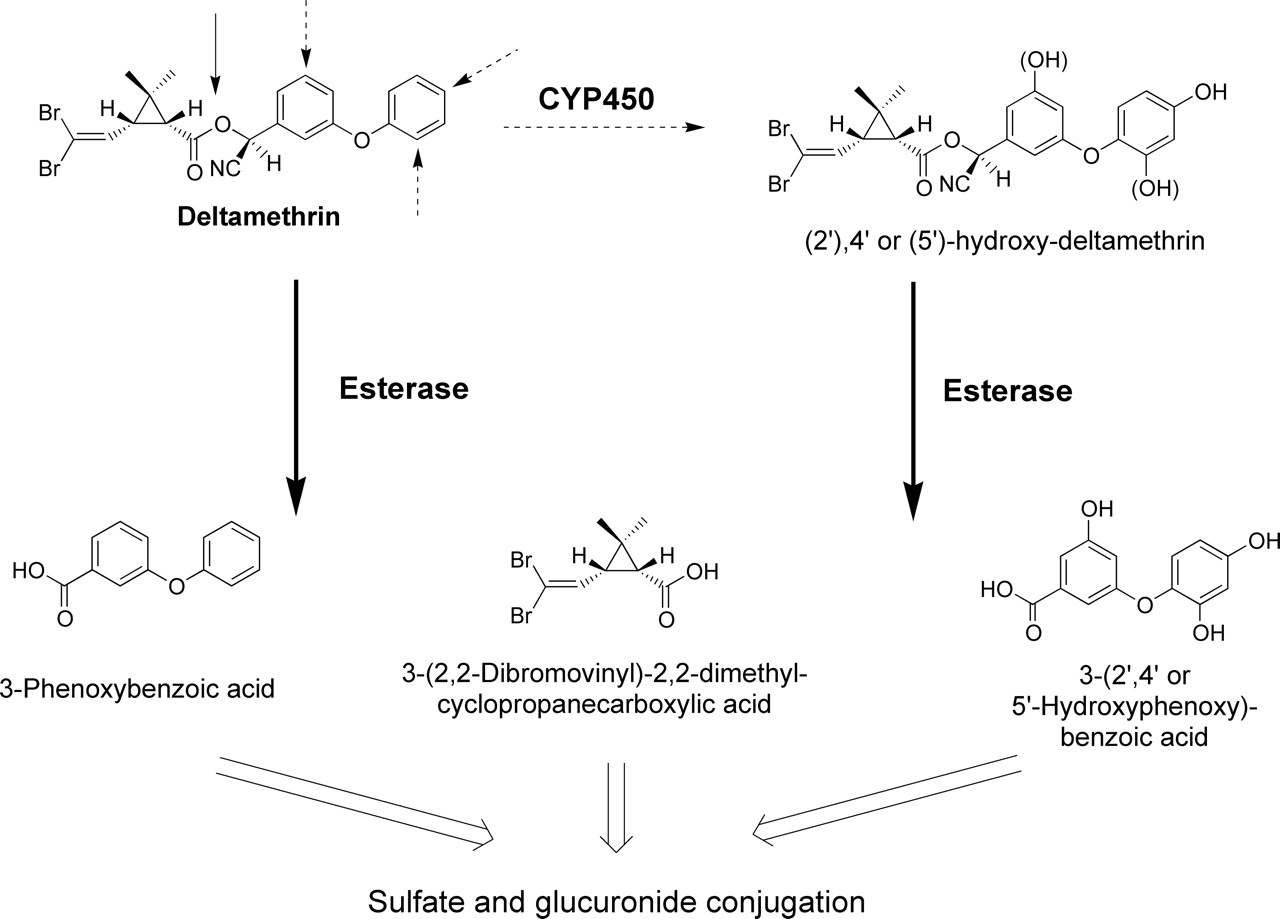
Metabolism of DLM. It is generally accepted that DLM is detoxified by P450-mediated oxidation (dotted arrows) and esterase-mediated hydrolysis (solid arrows), followed by conjugation. Reprinted from Toxicol Appl Pharmacol Anand SS, Bruckner JV, Haines WT, Muralidhara S, Fisher JW, and Padilla S Characterization of deltamethrin metabolism by rat plasma and liver microsomes. In press, 2005 with permission from Elsevier.
DLM has been studied in adults, but information on its metabolism and disposition in immature laboratory animals and humans is very limited. DLM appeared to be rapidly, but poorly absorbed from the gastrointestinal (GI) tract of adult rats, disparately distributed in different brain regions, and oxidized in the liver to 4′-hydroxy DLM (Anàdon et al., 1996). Gray and Rickard (1981) observed substantially higher radioactivity levels in plasma than in brain over time after i.v. injection of adult rats with 14C-labeled DLM. Levels in fat were significantly higher than in other tissues. DLM and other pyrethroids are metabolized in adult male rats and mice by ester cleavage and by cytochrome P450 (P450)-catalyzed hydroxylation, with subsequent glucuronidation and sulfation (Fig. 1) (Soderlund and Casida, 1977; Ruzo et al., 1978, 1979). Pyrethroids with a cyano group (e.g., DLM) were the least prone to ester hydrolysis and P450 oxidation. Anand et al. (2005) recently reported that adult male rat hepatic P450s, notably CYP1A1 and CYP1A2, play a more important role than hepatic microsomal and plasma carboxylesterases (CaEs) in biotransformation of DLM in vitro. Relatively low intrinsic clearance by plasma CaEs indicated that detoxification in the bloodstream should be modest compared with that in the liver.
A major objective of the present study was to characterize the ontogeny of DLM metabolism in the maturing male Sprague-Dawley rat. One aim was to conduct a qualitative and quantitative in vitro comparison of DLM biotransformation in the maturing rat with that observed previously in the adult (Anand et al., 2005). A related aim was to assess the influence of maturation on in vivo blood DLM time profiles and manifestations of neurotoxicity. The in vitro metabolic rate constants and in vivo blood DLM time profiles will subsequently be utilized for construction of a physiologically based pharmacokinetic (PBPK) model to predict target organ dosimetry of DLM in immature rats and in children.
Materials and Methods
Chemicals. DLM ((S)-α-cyano-3-phenoxybenzyl-(1R, cis)-2,2-dimethyl-3-(2,2-dibromvinyl)-cyclopropanecarboxylate) was obtained from ChemService (Westchester, PA; 98% pure). Acetonitrile [high performance liquid chromatography (HPLC)-grade] was procured from Burdick and Jackson (Muskegon, MI). Tetra isopropyl pyrophosphoramide (iso-OMPA) was purchased from Sigma Chemical Co. (St. Louis, MO). All other chemicals were of analytical grade.
Animals. Timed-pregnant (gestational day 12) and adult (90-day-old) male Sprague-Dawley (SD) rats were obtained from Charles River Inc. (Raleigh, NC). The rats were housed individually and allowed water and food (Purina Rat Chow 5001; Purina, St. Louis, MO) ad libitum. The Association for Assessment and Accreditation of Laboratory Animal Care-approved animal care facility was maintained at 72 ± 2°F and 50 ± 10% humidity with a 12-h light/dark cycle. The experimental protocol was reviewed and approved by the University of Georgia Animal Care Committee. The day each litter was born was considered postnatal day 0 (PND0). On PND3, pups were sexed and a total of 8 pups were housed with each dam. The pups (largely males) were housed with dams until sacrifice on PND10 or upon weaning (PND21). The remaining pups were housed separately (5/cage) until PND40, when they were utilized. Only male rats were used in this study.
Plasma and Liver Microsome Preparation. Male rat pups of different ages (10, 21, and 40 days) were sacrificed by cervical dislocation, and blood and liver samples were collected. Blood was drawn by cardiac puncture and centrifuged at 2500g for 10 min in a Microfuge 22R centrifuge (Beckman Coulter, Atlanta, GA) to obtain plasma. The plasma was frozen at –80°C until the time of use. It was necessary to combine the livers and plasma of 12 to 14 PND10 pups (interlitter pool) and 7 to 8 PND21 pups (intralitter pool) to obtain a total of ∼ 5 g of liver and 2.5 ml of plasma. Four separate pooled liver and plasma composites per age group were obtained for analyses. The liver and plasma of each PND40 rat were adequate to yield 5 g of tissue and 2.5 ml of plasma per animal.
Liver microsomes were prepared by differential centrifugation. Each 5-g portion of fresh liver was homogenized in 25 ml of ice-cold 0.02 M Tris-KC1 buffer, pH 7.4, using a Teflon/glass tissue homogenizer. The homogenized liver was centrifuged using a Beckman Avante (JE25) centrifuge (Beckman, Palo Alto, CA) for 30 min at 4°C at 12,000g. The supernatant was collected and centrifuged using a Beckman Coulter Optima XL-100K ultracentrifuge (Beckman Coulter, Fullerton, CA) at 4°C at 105,000g for 60 min. The microsomal pellet was resuspended in Tris-KC1, pH 7.4 buffer and frozen at –80°C. Protein content of the microsomes was measured by a standard colorimetric procedure. Because the liver microsomes were resuspended in a volume of buffer the same as the original volume of liver, enzyme activities measured in each milliliter of suspension could be expressed per gram of liver (Anand et al., 2005).
In Vitro Metabolism of DLM. The kinetics of DLM metabolism in plasma and hepatic microsomes was evaluated by measuring the rate of disappearance of parent compound. Hydroxylated metabolites of DLM are not commercially available to use as standards. Initial experiments (data not shown) were conducted to verify the optimal incubation conditions for each assay (plasma CaEs, liver CaEs, and liver P450s). Disappearance of DLM was linear with respect to time and tissue concentration over the DLM concentration ranges relevant to this study. Separate incubations were performed for plasma and liver microsomal CaE- and P450-mediated metabolism. In vitro incubations of each liver or plasma sample from the 10-, 21-, and 40-day-old pups were conducted concurrently. Incubation conditions varied with the animal donors' age, as described below. The velocity (Vo) of each reaction was expressed as nmol DLM disappearance/h/g liver or ml plasma. CaE metabolism was characterized using the inhibitor iso-OMPA (1 mM in plasma and 24 mM in liver, final concentration). Details on the choice and efficacy of the CaE inhibitor (i.e., iso-OMPA) have been described in our previous report (Anand et al., 2005). NADPH-dependent metabolism was considered as P450-mediated detoxification.
Plasma Metabolism of DLM. Various concentrations (2, 10, 20, 100, and 200 μM final) of DLM were incubated at 37°C with 500 μl of Tris-HCl buffer (pH 7.4)-diluted plasma (40% for PND10 and 30% for PND21 and 40). The incubation times were 120 min for PND10 and 21 and 60 min for PND40. One set of plasma samples was preincubated with iso-OMPA for 1 h for CaE inhibition (Anand et al., 2005) before addition of DLM. The reaction was initiated by adding 500 μl of diluted ice-cold plasma (with or without iso-OMPA) to 5 μl of various DLM concentrations (in acetonitrile, never more than 1% of total volume). Similar incubations were conducted with Tris-HCl buffer, which served as a nonenzymatic control. After the respective incubation periods, 1.5 ml of acetonitrile were added for DLM extraction and analysis, as described below. The difference in DLM concentrations, between tubes with and without isoOMPA, was considered to be due to CaE-mediated metabolism.
Liver Microsomal Metabolism of DLM.Liver CaEs. Liver microsomes from PND10 (1:2 dilution, ∼10 mg protein/ml), PND21 (1:5 dilution, ∼4 mg protein/ml), and PND40 (1:10 dilution, ∼2 mg protein/ml) pups were preincubated with or without 10 μl of iso-OMPA for 1 h at room temperature. One milliliter of ice-cold microsomes with or without iso-OMPA was added to 10 μl of a series of concentrations of DLM (2, 10, 20, 100, and 200 μM final) to initiate the reaction. PND10 microsomes were incubated for 60 min, whereas PND21 and 40 microsomes were incubated for 30 min at 37°C. A buffer control was included to account for background hydrolysis. At the end of the incubation periods, 3 ml of acetonitrile were added to the incubation mixture to stop the reaction and to extract the DLM. DLM extraction and analysis are described below. The difference in DLM concentrations between tubes with and without iso-OMPA was considered to be due to CaE-mediated metabolism.
Cytochromes P450. DLM metabolism by P450s was assessed by incubating 1 ml of Tris-HCl buffer (pH 7.4)-diluted microsomes (4 mg protein/ml for PND10 and 21; 1 mg protein/ml for PND40) with 10 μl of a series of DLM concentrations (2 to 200 μM final) with and without NADPH (1.2 mM final) at 37°C. PND10 microsomes were incubated for 30 min, whereas PND21 and 40 microsomes were incubated for 15 min. The reaction was initiated by adding the ice-cold microsomes. A buffer control was also included to account for nonenzymatic hydrolysis. At the end of the incubation period, 3 ml of acetonitrile were added to the incubation mixture to terminate the reaction and to extract the DLM. DLM extraction and analysis are described below. The difference in DLM concentrations between tubes with and without NADPH was considered to be due to P450-mediated metabolism.
DLM Extraction and HPLC Assay. Addition of acetonitrile to biological samples terminated enzymatic reactions by protein denaturation and precipitation. The tubes were vortexed for 5 min and centrifuged for 10 min at 2500g (Beckman Coulter, Atlanta, GA). Two hundred microliters of supernatant were transferred to a HPLC vial, from which 40 μl were injected onto the HPLC column. Duplicate injections were made of all samples, and an average of the two injections was used for calculation of the DLM concentration.
DLM in plasma and microsomes was measured according to the HPLC method of Kim et al. (2005). The HPLC system was manufactured by Waters Inc. (Milford, MA) and consisted of a Waters 717 autosampler, a 515 pump, and a 2487 UV detector. The columns used were a Waters Spherisorb 5-μm C8 column (150 mm long × 4.6 mm internal diameter) and a Waters Spherisorb S5 C8 guard column (10 mm long × 4.6 mm internal diameter). The mobile phase was 70:30 acetonitrile/1% sulfuric acid at a flow rate of 1 ml/min. Peak heights were quantified at 230 nm by the external standard technique using standard solutions of DLM (5 ng to 10 μg).
Since we have recently measured and reported (Anand et al., 2005) metabolic rate constants for adult rats (PND90), these data were included in the present report. Nonetheless, in vitro incubations of PND90 liver microsomes and plasma were performed along with other ages with the lowest (2 μM) and highest (200 μM) DLM concentrations to determine whether there was significant day-to-day variability in experimental results. This proved not be the case; thus, the previously published PND90 data were used here to compare with immature rat data.
Assessment of Toxic Signs and Blood DLM Levels in Vivo. The objective of this phase of the study was to assess the age dependence of toxicity and of internal DLM exposure after administration of equivalent doses of DLM to young and adult rats, so that in vivo findings could be related to the in vitro metabolism data. Ten-, 21-, 40-, and 90-day-old male SD rats received 10 mg DLM/kg in glycerol formal by gavage. This dose was selected on the basis of our own preliminary studies and the LD50 data of Sheets et al. (1994). Since there are no widely accepted biochemical markers for pyrethroid neurotoxicity (Soderlund et al., 2002), our rats were monitored for commonly manifested signs: salivation, tremors, and lethality. The signs were evaluated (n = 4) periodically for up to 6 h after DLM exposure. The salivation was graded using the following scoring system: 0, no saliva; 1, wet area around the mouth and nose (mild); 2, wet area around the mouth and nose, and some on the neck (moderate); 3, entire mouth, nose, and neck covered with saliva (severe). The severity of tremors was graded as follows: 0, no tremor; 1, slight tremor or loss of balance after being picked up (mild); 2, constant tremor (moderate); 3, violent tremor and no voluntary control of movement (severe). The observer was blinded to the treatment of the rats.
Rats (n = 3 or 4) of each age were serially sacrificed from 15 min to 24 h postdosing to collect blood and selected tissues for analyses of DLM. Because of the 100% mortality, it was possible to collect samples only up to 6 and 12 h for PND10 and 21 animals, respectively. The blood samples were processed immediately for DLM analysis as described by Kim et al. (2005). Only blood DLM profiles and toxicity data are presented here. The tissue DLM profiles will be reported elsewhere.
Data Analyses. Microsomal and plasma incubations were conducted with four pooled or individual samples for each age group. The Michaelis-Menten metabolic rate constants (Vmax and Km) were calculated by nonlinear regression using Prism (3.03) (GraphPad Software, Inc., San Diego, CA). Regression coefficients (r2) to corroborate age-dependent internal DLM exposure to toxic signs were also computed using the same software. Areas under blood DLM concentration versus time curves (AUCs) were calculated by the trapezoidal rule. The metabolic rate constants (Figs. 2, 3, 4, 5 and Table 1) were initially analyzed by a global, repeated ANOVA using Km, Vmax, and intrinsic clearance as the dependent variables, and age and enzyme (i.e., plasma CaE, liver CaE, or liver P450) as the independent variables. Where significant interactions were noted between the dependent variables, step-down ANOVAs were conducted on each dependent variable against age and enzyme activity. Based on the interactions, further step-down ANOVAs were conducted with subsequent comparisons between different ages or different enzymes made using a Tukey-Kramer post hoc test. All analyses were conducted using StatView (SAS, Cary, NC; version 5.0.1). Statistical significance at all levels of analysis was considered to be p ≤ 0.05. All data are presented as mean ± S.E.
Age-dependent in vitro metabolic rate constants for DLM metabolism
Results are expressed as mean ± S.E. (n = 4). Vmax values are expressed as nmol/h/g liver or ml plasma; Km values are expressed as μM; intrinsic clearance (Vmax/Km) values are expressed as h-1. Statistical comparison (p ≤ 0.05) was made between different ages for each parameter for each enzymatic pathway. a Significantly different from PND10, b significantly different from PND21, and c significantly different from PND40. Anand et al. (2005) is the source of the PND90 values. Reprinted from Toxicol Appl Pharmacol Anand SS, Bruckner JV, Haines WT, Muralidhara S, Fisher JW, and Padilla S Characterization of deltamethrin metabolism by rat plasma and liver microsomes. In press, 2005 with permission from Elsevier.

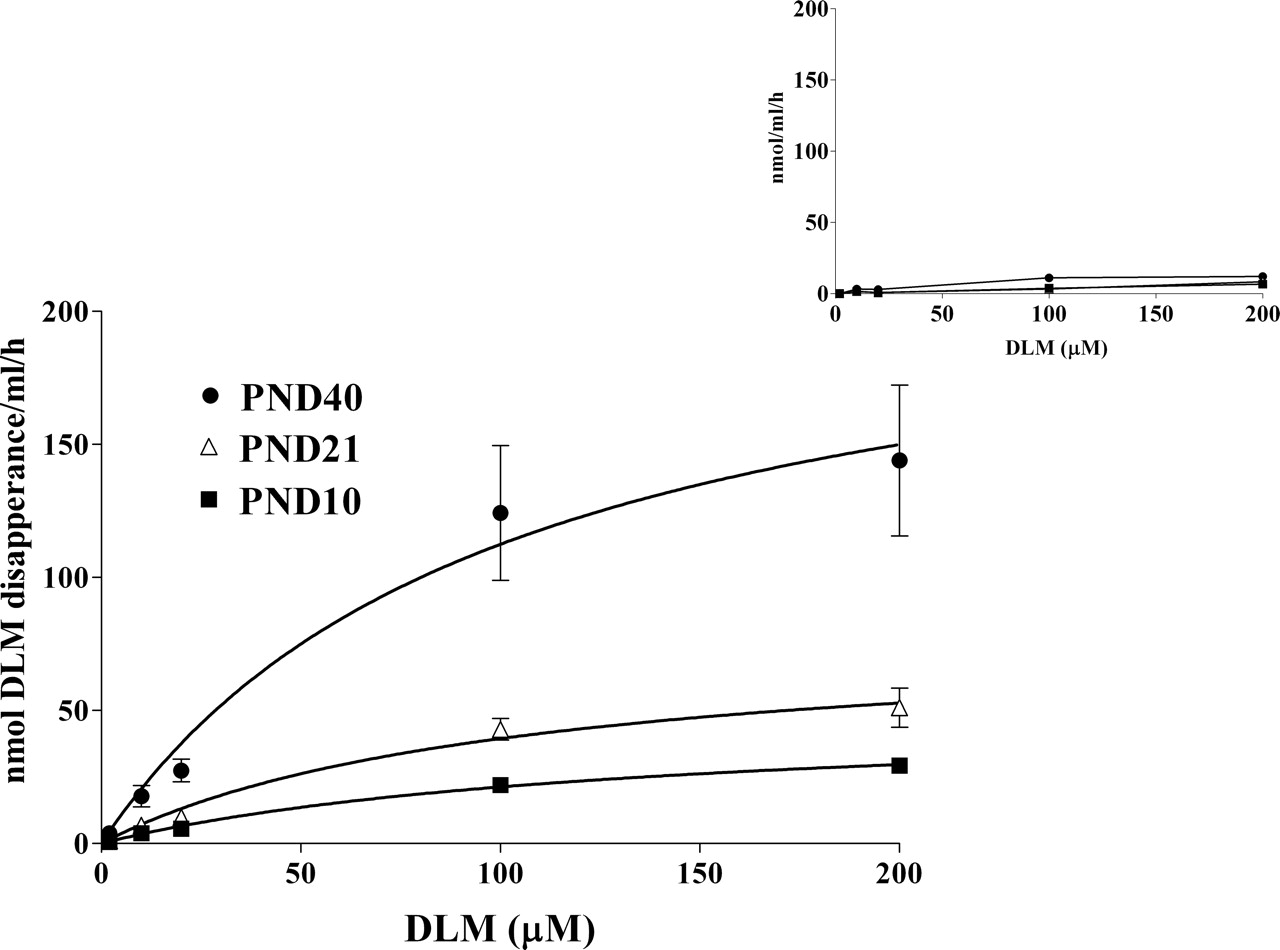
Age-related DLM metabolism in rat plasma. Various concentrations of DLM (2, 10, 20, 100, or 200 μM) were incubated with 500 μl of diluted plasma (PND10, 40% plasma; PND21 and 40, 30% plasma) at 37°C for 120 (PND10 and 21) or 60 (PND40) min. Values represent mean ± S.E. for four plasma samples (pooled or individual). The inset presents DLM metabolism after preincubation of diluted plasma with iso-OMPA for 1 h. Michaelis-Menten rate constants calculated by nonlinear regression are presented in Table 1.

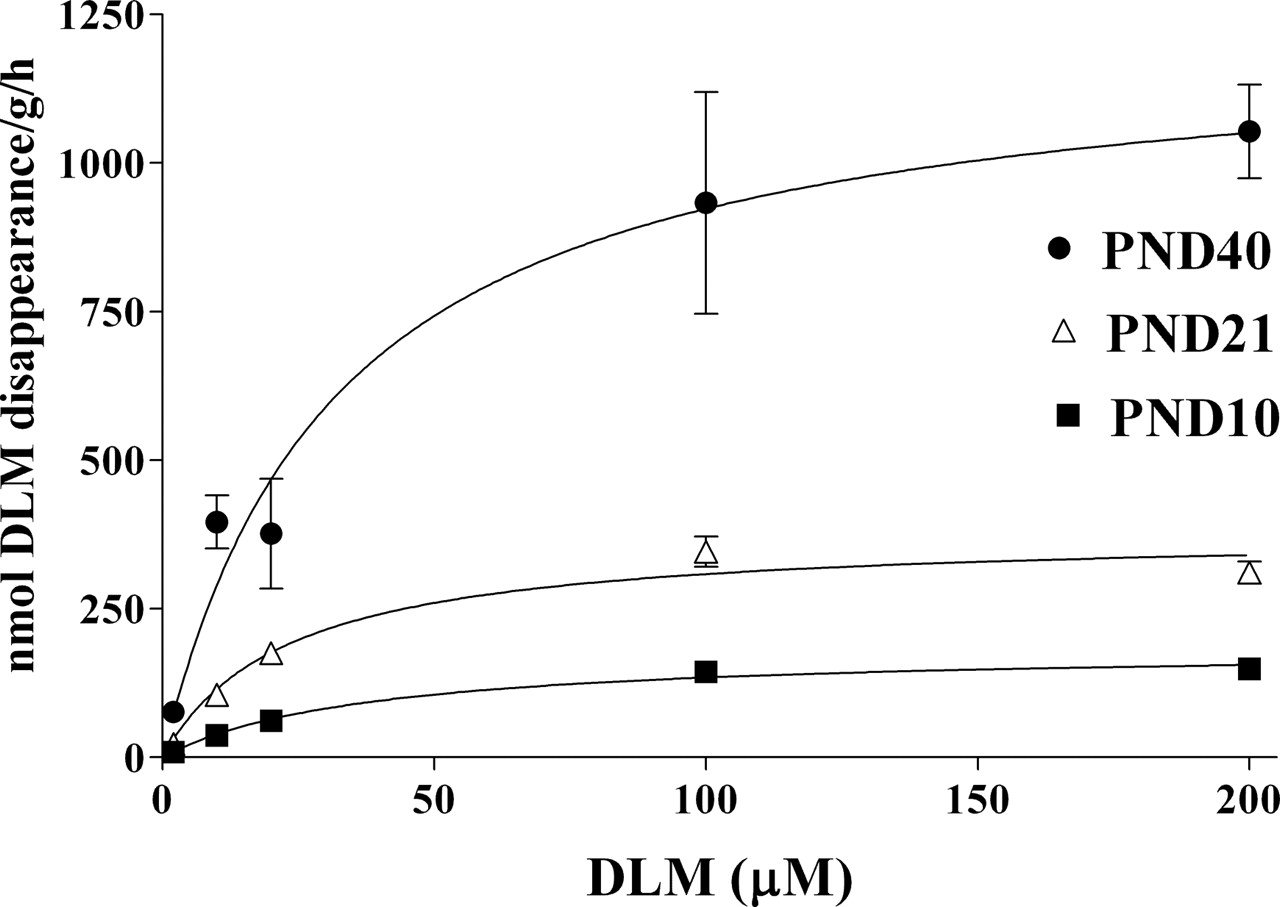
Age-dependent DLM metabolism by liver microsomal CaEs. A series of concentrations of DLM (2, 10, 20, 100, and 200 μM) was incubated with diluted liver microsomes for 30 (PND21 and 40) and 60 (PND10) min at 37°C. Values represent mean ± S.E. for four hepatic microsomal preparations (pooled or individual liver samples). The inset presents DLM metabolism after preincubation of liver microsomes with iso-OMPA for 1 h. Michaelis-Menten rate constants calculated by nonlinear regression are presented in Table 1.

Age-dependent DLM metabolism by liver microsomal P450s of 10-, 21-, and 40-day-old rats. Two, 10, 20, 100, or 200 μM DLM were incubated with 1 ml of liver microsmes (PND10 and 21, 4 mg protein/ml; PND40, 1 mg protein/ml) with or without NADPH (1.2 mM) for 30 (PND10) or 15 (PND21 and 40) min at 37°C. Values present mean ± S.E. for four different microsomal samples (pooled or individual liver samples). Michaelis-Menten rate constants calculated by nonlinear regression are presented in Table 1.
Results
Plasma Metabolism by CaEs. The age dependence of DLM biotransformation by plasma CaEs in rats is illustrated in Fig. 2. The rate of DLM metabolism increases with the age, as reflected by the progressive increases in Vmax. The calculated Vmax and Km values are presented in Table 1. Vmax values of 90-day-old adults are ∼6.8 and 3.7 times higher (p ≤ 0.05) than for PND10 and 21 pups, respectively. The PND10 and 21 Vmax values are not statistically different, nor are the PND40 and 90 Vmax values. Km values remain relatively constant from PND10 to 40. The mean PND90 Km appeared to be somewhat higher, but the elevation was not statistically significant. Intrinsic clearance increases significantly with age, reaching adult levels by PND40. Addition of iso-OMPA almost completely inhibited DLM metabolism in each age group (Fig. 2 inset), confirming the role of CaEs in DLM biotransformation in plasma of immature rats.
Hepatic Microsomal Metabolism by CaEs. DLM metabolism by liver microsomal CaEs (Fig. 3) is age-dependent, but its developmental pattern differs in some respects from that of plasma CaEs. The male rat liver CaE metabolic capacity increases significantly with age (Table 1). There is roughly a 5-fold increase (p ≤ 0.05) in liver CaE activity between PND10 and 21, in contrast to the lack of significant rise in plasma CaE activity during this period. The liver microsomal Vmax for PND40 rats is significantly higher than for PND10 (∼12-fold) and 21 rats (∼2.6-fold). Nevertheless, the PND40 Vmax is only 15% (p ≤ 0.05) of that in 90-day-old adults (Table 1). Km values remain relatively constant during maturation. Intrinsic clearance in the PND90 adults was significantly higher than in the immature age groups. The PND21 and 40 clearances were not significantly different from one another, but were higher than in PND10 pups. The addition of iso-OMPA completely inhibited DLM metabolism in each immature age group at all the concentrations studied (Fig. 3 inset).
Hepatic Microsomal Metabolism by P450s.Figure 4 presents age-dependent metabolism of DLM by P450s. The maximal velocity of DLM detoxification by liver P450s doubles between PND10 and 21, and triples again by PND40 (Table 1). The net result is almost a 7-fold increase in metabolic capacity between PND10 and 40. As with plasma and liver microsomal CaEs, affinity of P450 for DLM does not vary with the age of the immature rats, but the Km of adult rats is significantly higher than those of the young rats. The Vmax and Km of the PND40 rats are each only about one-half those of the adults; thus, intrinsic clearance does not change between 40 and 90 days (Table 1). Statistically significant increase in intrinsic clearance is noted with increasing age until PND40.

Age-dependent changes in the intrinsic clearance (Vmax/Km) of DLM by liver microsomal and plasma CaEs and liver microsomal P450s (A). Statistical comparison (p ≤ 0.05) was made between enzymatic pathways for each age. a, significantly different from plasma CaE; b, significantly different from liver CaE. Intrinsic clearance is expressed as percentage of adult values in B.
Findings of the age dependence of plasma and liver microsomal CaE-catalyzed and liver microsomal P450-mediated intrinsic clearance of DLM are presented graphically in Fig. 5. The efficiency of liver microsomal and plasma CaEs in metabolizing DLM is comparable in the 10-, 21-, and 40-day-old rats (Fig. 5A). In contrast, Anand et al. (2005) found that liver microsomal CaEs of adult male SD rats were more efficient (p ≤ 0.05) than plasma CaEs in biotransformation of DLM, as is evident in Fig. 5A. It is worthy of note, again, that liver CaE activity of PND40 pups is only about 15% of that in PND90 adults (Fig. 5B). P450-catalyzed DLM metabolism is substantially greater than metabolism by plasma and/or hepatic microsomal CaEs at each developmental stage (Fig. 5A). Liver P450 and plasma CaE activities increase to approximately adult levels between days 10 and 40 in the maturing male SD rats (Fig. 5B). The relatively modest increase in liver microsomal CaE activity during this developmental period appears to be primarily responsible for the 40-day-old rats' inability to attain adult DLM metabolic capacity (Fig. 5B).
DLM Internal Exposure and Neurotoxicity after in Vivo Dosing. Blood DLM concentration versus time profiles are plotted in Fig. 6A. DLM is rapidly absorbed from the GI tract. Blood levels appear to peak around 1 h after the oral dosing in the 40- and 90-day-old rats, but plateau in the 10- and 21-day-old rats. Pronounced toxicity likely contributed to the unusual kinetic profiles in the latter two groups. AUCs are truncated at 6 and 12 h, because of the death of each PND10 and 21 animal, respectively, soon after these times (Fig. 6B). In each instance, the AUCs vary inversely with age. Age-dependent decreases in blood AUCs were statistically significant except the decrease noted between PND40 and 90. The Cmax values also appeared to progressively decrease with increasing age, although the PND21, 40, and 90 values were not statistically different from one another (Fig. 6C).

Age-related changes in the blood DLM concentration versus time profiles in rats receiving 10 mg DLM/kg p.o. (A). Treatment details are provided under Materials and Methods. Results represent mean ± S.E. (n = 3 or 4). Mean blood DLM AUC0–6 h and AUC0–12 h values are presented in B. Statistical comparison (p ≤ 0.05) was made between ages. a, significantly different from 10 days; b, significantly different from 21 days. C shows mean ± S.E. (n = 3 or 4) Cmax values. * indicates statistically significant difference from PND10 group (p ≤ 0.05).
The severity of signs of DLM neurotoxicity was inversely related to age (Table 2). It is evident that the most pronounced salivation and tremors were experienced by the youngest rats. Both manifestations of neurotoxicity increased over time after dosing in the PND10 and 21 animals. This was particularly true for tremors. All of the PND10 and 21 rats died between 6 and 8 h and 12 and 16 h, respectively. None of the PND40 or 90 rats succumbed. No tremors were observed in the members of either older group. Salivation was mild (PND90) to moderate (PND40) and lasted only 2 to 3 h.
Age-dependent toxic signs following DLM administration
Male SD rats were given 10 mg DLM/kg by gavage. Whereas 10- and 21-day-old rats showed salivation (S) and tremors (T), 40- and 90-day-old rats experienced only transient salivation. Sustained toxic signs resulted in 100% mortality between 6 and 8 h in 10-day rats and between 12 and 16 h in 21-day rats. All the 40- and 90-day rats survived. The severity of the signs was assessed using subjective scoring as follows: 0, no appearance of signs; 1, mild; 2, moderate; and 3, severe. The scale is defined under Materials and Methods. Mean values of four rats are presented.
There was concordance between the severity of acute neurotoxic signs and the magnitude of systemic DLM exposure. Total scores were obtained for salivation and for tremors by summing the 1-, 2-, 3-, 4-, and 6-h scores in Table 2 for each age group. These total scores for each toxic sign were plotted against the groups' blood DLM AUCs (0–6 h for PND10 and 0–12 h for PND21, 40, and 90 rats) (Fig. 7). There was a good correlation between AUCs and severity of salivation (r2 –0.87) and between AUCs and severity of tremors (r2 –0.90). Lower correlation coefficients were obtained when relating the total toxicity scores to Cmax values (r2 –0.63 for salivation versus Cmax; r2 –0.70 for tremors versus Cmax) (data not shown).
Discussion
An important aim of this study was to test the hypothesis that inefficient metabolic detoxification is a contributing factor to the greater susceptibility of neonatal and weanling rats to acute DLM neurotoxicity. Sheets et al. (1994) clearly showed that DLM toxicity varied with age of immature Long-Evans rats. In the present study, 10- and 21-day-old male SD rats gavaged with 10 mg DLM/kg exhibited pronounced salivation and tremors leading to 100% mortality between 6 and 8 h and 12 and 16 h in the PND10 and 21 groups, respectively. PND40 and 90 rats receiving the same dose experienced only transient salivation, and all the rats survived. Decreased internal exposure to DLM during maturation, as evidenced by AUCs and Cmax values (Fig. 6, B and C) paralleled the severity of poisoning (Table 2). AUC (Fig. 7) showed a higher degree of correlation with toxic signs than did Cmax (data not shown). DLM metabolism in vitro by plasma and liver CaEs and hepatic P450s exhibited progressive increases with maturation (Table 1). These findings strongly support the aforementioned hypothesis.
There has been a paucity of recent data on the metabolism and systemic disposition of DLM and other pyrethroids, despite their rapidly expanding use in the United States and other countries. This is particularly true of immature laboratory animals and humans. Anand et al. (2005) identified and clarified the relative contributions of liver P450s and liver and plasma CaEs to DLM metabolism in adult, male SD rats. The current study is the first to characterize the ontogeny of DLM metabolism. The age-dependent increases in the capacity of plasma CaEs and P450s to detoxify DLM reach adult levels by 40 days, but the contribution of liver CaEs is more limited in the PND40 rats than in adults (Fig. 5). These data are in agreement with the age-dependent reductions in blood DLM levels (Fig. 6) and toxic signs (Table 2). The findings should be generally applicable to other pyrethroids, although the extent to which these compounds are hydrolyzed and oxidized depends upon their isomerization and substitutions (Soderlund and Casida, 1977). Age-, sex-, or species-dependent differences may assume greater importance in the detoxification of pyrethroids that are designed to be metabolically stable than would be the case with very rapidly metabolized congeners.

Correlation of internal exposure to DLM (blood AUCs) with salivation and tremors. The total score was calculated by summing the 1-, 2-, 3-, 4-, and 6-h scores of each toxic sign for each age group. Correlation coefficient (r2) between AUCs and severity of salivation is 0.87 and between AUCs and severity of tremors is 0.90.
Plasma and hepatic microsomal CaEs that cleave DLM exhibit different developmental profiles. Intrinsic clearance of DLM by plasma CaEs nearly matches adult levels by PND40. Intrinsic clearance by liver CaEs at PND40 is only about 15% of that in adult rats (Fig. 5), suggesting that liver CaEs undergo substantial maturational changes between PND40 and 90. Investigations of the heightened sensitivity of young rats to acute organophosphate toxicity also reveal age-related increases in the activities of plasma and liver CaEs, but most of these developmental data are limited to preweanling rats (Moser et al., 1998; Karanth and Pope, 2000). Sterri et al. (1985) reported that plasma CaE activity in male Wistar rats reaches adult levels by PND30, but that liver CaE activity is still only 70 to 80% of that in adults. In contrast, Morgan et al. (1994) reported that liver CaE activity reached adult levels by 42 days, but these researchers evaluated just two isozymes (hydrolases A and B). The relationship of these isozymes to the metabolism of DLM is not known. The significantly lower metabolic capacity of PND40 than of PND90 liver CaEs (Fig. 5) apparently reflects the relatively slow development of hepatic CaEs that cleave DLM.
There are apparently qualitative, as well as marked quantitative, age-related differences in both rat plasma and liver CaEs that metabolize DLM (Table 1). There is relatively little variation in plasma or liver Km values during this developmental period. Bell and Ecobichon (1975) used α-naphthyl acetate as a substrate to monitor CaE activity in the S9 liver supernatant of unsexed Wistar rats, ranging in age from 3 days prepartum to PND35. These researchers report a progressive decrease in Km from prepartum day 3 to PND 14, followed by a progressive rise through PND 35. Adult Km values were not reported. We, like Bell and Ecobichon (1975), observed marked increases in Vmax values throughout maturation. The substantial increases in intrinsic clearance of DLM were thus largely quantitative in nature. CaEs are group of substrate-specific isozymes that are initially synthesized as precursors, which undergo structural modifications to yield the mature forms (Satoh and Hosokawa, 1998). Maturational changes associated with different isoforms and structural modifications may contribute to the modest qualitative and marked quantitative changes, as well as the differences in the developmental profile we observed in rat plasma and liver CaEs that metabolize DLM.
DLM metabolism in vitro by P450 is substantially greater than that by plasma or liver CaEs in maturing male SD rats (Table 1), as is the case in adults (Anand et al., 2005). This finding is supported by the results of an in vivo toxicity and metabolism study by Ruzo et al. (1979). These investigators found that pretreatment of mice with tributyl phosphorothionate, an esterase inhibitor, decreased urinary DLM metabolite excretion and also decreased the LD50 by 2-fold. Pretreatment of mice with piperonyl butoxide, a P450 inhibitor, inhibited metabolite excretion and produced a 3- to 5-fold decrease in LD50. Age-dependent increase in the intrinsic clearance by P450 in the current study was mainly due to increases in Vmax, not Km. Intrinsic clearance by P450 reached adult levels by PND40, but adult Vmax and Km values were still significantly higher than PND40 values. The differences between 40 and 90 days may be associated with maturational changes in the P450s associated with DLM biotransformation.
The identity of the P450 enzyme(s) that metabolizes DLM has not been thoroughly investigated, although CYP1A1/2 and CYP2B1/2 are implicated. Using commercially available rat P450 enzymes, Anand et al. (2005) found that CYP1A1 and CYP1A2 were quite effective, and CYP2C11 exhibited modest activity in transforming DLM in vitro. Dayal et al. (1999) reported induction of CYP1A1/2 and CYP2B1/2 activities in liver after repeated (7-, 15-, or 21-day) exposures to DLM. Whereas reports on developmental changes (Gebremichael et al., 1995; Atterberry et al., 1997) of CYP1A1/2 and CYP2B1/2 activities are contradictory, ontogeny of DLM metabolism (Table 1) is not in accordance with the developmental changes of these enzymes. The possible reasons could be that 1) ethoxyresorufin O-deethylase and pentoxyresorufin O-dealkylase are usually considered to be catalyzed by CYP1A1/2 and CYP2B1/2, respectively, but other P450s (i.e., CYP2C6 and 2C11) also contribute to deethylase and dealkylase activities (Kobayashi et al., 2002); and 2) there is no consensus on constitutive protein expression of CYP1A1/2 and CYP2B1/2 in rats (de Waziers et al., 1990; Agrawal and Shapiro, 2003).
We did recently find that among the commercially available human P450 enzymes, CYP1A1, 1A2, and 2B6 were capable of metabolizing DLM (unpublished data). This suggests that the same enzymes could metabolize DLM in rats and humans. Constitutive CYP1A1 expression is often undetectable in adult human liver, and the developmental profile of CYP2B6 is largely unknown (Hines and McCarver, 2002). Sonnier and Cresteil (1998) demonstrated that CYP1A2 expression was absent in the liver of human neonates, gradually increased in infants, but still only one-half that in adults. Although human CaEs that metabolize DLM have not yet been identified, a limited study of the maturational expression of human liver CaEs showed that activity in five infants <2 years old was not significantly lower than that in five adults (Pope et al., 2005). There may well be qualitative and quantitative interspecies differences in enzymes involved in DLM biotransformation. Nevertheless, the general deficit in CaEs and P450s in the more immature humans and rodents suggests that these age groups of both species will be particularly susceptible to acute DLM neurotoxicity.
In summary, the age-dependent increase in the DLM metabolism in vitro is consistent with the age-dependent decreases in the blood DLM levels and toxicity in vivo. Nevertheless, it should be recognized that age-dependent pharmacokinetic factors other than metabolism may also affect the bioavailability of DLM and the magnitude of its adverse effects. Preweanling rats are shown to have relatively high GI absorption rates (Hoffman, 1982; Naylor and Harrison, 1995). P-glycoprotein, which may play a role as an efflux transporter of pyrethroids (Bain and LeBlanc, 1996; Buss et al., 2002) has been found to be deficient in young mice (Mahmood et al., 2001). Extensive absorption and/or decreased efflux from the GI tract of young animals could contribute to the higher internal exposure and toxicity of DLM. Adult rats have larger amounts of body fat than perinatal animals. This may contribute to greater deposition of the highly lipophilic compound in the young animal's brain, as could an ineffectual blood-brain barrier. Srinivas et al. (2004) report that P-glycoprotein expression, relatively high levels of esterases and phosphatases, and pharmacodynamic effects are all responsible for resistance of moth larvae, a relatively simple organism, to pyrethroids. Investigations of the potential role of such age-dependent processes in appropriate mammalian species should improve our understanding of their relative importance, as well as the applicability of different species to children's risk assessments. The metabolic parameters determined here are being used in construction of a PBPK model for DLM in immature rats. Other key input parameters can be incorporated as they are identified. PBPK models, in turn, can be useful in informing the investigator of processes or parameters that deserve attention.
Acknowledgments
We thank Dr. Kevin Crofton and Dr. Mike DeVito for critical review of an earlier version of the manuscript. We also thank Dr. Catherine White for input in the in vivo experiments.
Footnotes
-
This project was supported by EPA STAR Grant R830800. This research has been reviewed by the National Health and Environmental Effects Research Laboratory, U.S. Environmental Protection Agency, and approved for publication. Approval does not signify that the contents necessarily reflect the views and policies of the Agency, nor does mention of trade names or commercial products constitute endorsement or recommendation for use.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.007807.
-
ABBREVIATIONS: DLM, deltamethrin; GI, gastrointestinal; P450, cytochrome P450; AUC, area under the curve; CaE, carboxylesterase; HPLC, high performance liquid chromatography; PBPK, physiologically based pharmacokinetic; iso-OMPA, tetra isopropyl pyrophosphoramide; SD, Sprague-Dawley; PND, postnatal day; AUC, area under the concentration versus time curve; ANOVA, analysis of variance.
- Received October 10, 2005.
- Accepted November 30, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}