


Abstract
CYP2C9 is one of the major drug-metabolizing enzymes, and it is involved in the oxidative metabolism of approximately 10% of clinically important drugs, among which some, such as the anticoagulant warfarin, have a narrow therapeutic index. The human CYP2C9 gene is highly polymorphic. We found a new sequence variation in exon 7 of the CYP2C9 gene (1060G>A) resulting in a substitution of acidic amino acid glutamate to basic lysine (E354K) when translated. The allele, designated CYP2C9*24, was present in heterozygous state in one warfarin-treated patient. To characterize the CYP2C9*24 allele, we expressed the wild-type and CYP2C9.24 protein in a recombinant yeast expression system and a human embryonic kidney (HEK)-293 cell system. Carbon monoxide difference spectra were recorded on dithionite-reduced microsomes, and protein was determined by Western blotting. Transfection with CYP2C9.1 cDNA resulted in detectable CYP2C9 protein in yeast or HEK-293 cells, whereas only small amounts of the protein were detected in yeast transfected with CYP2C9.24 cDNA. A strong differential absorption peak at 450 nm was observed with microsomes of yeast transfected with CYP2C9.1 cDNA, whereas no peak was detected with microsomes of yeast transfected with CYP2C9.24 cDNA or empty pYeDP60 plasmid. These results suggest that CYP2C9.24 may be improperly folded, both in yeast and mammalian cells, resulting in improper heme incorporation and rapid intracellular degradation. The data obtained in the expression systems are consistent with our findings in vivo. In conclusion, we have identified a novel defective CYP2C9 variant allele of potential importance for drug metabolism in vivo.
CYP2C9 is involved in the metabolism of more than 100 currently used drugs, corresponding to about 10 to 20% of commonly prescribed drugs (Kirchheiner and Brockmoller, 2005). Some of them (e.g., anticoagulant warfarin) have a narrow therapeutic index (Miners and Birkett, 1998). Clinically available warfarin is a racemic mixture, and CYP2C9 is responsible for the metabolism of more potent S- enantiomer (Kaminsky and Zhang, 1997). The human CYP2C9 gene is highly polymorphic (http://www.imm.ki.se/CYPalleles/). The two most common variant alleles in Caucasian populations are CYP2C9*2, with a point mutation in exon 3 (430C>T), and CYP2C9*3, with a point mutation in exon 7 (1075A>C). Both are associated with a decrease in the catabolic activity of the enzyme (Rettie et al., 1994; Sullivan-Klose et al., 1996). It was shown in many studies that the patients with polymorphic alleles require significantly lower doses of warfarin and are more susceptible to bleeding complications than the carriers of two wild-type alleles (Kirchheiner and Brockmoller, 2005).
We have previously reported a novel 1060G>A sequence variant in exon 7 of CYP2C9 gene, leading to substitution of acidic amino acid glutamate to basic lysine (E354K). The new allele, designated CYP2C9*24, was present in the heterozygous state in one warfarin-treated patient, who was also a heterozygous carrier of the CYP2C9*2 allele. Because exons 3 and 7 are separated by a long distance (>39 kilobase pairs), we could not assess whether these two alleles were located on the same or separate alleles.
The carrier of the CYP2C9*24 allele was 77-years-old and was treated with warfarin because of atherosclerotic cardiomyopathy with atrial fibrillation. The index patient had the lowest daily warfarin dose requirement (1.5 mg/day) and the lowest (S)-warfarin clearance (0.809 ml/min) when compared with a group of warfarin patients matched with the index patient for age, indication, drug cotreatment, and CYP2C9*1/*2 heterozygous genotype [mean (S.D.) warfarin dose 3.04 (1.16) mg/day and (S)-warfarin clearance 2.400 (0.864) ml/min] (Herman et al., 2006).
To assess the functional importance of the CYP2C9*24 allele, we expressed the wild-type and CYP2C9.24 protein in a recombinant yeast expression system and a mammalian human embryonic kidney (HEK)-293 cell system.
Materials and Methods
Preparation of CYP2C9 Expression Constructs. The yeast expression vector pYeDP60, which contained the wild-type CYP2C9 cDNA (AstraZeneca, R&D, Umeå, Sweden), was used as a template to generate mutated CYP2C9*24 cDNA. QuikChange Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA) was used to introduce 1060G>A mutation with mutagenic oligonucleotides mut2C9_F (5′-CAGATGCTGTGGTGCACAAGGTCCAGAGATACATTG-3′) and mut2C9_R (5′-CAATGTATCTCTGGACCTTGTGCACCACAGCATCTG-3′). The CYP2C9*1 and CYP2C9*24 alleles were amplified from pYeDP60 vector using primers that introduced KpnI restriction site before the start codon (5′-ATAGGTACCATGGATTCTCTTGTGGTCC-3′) and EcoRI site after the stop codon (5′-ATAGAATTCTCAGACAGGAATGAAGCAC-3′). Fragments were purified by QIAquick Gel Extraction Kit (Qiagen, Valencia, CA) and subcloned into pcDNA3.1 (Invitrogen, Carlsbad, CA). All the constructs were verified by sequencing.
Expression in Yeast. pYeDP60 containing CYP2C9*1 or CYP2C9*24 cDNA were transfected by lithium acetate procedure in Saccharomyces cerevisiae strain W(R) that overexpresses yeast reductase (Pompon et al., 1996). Yeast cells transfected with empty pYeDP60 were used as a negative control. After 2 to 4 days of incubation at 28°C, the colonies were transferred to a new SGI (Mountain View, CA) plate (l g/l casamino acids, 7 g/l yeast nitrogen base, 20 g/l glucose, and 20 mg/l tryptophan) making 2- to 3-cm lines and incubated for an additional 2 to 4 days at 28°C until the yeast had grown thick. Then the yeasts were transferred into 20 ml of SGI medium and grown overnight at 28°C until OD600 nm was around 10. The culture was poured into 230 ml of yeast peptone glycerol ethanol medium (10 g/l yeast extract, 10 g/l bacto-peptone, 5 g/l glucose, and 2% ethanol) and grown at 28°C to OD600 nm around 10 to 13. Induction was started by the addition of 28 ml of 20% galactose, and the growth was continued until OD600 nm reached 30 (Pompon et al., 1996).

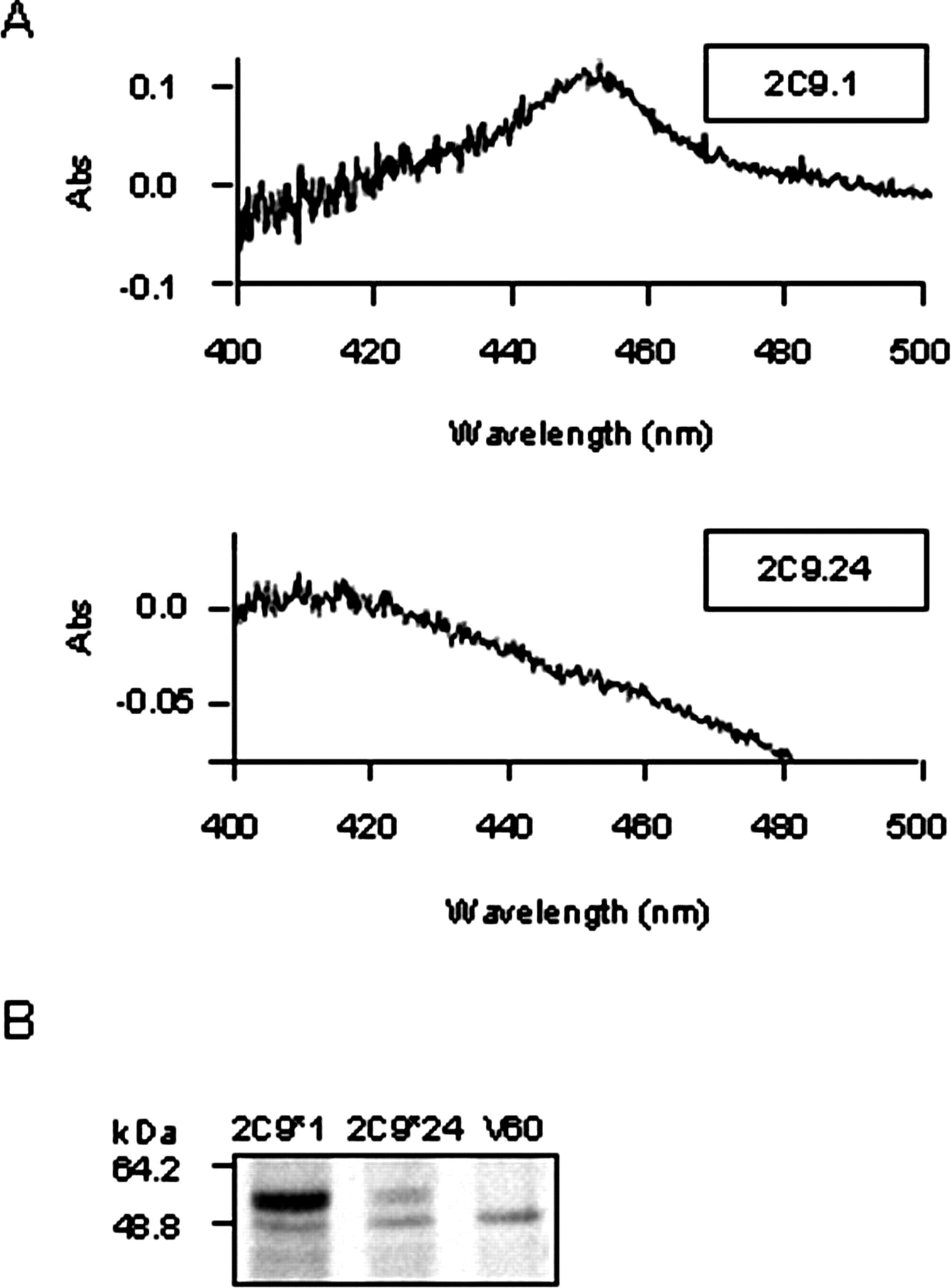
Analysis of CYP2C9 expressed in yeast. A, carbon monoxide-binding spectra analysis of yeast cells transformed with wild-type CYP2C9*1 and CYP2C9*24 cDNA cloned in pYeDP60 (V60) expression vector. B, Western blot analysis of CYP2C9 using CYP2C9 antibody. Microsomes were prepared from yeast cells transformed with CYP2C9*1 and CYP2C9*24 cDNA or empty pYeDP60. Thirty micrograms of protein was loaded per lane.
The yeast cells were recovered by centrifugation, washed once with distilled water, and resuspended in Tris EDTA KCl buffer (50 mM Tris-HCl, pH 7.4, 1 mM EDTA, and 0.1 M KCl). Cells were spun down again and resuspended in 2.5 ml of TES buffer (50 mM Tris-HCl, pH 7.4, 1 mM EDTA, and 0.6 M sorbitol), with added Complete protease inhibitor mixture (Roche Applied Science, Pentzberg, Germany). The yeast cells were mechanically disrupted using glass beads. Five milliliters of cold TES buffer was added to the crude extract three times, and the supernatant was withdrawn and centrifuged at 15,000g for 10 min at 4°C. The supernatant was then transferred to a new tube and centrifuged at 34,000g for 1 h at 4°C. The pellet, consisting of microsomes, was dissolved in 2.5 ml of cold Tris EDTA glycerol medium (50 mM Tris-HCl, pH 7.4, 1 mM EDTA, and 20% glycerol) (Pompon et al., 1996). The protein concentration was determined according to Lowry et al. (1951). The carbon monoxide-binding spectra analysis was recorded on dithionite-reduced microsomes (Omura and Sato, 1964). Yeast transfection and all the subsequent analysis were repeated three times.
Expression in HEK-293 Cells. Cells were grown at 37°C in atmosphere of 5% CO2 in minimum essential medium supplemented with fetal bovine serum, sodium pyruvate, nonessential amino acids, and antibiotics (Invitrogen). When HEK-293 cells reached 80 to 90% confluence, they were transiently transfected with pcDNA3.1 constructs using Lipofectamine 2000 transfection reagent (Invitrogen). Empty pcDNA3.1 was used as a negative control. After 60 h, the cells were harvested, homogenized, and fractionated by centrifugation (Karlgren et al., 2004). The protein concentration of microsomal fraction was determined according to Lowry et al. (1951), and the carbon monoxide-binding spectra analysis was recorded on dithionite-reduced microsomes (Omura and Sato, 1964). Transfection of HEK-293 cells and all the subsequent analysis were repeated three times.
Western Blot Analysis. Microsomes from transfected yeast or HEK-293 cells, corresponding to 30 μg of protein, were separated on 10% SDS-polyacrylamide gels and transferred to a Hybond-C extra membrane (Amersham Biosciences, Uppsala, Sweden). The membrane was first incubated with a CYP2C9 antibody diluted 1:5000 (BD Bioscience, San Jose, CA) and then with the secondary antibody conjugated with horseradish peroxidase diluted 1:2000 (BD Bioscience). SuperSignal West Pico Chemiluminescent Substrate (Pierce, Rockford, IL) was used for detection. Molecular weight marker was the BenchMark Prestained Protein Ladder (Invitrogen). Membranes were scanned using luminescent image analyzer LAS-3000 (Fujifilm Life Science, Stamford, CT).
Results and Discussion
Most of the genes encoding enzymes involved in drug metabolism are genetically diverse. CYP genes, in particular, have higher density of polymorphic sites and higher genetic diversity than any other human genes examined. Among subfamilies involved in phase I drug metabolism, CYP2 displays the highest level of genetic diversity. It is particularly important to predict the function of nonsynonymous sequence variation in CYP genes and to correlate that function to phenotypic observation because of their role in drug metabolism and adverse drug reactions (Solus et al., 2004).
In the present study, we have characterized a rare variant CYP2C9*24 allele that was previously found in one warfarin-treated patient (Herman et al., 2006). We found it interesting because the 1060G>A mutation resulted in substitution of glutamate to lysine (E354K) within the region where many genetic polymorphisms were reported to affect enzyme activity (CYP2C9*3, CYP2C9*4, and CYP2C9*5). Indeed, the patient, who was a heterozygous carrier of CYP2C9*24 allele, had slower metabolism of (S)-warfarin and consequently required a lower warfarin dose than other warfarin-treated patients matched for age, indication, drug cotreatment, and CYP2C9*1/*2 heterozygous genotype (Herman et al., 2006), indicating that 1060G>A mutation may be of functional importance.
To study a potential functional role of CYP2C9*24 allele, CYP2C9 cDNA with and without 1060G>A mutation was first expressed in yeast S. cerevisiae strain W(R) because it was previously shown that CYP2C9 protein can be efficiently expressed in this system (Yasar et al., 2001). Carbon monoxide difference spectra analysis of microsomal fractions revealed a strong peak at 450 nm for yeast transfected with pYeDP60 constructs containing wild-type CYP2C9 cDNA (Fig. 1A), indicating the presence of correctly folded cytochrome P450 enzyme (about 400 pmol/mg microsomal protein). On the other hand, no peak at 450 nm was observed for yeast transfected with CYP2C9*24 cDNA or with empty pYeDP60 plasmid. By Western blot analysis we showed that transfected yeast produced a highly detectable level of CYP2C9.1 protein, whereas only a small amount of CYP2C9.24 protein was seen in all the independent replications of transfection (Fig. 1B). These results suggested that CYP2C9.24 protein might be improperly folded in yeast cells, presumably because of improper heme incorporation, and therefore might be subjected to a rapid intracellular degradation. Although it could also be possible that protein synthesis was decreased or immunoreactivity of the variant protein changed, decreased protein levels and faster proteolytic degradation rate were recently reported in variant CYP1B1 allele with N453S substitution (Bandiera et al., 2005).

Analysis of CYP2C9 expressed in mammalian HEK-293 cells. A, carbon monoxide-binding spectra analysis of HEK-293 cells transformed with wild-type CYP2C9*1 and CYP2C9*24 cDNA cloned in pcDNA3.1 expression vector. B, Western blot analysis of CYP2C9 using CYP2C9 antibody. Microsomes were prepared from HEK-293 cells transformed with CYP2C9*1 and CYP2C9*24 cDNA or empty pcDNA3.1. Thirty micrograms of protein was loaded per lane.
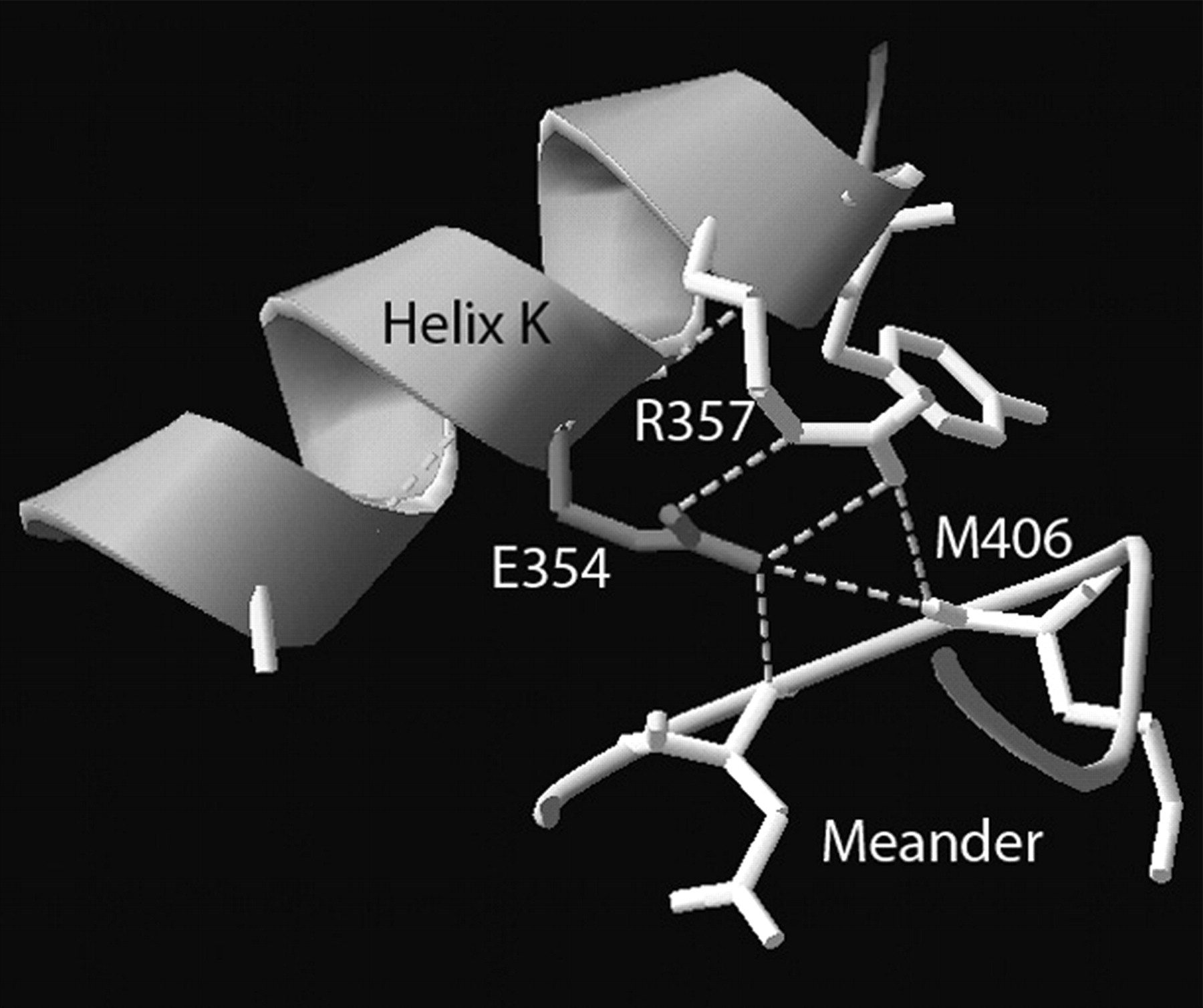
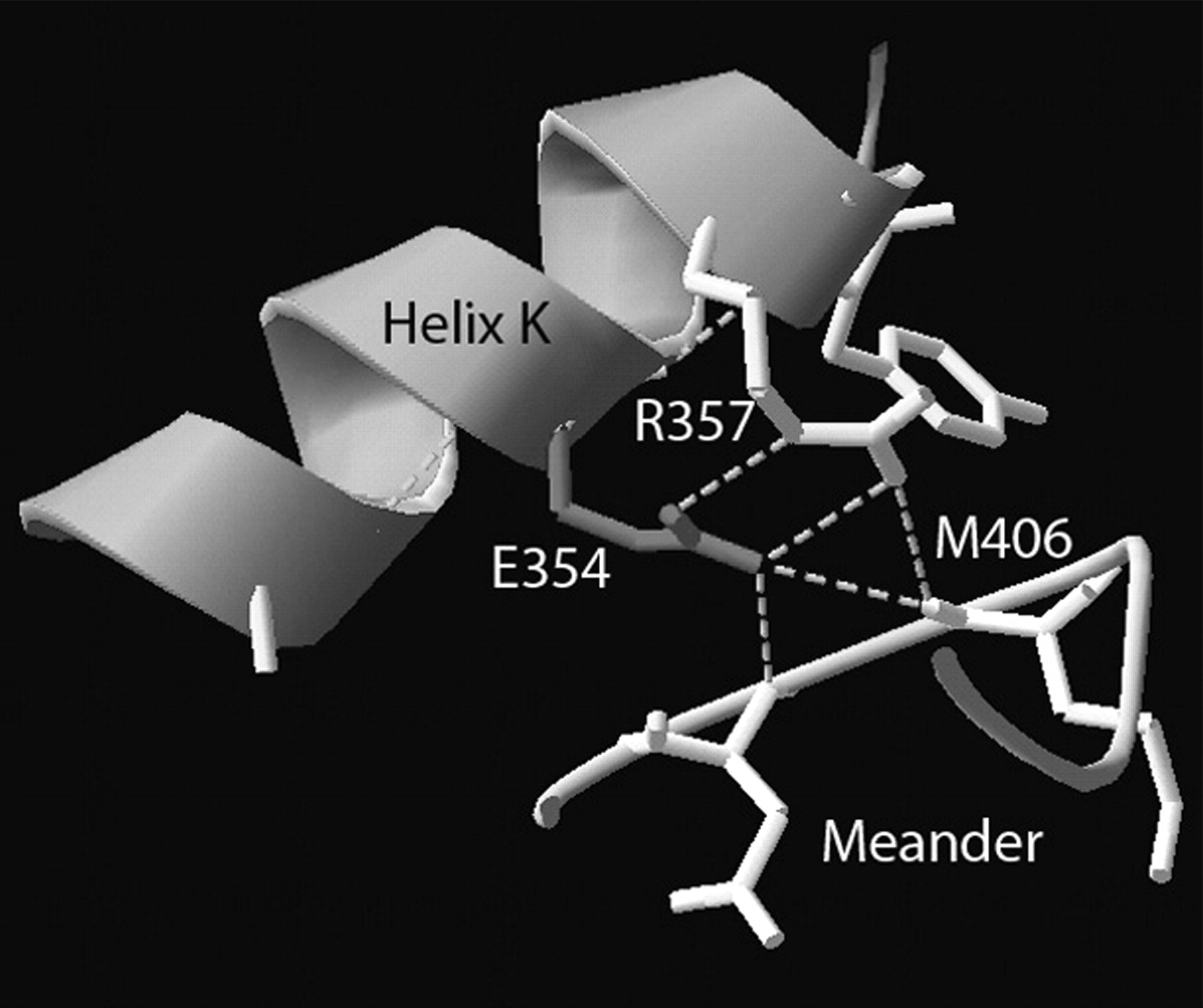
View of glutamate residue at position 354 present in the helix K. E354 forms hydrogen bonds (indicated by dashed lines) to R357, and together they form hydrogen bonds to the backbone of the meander region. This figure was produced from CYP2C9 crystal structure image (http://www.rcsb.org/pdb/explore.do?structureId=1OG5) using DeepView (http://www.expasy.org/spdbv/).
To further test this hypothesis, we heterologously expressed wild-type and mutated CYP2C9 proteins in mammalian HEK-293 cells. Carbon monoxide difference spectral analysis of the microsomal fraction obtained from transfected HEK-293 cells showed that only wild-type CYP2C9.1 protein was correctly folded (60 pmol/mg microsomal protein) (Fig. 2A), whereas no cytochrome P450 carbon monoxide spectrum was observed for the cells transfected with CYP2C9*24 cDNA or with empty pcDNA3.1 plasmid. Western blot analysis revealed that only cells transfected with wild-type construct produced detectable levels of CYP2C9.1 in all the independent replications of transfection (Fig. 2B). These results were confirmative of those obtained from the yeast expression system. The only difference was that in the HEK-293 expression system not even a small amount of incorrectly folded protein could be detected. Similar discrepancies have been reported between yeast and mammalian systems (Oscarson et al., 1999), suggesting that yeast may be a more permissive expression system for poorly stable proteins.
E354 is positioned in the carboxyl-terminal portion of the helix K and is a part of the absolutely conserved EXXR sequence, which may be involved in stabilizing the core structure. This sequence is buried in the interior of the protein and participates in a hydrogen bond network, together with the amino acid residues from the meander region (Fig. 3). Meander region is also highly conserved in structure (Graham and Peterson, 2002). The substitution of glutamate to lysine would disrupt the salt bridges between E354, R357, and M406 from the meander region. The bigger size of the lysine is also inconsistent with tight packing interactions with the amino acid residues that surround E354. Therefore, the substitution would probably result in disruption of the interaction of helix K with the meander region and would lead to protein misfolding or instability, which was also suggested by our experimental data.
In conclusion, our data suggest that the substitution of acidic amino acid glutamate to basic lysine may lead to incorrectly folded CYP2C9.24 protein that is rapidly degraded in both yeast and mammalian cells. Furthermore, our data support our observations of lower warfarin dose requirements and lower (S)-warfarin clearance in a patient with the CYP2C9*24 allele (Herman et al., 2006).
Acknowledgments
We thank Mats Hidestrand, Karolinska Institute, Stockholm, Sweden, for providing the wild-type CYP2C9 cDNA and for assistance in yeast transformation, and Eric F. Johnson, The Scripps Research Institute, La Jolla, CA, for comments on the possible structural alterations in CYP2C9.24.
Footnotes
-
doi:10.1124/dmd.106.013722.
-
This work was financially supported by The Swedish Research Council and National Institutes of Health (National Institute of General Medical Sciences Grant 1-R01 GM60548) and Slovenian Research Agency (P0-0503-0381). The stay of Darja Herman in Sweden was supported by the Swedish Institute scholarship for advanced study/research.
-
ABBREVIATIONS: HEK, human embryonic kidney; TES, 2-{[2-hydroxy-1,1-bis(hydroxymethyl)ethyl]amino}ethanesulfonic acid.
- Received November 7, 2006.
- Accepted February 26, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}