


Abstract
Isolated primary human hepatocytes are a well accepted system for evaluating pharmacological and toxicological effects in humans. However, questions remain regarding how culturing affects the liver-specific functions of the hepatocytes. In addition, cryopreservation could also potentially affect the differentiation state of the hepatocytes. The first aim of the present study was to compare gene expression in freshly isolated primary hepatocytes to that of the liver of origin and to evaluate the expression changes occurring after cryopreservation/thawing, both when maintained in suspension and after plating. The second aim of the present study was to evaluate gene expression in hepatocytes after cold storage of suspensions up to 24 h compared with freshly isolated hepatocytes in suspension. Our results show that the gene expression in freshly isolated human hepatocytes in suspension after isolation is similar to that of the liver of origin. Furthermore, gene expression in primary human hepatocytes in suspension is not affected by hepatocyte cold storage and cryopreservation. However, the gene expression is profoundly affected in monolayer cultures after plating. Specifically, gene expression changes were observed in cultured relative to suspensions of human hepatocytes that are involved in cellular processes such as phase I/II metabolism, basolateral and canalicular transport systems, fatty acid and lipid metabolism, apoptosis, and proteasomal protein recycling. An oxidative stress response may be partially involved in these changes in gene expression. Taken together, these results may aid in the interpretation of data collected from human hepatocyte experiments and suggest additional utility for cold storage and cryopreservation of hepatocytes.
The liver serves as the primary site of detoxification of exogenous and endogenous compounds in the systemic circulation. Other biological and physiological functions include the production and secretion of critical blood and bile components, such as albumin, bile salts, and cholesterol. The liver is also involved in protein, steroid, and fat metabolism, as well as vitamin, iron, and sugar storage.
One of the most complex functions specific to liver is its ability to metabolize an enormous range of xenobiotics. Many drugs present in the blood are absorbed by hepatocytes, where they can be metabolized via phase I and II biotransformation reactions. Information concerning hepatic drug uptake and metabolism, phase I and phase II induction, drug interactions affecting hepatic metabolism, and hepatotoxicity are essential for the pharmacology and toxicology of a given drug. Because of major species differences both in the catalytic activities and regulation of enzymes involved in drug metabolism, many of these evaluations can only be accurately investigated with human tissue. Because intact cells more closely reflect the environment to which drugs are exposed in the liver, isolated primary human hepatocytes are a well accepted system for evaluating pharmacological and toxicological effects in humans (Gomez-Lechon et al., 2004; O'Brien et al., 2004; Liguori et al., 2005) and have been described as one of the best in vitro models for the prediction of in vivo metabolism in humans (Lave et al., 1999; Li, 2001). This has prompted research groups, including ours, to optimize the isolation and culture conditions of human hepatocytes from both surgical liver resections and nontransplantable livers (Richert et al., 2004; Donato et al., 2005; LeCluyse et al., 2005).
Sparse availability of high quality human liver tissue for research purposes, the demand for standardized cell populations, and the need for proper storage of cells for future research investigations have resulted in the development of cryopreservation techniques (Alexandre et al., 2002; Garcia et al., 2003; Roymans et al., 2005). However, several problems exist with the use of thawed human hepatocytes. For instance, cryopreserved hepatocytes have displayed a substantial variability between donors both in terms of recovery of viable hepatocytes and in their capacity to attach after thawing (Alexandre et al., 2002; Blanchard et al., 2005). This has profound consequences in terms of hepatocyte differentiation rates in culture and responses to xenobiotics because it has been found that isolated hepatocytes in culture display markedly different gene expression patterns depending on attachment status (Waring et al., 2003). In addition, it has previously been shown that constitutive expression and inducibility of the cytochrome P450s (P450s) are dependent on cell density (Hamilton et al., 2001). In addition, studies have shown that cultured thawed hepatocytes in monolayers appear smaller than their freshly isolated hepatocyte counterparts in monolayers, suggesting a variability in the resistance to cryopreservation of the various liver hepatocyte populations (Alexandre et al., 2002).

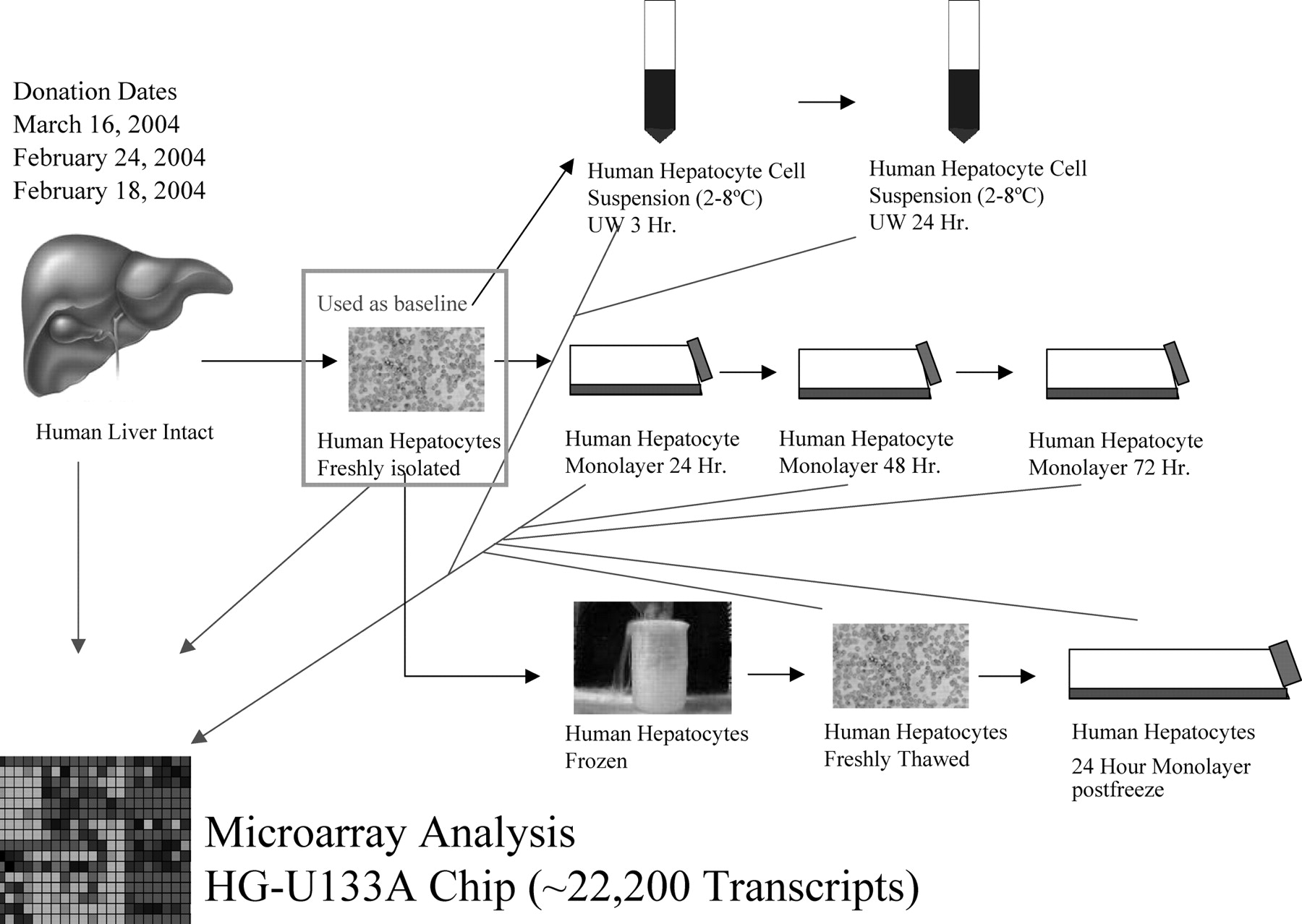
Study design is illustrated. Three distinct human liver donors were evaluated using gene expression analysis along multiple steps of the human hepatocyte preparations. Gene expression changes were monitored using freshly isolated primary human hepatocytes (t = 0 h) as the baseline.
The first aim of the present study was thus to compare gene expression in freshly isolated hepatocytes to the liver of origin and to evaluate the changes occurring after cryopreservation/thawing, both when maintained in suspension and after plating. The second aim of the present study was to evaluate gene expression in hepatocytes after cold storage in suspensions up to 24 h compared with freshly isolated hepatocytes in suspension. A pictorial depiction of the intended study is represented in Fig. 1.
Materials and Methods
Materials. Bovine serum albumin, phosphate-buffered saline, fetal calf serum, Dulbecco's modified Eagle's medium (DMEM), and TRIzol were purchased from Invitrogen-Life Technologies (Cergy-Pontoise, France). Collagenase type IV, Percoll, and EDTA were purchased from Sigma (St. Louis, MO).
Human Hepatocyte Isolation. Adult normal liver samples were obtained from three patients undergoing partial hepatectomy for primary or secondary tumors. All experimental procedures were done in compliance with French laws and regulations and were approved by the National Ethics Committee. Table 1 depicts the patients' demographics. Liver resections were handled, and a small portion of liver was suspended in RNALater (In Vitrogen-Life Technologies) and frozen at –80°C. Hepatocytes were isolated by a two-step collagenase perfusion as recently described (Richert et al., 2004). After Percoll purification, cell viability was estimated by trypan blue exclusion. Part of freshly isolated human hepatocytes was kept directly in TRIzol (0.5 ml of TRIzol for 1 million cells) and frozen at –80°C.
Characteristics of donors and yield of viable isolated hepatocytes
Human Hepatocyte Cold Storage or Cryopreservation and Thawing. Part of the freshly isolated human hepatocytes was kept in UW solution at 4°C for 3 or 24 h. Another portion of the freshly isolated human hepatocytes was immediately subjected to cryopreservation as described previously (Alexandre et al., 2002).
Human hepatocytes after cold storage (3- and 24-h time points) were resuspended in DMEM and washed one time before being subjected to Percoll purification. The cell pellet obtained was suspended in TRIzol (0.5 ml of TRIzol for 1 million cells) and frozen at –80°C (see Fig. 1).
Human Hepatocyte Culture. Human hepatocytes, either directly after isolation or after cryopreservation/thawing, were subjected to Percoll purification and suspended in DMEM supplemented with fetal calf serum (5%), insulin (4 mg/l), hydrocortisone (10–6 mM), and gentamycin (50 mg/l). Some of the thawed hepatocyte cells were directly suspended in TRIzol and frozen at –80°C. Freshly isolated hepatocytes and the remaining thawed hepatocytes were plated onto 60-mm dishes at a density of 3.5 × 106 cells/dish. At each time point of culture (24, 48, and 72 h for the fresh hepatocytes and only 24 h for the thawed hepatocytes), hepatocyte cells were scraped in TRIzol (0.5 ml of TRIzol for 1 million cells) and frozen at –80°C (see Fig. 1).
RNA Extraction and Microarray Analysis. Total RNA was isolated from the TRIzol extracts using the procedure from Invitrogen. Optical density at 260 nm determined RNA concentration. RNA quality was accessed using an Agilent Technologies bioanalyzer (Agilent Technologies, Palo Alto, CA) before proceeding to microarray sample preparation.
Microarray analysis was performed using the standard protocol provided by Affymetrix Inc. (Santa Clara, CA) and, as described previously, starting with 5 μg of total RNA (Liguori et al., 2005). Fragmented, labeled cRNA was hybridized to an Affymetrix human genome U133A array, which contains sequences corresponding to roughly 22,200 transcripts at 45°C overnight.
Microarray Data Analysis. The microarray scanned image and intensity files were imported in Rosetta Resolver gene expression analysis software version 5.0 (Rosetta Inpharmatics, Kirkland, WA). Resolver's Affymetrix error model was applied, and ratios were built for each step in the study using freshly isolated human hepatocytes as the baseline. A p value is calculated for every -fold change using the Rosetta Resolver error model. For all of the gene changes shown, the p value is displayed in the figure legend. If the p value is greater than the listed value, the -fold change is shown as 1.0 (or black on a heat map). Gene ontologies were deciphered using NETAFFX or GenMAPP (Dahlquist et al., 2002; Liu et al., 2003).

Hepatocytes in suspension after isolation from donor 2 (A), in primary culture 24 h after seeding (B), 48 h after seeding (C), 72 h after seeding (D), and in primary culture 24 h after thawing (E). All panels are at 40× magnification.
Results
Study Design. Gene expression analysis was conducted for three distinct human donors according to the study design as revealed in Fig. 1. Briefly, microarray analysis was performed on human liver and freshly isolated primary human hepatocytes as well as hepatocyte monolayers from 24 to 72 h. In addition, a portion of the primary hepatocytes was cryopreserved, and these cells underwent genomic analysis immediately after thaw and again after culture for 24 h. Some of the hepatocytes were stored in a cell suspension of UW solution, and subsequent gene expression analysis was conducted at the 3- and 24-h time points. For data analysis purposes, the freshly isolated primary human hepatocytes were considered a control and were used as a baseline for ratio builds.
Primary Human Hepatocyte Morphology, Viability, and Attachment.Figure 2 displays light micrographs of different stages of the study for hepatocytes from donor 2. Table 2 exhibits the percent recovery of viable primary human hepatocytes after cold storage and cryopreservation. For all donors, viability declined upon continued cold preservation in UW solution up to 24 h, the loss of cells being more evident for donor 1 than for donor 2 and donor 3. In addition, viability was decreased upon cryopreservation (liquid nitrogen), with recovery being equivalent for donor 2 and donor 3, approximately 50%, higher than donor 1 (10%). Table 3 illustrates that primary human hepatocyte attachment efficiency declined after cryopreservation. Again, attachment rate of plated viable thawed hepatocytes was lower for donor 1 than for donors 2 and 3. As also seen in Table 3, when considering total cell protein level of attached thawed hepatocytes, the values were very similar for the three donors, being 0.85, 0.99, and 0.83 mg of protein/dish, respectively. After thawing, this total cell protein level was lower: 53% (0.45 mg/dish; donor 1), 60% (0.59 mg/dish; donor 2), and 82% (0.68 mg/dish; donor 3), respectively, than that of the respective freshly isolated attached total protein level.
Recovery of viable hepatocytes after cold preservation and cryopreservation/thawing
Hepatocyte viability was assessed by trypan blue exclusion before and after cold storage or cryopreservation. From this, the recovery of viable cells was calculated and expressed as a percentage of viable cells before preservation.
Extent of attachment, 24 h after seeding, of hepatocytes freshly isolated and after cryopreservation/thawing
Hepatocytes were plated at a density of 3.5 × 106 viable cells/dish (see Materials and Methods). After a 24-h attachment period, monolayers were washed, and cells were detached with trypsin and counted. From this, the extent of attachment of viable seeded hepatocytes was calculated. In addition, the protein content of the cell monolayers was determined.
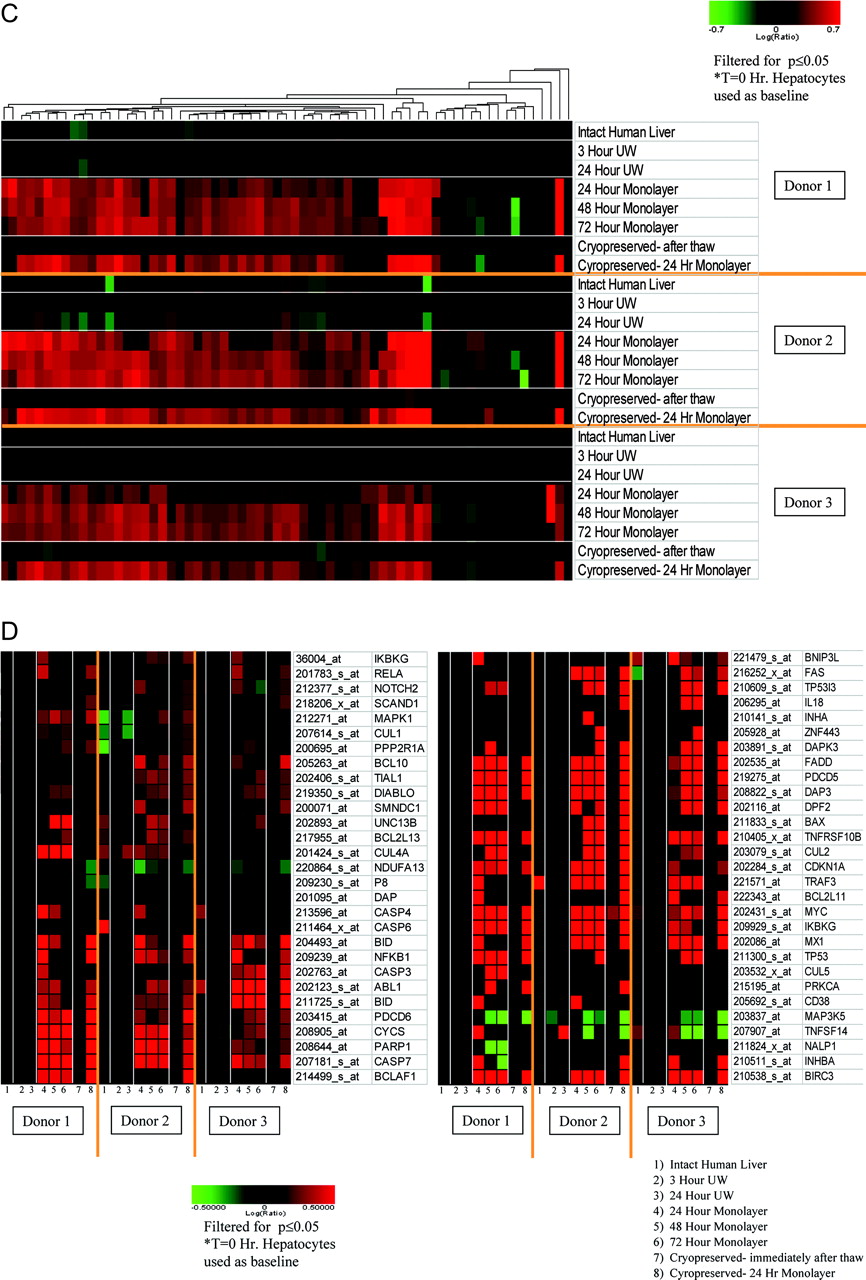
Gene Expression Analysis. Using freshly isolated primary human hepatocytes as the baseline, microarray analysis on human liver and primary human hepatocytes at various stages of preparation revealed relatively few gene expression changes for intact human liver, hepatocytes from 3- and 24-h cold storage, and thawed primary hepatocytes postcryopreservation (Fig. 3). However, the gene expression profiles from primary human hepatocytes after plating for 24 to 72 h both from fresh and cryopreserved preparations resulted in a markedly increased number of differentially regulated transcripts relative to freshly isolated primary hepatocytes. The gene expression changes in plated hepatocytes relative to freshly isolated hepatocytes were, in general, consistent over time for all donors (Fig. 3). A supplemental Microsoft Excel table is included with this communication detailing all gene expression changes (relative to freshly isolated human hepatocytes) with at least ±1.5-fold change and p value of 0.05 or less. The -fold changes for probe sets with p values greater than 0.05 are represented as “1”.
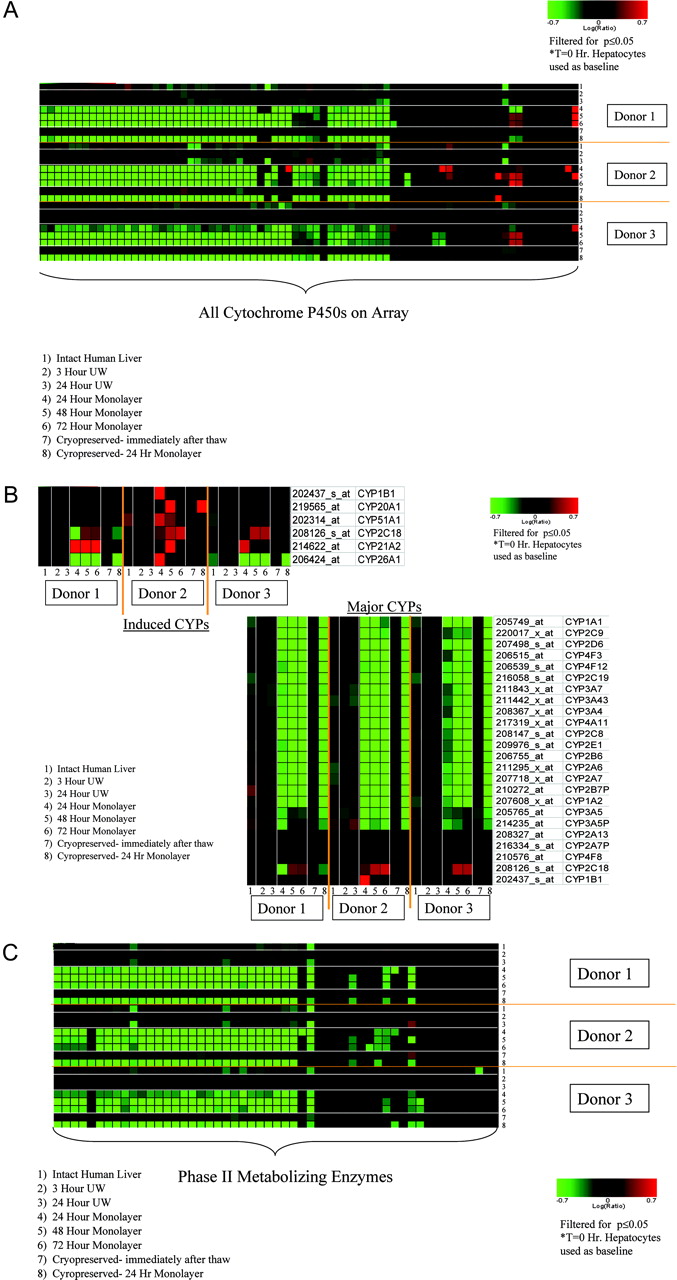
Hepatic phases I and II biotransformation reactions are critical to xenobiotic metabolism. Examination of P450 gene expression reveals a down-regulation of most transcripts upon plating of the primary hepatocytes relative to freshly isolated primary hepatocytes, whereas the P450 expression level is predominantly unchanged in samples from intact liver, cold storage, and immediate postcryopreservation (Fig. 4A). Most of the major P450 isoforms (CYP1, 2, 3, and 4) have a reduced expression level upon plating of the primary hepatocytes, with the exception of CYP2C18, which was slightly up-regulated upon plating for all donors. In addition, there are some cytochrome P450 genes (CYP1B1, CYP20A1, CYP51A1, CYP2C18, CYP21A2, and CYP26A1) that had elevated levels of expression upon primary hepatocyte plating, especially for donor 2 (Fig. 4B). However, many of these described P450s vary from donor to donor and are not induced in every donor or at every plating time point. Many gene products for the phase II conjugation system, including transcripts encoding several UDP-glucuronosyltransferases (UGTs), initially exhibited reduced levels of expression upon plating while remaining unchanged in samples from liver, cold storage, and postcryopreservation (Fig. 4, C and D). Most of the phase II genes that were regulated for the plated human hepatocytes were decreased quite consistently across all three donors evaluated.

Global gene expression profiles for all of the primary human hepatocyte donors at various stages of preparation. Each horizontal lane in the heat map represents a different stage of human hepatocyte preparation for its corresponding donor. Each vertical lane represents a single Affymetrix probe set. For the heat maps in all the figures, sequence agglomerative cluster analysis, using average link heuristic criteria and Euclidean distance for similarity measure, was performed to group together genes with similarities in gene expression. Genes represented were regulated at least ±2.0-fold with a p value of 0.01 or less. Increases in mRNA level are represented as shades of red with decreases in shades of green. If the p value for a particular gene expression change is greater than 0.01 (or other value as indicated for subsequent figures), the log(ratio) is represented as zero or black on a heat map.
Hepatic transport is a critical function for the uptake and excretion of xenobiotic compounds. Microarray analysis revealed that the greatest effect on transporter mRNA levels was seen in plated hepatocytes. Figure 4E shows that many of the solute carrier family of transporters have reduced levels of transcripts with respect to freshly isolated hepatocytes. For instance, SLC22A7 (solute carrier family 22), an organic anion transporter, was down-regulated after plating for all donors. Alternatively, there are transcripts such as SLC38A1, which is responsible for transport of glutamine (Gu et al., 2001), that have elevated levels upon primary hepatocyte plating. Most transporter mRNA levels remained unchanged for human hepatocytes in cold storage and immediately after cryopreservation, suggesting that the transporter gene expression changes result from plating of the primary hepatocytes. Furthermore, many transporter genes, such as ABCG5, SLCO2B1, and ABCB11, were consistently regulated across all three donors, whereas the expression of others, including ABCC3, SLC38A2, and SLC4A4, varied quite extensively from donor to donor.
Another important function of the liver pertains to its role in lipid and fatty acid metabolism. In particular, several genes involved with fatty acid degradation exhibited a trend of down-regulation. For the 24-h primary hepatocyte monolayer for donor 1, genes encoding enzymes such as carnitine palmitoyltransferase II, carnitine/acylcarnitine translocase, acyl-CoA dehydrogenases, and long-chain fatty acid CoA ligase 5 were all down-regulated relative to freshly isolated primary hepatocytes (Fig. 5A). In addition, a gene for short-chain 3-hydroxy-acyl-CoA dehydrogenase (probe ID 201035_at), an essential enzyme for mitochondrial β-oxidation catalyzing the conversion of 3-hydroxyacyl-CoAs to 3-ketoacyl-CoAs, was strongly reduced. Furthermore, for many transcripts, a trend of down-regulation is apparent with genes involved in lipid metabolism (as defined by gene ontology) for primary hepatocyte monolayers (Fig. 5B). Intact human liver, primary hepatocytes in cold storage, and postcryopreservation primary hepatocytes show the fewest gene expression changes for lipid metabolism.
The proteasome is the cellular machinery responsible for protein regulation, degradation, and recycling, and it serves a vital function in maintaining homeostasis. It is a multisubunit ubiquitin-dependent holoenzyme with a core complex responsible for hydrolytic activity (20 S core) and a regulatory complex with ATPase activity (19 S regulator) that mediates identification and proper trafficking of peptide substrates into the active site (Meiners et al., 2003). There was a significant up-regulation in the expression level of genes that encode for proteasome subunits upon plating of primary hepatocytes for all donors (Fig. 5C). More specifically, genes such as PSMD12 (a non-ATPase regulatory subunit), PSMD8 (a non-ATPase regulatory subunit), and PSMC6 (a subunit with ATPase activity) were all significantly overexpressed upon plating of the human hepatocytes (data not shown). This effect was largely not observed in samples from intact liver, primary hepatocyte cold storage, and postcryopreservation, which suggests the possibility that some aspect of the plating process may play a role in this enhancement of expression.
Apoptosis is a type of cell death that has distinct morphological and biochemical characteristics and that is under strict regulation. As displayed in Fig. 5D, the expression level of several key transcripts associated with induction of apoptosis were increased in primary hepatocyte monolayers relative to freshly isolated hepatocytes. For instance, caspases, which are enzymes involved in the execution phase of programmed cell death, such as CASP7 (all donors) and CASP3 (donor 1 in 24-h monolayer and donor 3), were significantly overexpressed in the plated human hepatocytes. Furthermore, other prominent players in apoptotic cell death exhibited enhanced expression in plated human hepatocytes such as poly(ADP-ribose) polymerase, an activator of cellular proliferation and of various nuclear proteins involved with DNA repair mechanisms. In addition, BID, involved in apoptosis activation via mitochondrial cytochrome c release, and c-myc, a transcription factor that assists with cell cycle regulation, were induced in samples of plated human hepatocytes for most donors and time points. The expression of these proapoptotic genes was changed upon cultivation of the primary hepatocyte monolayer but was largely unchanged in samples from intact liver, cold storage, and postcryopreservation.
Discussion
Gene Expression in Human Hepatocytes in Suspension after Isolation Is Similar to That of the Liver of Origin. The relatively few genes that were regulated in the intact livers versus freshly isolated hepatocytes tended to be down-regulated, suggesting that some of these expression changes may result from the fact that the liver is composed of a heterogeneous cell population, whereas primary hepatocyte cultures are a more homogenous population. These small changes in gene expression in hepatocytes after their isolation are consistent with our recent observation that P450-dependent and UGT-dependent activities are equivalent or only slightly decreased in microsomes prepared from human hepatocytes after their isolation compared with liver microsomes from the liver of origin (data not shown).
Gene Expression in Human Hepatocytes in Suspension Is Not Affected by Hepatocyte Cold Storage and Cryopreservation. The equivalent gene expression profile observed in suspension of freshly isolated human hepatocytes, after cryopreservation/thawing and after cold storage, was also in line with our recent observations that metabolic capacity of human hepatocytes in suspension after cold storage or after cryopreservation was intact and clearance predictions were equivalent to suspensions of freshly isolated hepatocytes from the same donor (Blanchard et al., 2005). Shitara et al. (2003) also observed that the transporter systems in human hepatocytes were functional after cryopreservation. Interestingly, gene expression for primary human hepatocytes in cold storage and cryopreservation was similar to that of the liver of origin, which could have implications for hepatocyte model systems being translated to intact liver.


A, gene expression changes for the cytochrome P450 transcripts present on the array. B, observed gene expression changes for the major P450 isoforms (right panel) and for those P450s that were induced (left panel). The majority of P450s were down-regulated upon plating the primary human hepatocytes, whereas cells in suspension or after cryopreservation showed few changes. C, underexpression of phase II transcripts upon plating the hepatocytes. D, gene expression changes for several UGTs during various stages of human hepatocyte preparation. E, regulation of transcripts associated with canalicular and basolateral transporter systems in primary human hepatocytes. The greatest number of changes in transporter expression is observed in plated primary human hepatocytes. For all panels, genes were filtered for p value of 0.05 or less.
The recovery of viable hepatocytes after a 24-h cold storage was at least equivalent to that after cryopreservation (see Table 2), suggesting that cold storage could be an alternative for human hepatocyte short-term shipping and storage. The data also suggest that P450 expression levels remain relatively stable at baseline levels upon cold storage of human hepatocytes. In addition, few gene expression changes were evident for critical hepatocellular processes, including transport, lipid metabolism, phase II metabolism, and protein processing, degradation, and recycling (proteasome subunits). Furthermore, human hepatocytes after cryopreservation and in cold storage may be less prone to initiating and undergoing apoptosis, as evidenced by the few expression changes for genes involved in apoptosis induction for these samples.

A, regulation of genes involved in the cellular fatty acid degradation pathway for 24-h monolayer human hepatocytes from donor 1 using GenMAPP visualization software (Dahlquist et al., 2002). An overexpressed gene is colored in red, and an underexpressed gene is colored in green. The quantitative -fold change for each gene is shown as a number to the right of the gene box. The statistical cutoffs for a gene being designated as regulated are at least ±2.0-fold change with a p value of 0.01 or less. B, a global view of the gene expression changed for fatty acid and lipid metabolism transcripts (as defined by gene ontology designation) for the various stages of human hepatocyte preparation. For many of these probe sets, transcription is down-regulated. C, expression profiles for genes that code for proteasomal subunits for the various stages of human hepatocyte preparation. These genes are overwhelmingly induced upon plating of human hepatocytes. For the heat maps, genes were filtered for a p value of 0.05 or less. D, a heat map for mRNAs that are associated with positive regulation of apoptosis (as defined by gene ontology). Many transcripts that code for proteins associated with apoptosis induction were induced upon hepatocyte plating.
Gene Expression in Human Hepatocytes Is Strongly Changed after Plating. Significant gene expression changes occur upon plating of hepatocytes. It has been previously shown that major cytochrome P450-dependent enzyme activities decline in culture, and this is associated with decreases in both gene and protein expression, symptomatic of a dedifferentiated state (Madan et al., 2003). The data of the present communication thus confirm these observations and highlight that some other P450 isoforms are, in contrast, up-regulated after plating. We (Richert et al., 2002; Binda et al., 2003) have previously suggested that the decline in P450 expression could be related to oxidative stress occurring during time in culture. Several genes associated with a cellular oxidative stress response, such as superoxide dismutase 2, glutathione reductase, p53, and peroxiredoxin, are overexpressed upon plating of human hepatocytes (data not shown). The present study therefore presents further evidence that oxidative stress occurs during plating and also further highlights that pathways leading to apoptosis are activated as well, most likely, at least in part, oxidative stress-related. In support of this, it has been reported that reactive oxygen species mediate apoptosis in hepatocytes (Rauen et al., 1999; Ishihara et al., 2005).
UGTs are a family of phase II enzymes critical for xenobiotic biotransformation reactions. The findings from this study indicate that the mRNA levels of several members of this class are reduced upon primary hepatocyte plating at the 24-h time point. At 72 h, there seems to be a slight trend toward increasing levels of mRNA relative to the 24- or 48-h time point for some UGT genes, especially for donor 2. This reflects a similar tendency observed in other reports with primary rat hepatocytes (Zurlo et al., 1996; Li et al., 1999). Furthermore, in activity measurements, UGTs retain basal metabolizing capabilities upon primary rat hepatocyte plating compared with whole liver (Richert et al., 2002). In addition, cryopreservation did not affect the gene expression of UGTs in this study in concordance with other studies (Li et al., 1999).
Hepatocellular transporter systems are essential regulators of homeostasis, are involved in drug disposition and bile formation, and may be critical for toxicological and pharmacological studies (Chandra and Brouwer, 2004). The data indicate that the expression of many transporter systems, especially those in the solute carrier family series, is decreased upon plating of the primary human hepatocytes. Messenger RNA from major basolateral transporters such as OATP2 (SLC22A7) and Na+/taurocholate-cotransporting peptide (SLC10A1) have been previously shown to be markedly reduced upon plating of primary rat hepatocyte cultures (Rippin et al., 2001; Luttringer et al., 2002). Our results also support this trend and, furthermore, offer evidence of down-regulation of additional basolateral transporters such as OCT1 (SLC22A1) and MRP6 (ABCC6) upon plating of primary human hepatocytes, in agreement with studies from other laboratories (Jigorel et al., 2005).
Hepatic canalicular transport proteins incorporate a variety of ATP-dependent membrane associated pumps that encompass the ATP binding cassette superfamily, including the well studied MDR1 or P-glycoprotein (Rippin et al., 2001; Chandra and Brouwer, 2004). In primary rat hepatocytes, MDR1b (ABCB1) and MRP3 (ABCC3) transcription products have been observed to increase upon plating and cultivation (Luttringer et al., 2002). Likewise, in plated primary human hepatocytes, we observe overexpression of ABCB1 mRNA relative to freshly isolated hepatocytes, which could result in more informed interpretation for studies involving xenobiotic metabolism of ABCB1 substrates. Other canalicular pumps, such as ABCB11 and ABCB4, tend to be underexpressed in plated primary human hepatocytes for these evaluated donors.
To our knowledge, the present study is the first report on drastic changes in the expression of genes involved with fatty acid and lipid metabolism in primary human hepatocytes after plating as determined by microarray analysis. A large number of lipid metabolism transcripts are underexpressed in monolayers of primary human hepatocytes relative to their freshly isolated counterparts. This, in part, could be related to an oxidative stress response because both arachidonic acid mobilization and phospholipase A2 activity were found to be affected by oxidative stress (Babenko et al., 1998). Even more relevant to the present results, a direct link between oxidative stress and ApoB100 degradation and very low-density lipoprotein secretion has been recently reported (Pan et al., 2004).
It has been well documented that free fatty acids can have dramatic direct or indirect effects on expression of genes involved in lipid metabolism via regulation of nuclear transcription factors such as the peroxisome proliferator-activated receptors, sterol regulatory element-binding proteins, liver X receptors, or hepatic nuclear factor (Jump, 2002; Swagell et al., 2005). Although no significant gene expression regulation of peroxisome proliferator-activated receptors (α, β, or γ) or hepatic nuclear factor 1 was detected, LXRa (203920_at) was underexpressed for all donors, and SREBP1 (202308_at) was underexpressed for donors 1 and 3 only in the monolayer cultures relative to freshly isolated human hepatocytes (data not shown). Perhaps changes in the abundance and variety of fatty acids from intact liver to cell culture may be at least partially responsible for the observed changes in expression.
Interestingly, many proteasomal subunits, the proteolytic machinery responsible for degradation and recycling of ubiquitin-protein complexes, are overexpressed in primary human hepatocyte monolayers compared with freshly isolated hepatocytes. Increased expression of these subunits may be associated with an increased proteasome presence and, ultimately, potentially elevated proteolytic activity relative to the native state (Hobler et al., 1999; Fang et al., 2000). A recent communication highlights that mild oxidative stress may augment proteasome activity and expression in response to a heightened level of oxidatively damaged proteins (Elkon et al., 2004), whereas severe oxidative stress may result in proteasome failure and in abolishment of proteasome activity. Therefore, it is possible that some degree of oxidative stress in plated human hepatocytes may drive, in part, the overexpression of these proteasomal subunits. Awareness of this potential increased proteolytic activity and turnover may assist in some types of data interpretation for primary human hepatocyte studies.
Gene Expression after Plating Is Not Affected by Cold Storage or Cryopreservation. In the present study, we found that the gene expression pattern for the most of the major genes evaluated in plated cryopreserved/thawed human hepatocytes was similar to that of their respective plated fresh monolayers (24-h time point) for all donors. One should be aware, however, that in the present study, the plating density of thawed hepatocytes was 50 to 80% of the plating density of their respective freshly isolated hepatocytes, and this could contribute to the similar pattern of gene expression for plated thawed human hepatocytes. Indeed, in a previous study, we found that human hepatocytes in culture displayed markedly different gene expression patterns depending on attachment status (Waring et al., 2003), in accordance with the observations of Hamilton et al. (2001), who described a decrease in constitutive levels of CYP3A4 in human hepatocytes that correlated with the plating density. Nonetheless, it is noteworthy that in the latter work, the -fold induction of CYP3A4 observed after exposure to rifampicin was equivalent in cultures as long as the cell density was not reduced to more than 50% of standard density. Taken together, these results suggest that plated cryopreserved hepatocytes are a valid tool for drug metabolism and toxicological evaluations as long as the plating density is at least 50% of that of a confluent monolayer.
It is possible that the expression profiles for some genes in other human donors may behave differently than observed in this study because of the large degree of primary human hepatocyte donor-to-donor heterogeneity (Liguori et al., 2005). However, it is clear from these results that plating of primary human hepatocytes leads to a vigorous change in gene expression from the native state, which may be a critical factor for some studies involving human hepatocytes. Taken together, these results may aid in the interpretation of data collected from human hepatocyte experiments and suggest additional utility for cold storage and cryopreservation of hepatocytes.
Footnotes
-
L.R. and M.J.L. contributed equally to this work.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.007708.
-
ABBREVIATIONS: P450, cytochrome P450; DMEM, Dulbecco's modified Eagle's medium; UW, University of Wisconsin solution; UGT, UDP-glucuronosyltransferase; CoA, coenzyme A.
-
↵
 The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material. - Received October 3, 2005.
- Accepted February 3, 2006.

- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}