


Abstract
The neonatal Fc receptor (FcRn) plays a critical role in maintaining homeostasis of IgG antibodies. Recent studies have shown that the FcRn-IgG interaction can be modulated to alter the pharmacokinetics of the antibody. This has been achieved by altering amino acid residues in the FcRn-binding domain of the antibody, resulting in a change in the pH-dependent binding affinity of the antibody to FcRn. The purpose of this study was to examine the impact of the pH-dependent FcRn binding affinity on the pharmacokinetics of the antibody with changes in the Asn434 residue. Two anti-tumor necrosis factor-α monoclonal antibody (mAb) FcRn variants (N434A and N434H) were engineered, and pharmacokinetic studies of the two FcRn variants together with the wild type (WT) were conducted in mice and cynomolgus monkeys. N434A, which had binding properties to murine FcRn similar to those of the WT, had the same pharmacokinetic profile as the WT in mice. N434H, with the highest binding affinity to murine FcRn at pH 7.4, had a faster clearance (16.1 ml/day/kg) and a lower bioavailability (61.3%) compared with the WT (5.07 ml/day/kg, 73.2%) and N434A (5.90 ml/day/kg, 72.4%) in mice. N434A and N434H, which had higher binding affinity at pH 6.0 to monkey FcRn with comparable affinity at pH 7.4, had significantly higher areas under the serum concentration-time curve from time 0 to day 7 than the WT (749 ± 71.9 and 819 ± 81.5 versus 592 ± 56.8 μg/ml · day) in monkeys. Thus, increasing the binding affinity of mAbs to FcRn at pH 6.0 while keeping a low binding affinity at pH 7.4 improves the pharmacokinetics of these molecules.
The neonatal Fc receptor (FcRn) was first postulated by Brambell et al. in 1964 and cloned in the late 1980s (Simister and Mostov, 1989). FcRn is a heterodimer comprising a β2-microglobulin (β2m) light chain and a major histocompatibility complex class I-like heavy chain. It is expressed widely in endothelial and epithelial cells including organs such as the skin, muscle, kidney, liver, and placenta. A unique feature of the interaction of FcRn with its ligands is strict pH dependence. FcRn binds to IgG in the acidic environment (pH 6.0) of the endosome and releases it at physiological pH. IgG that is not bound to FcRn in the endosome proceeds to the lysosome and undergoes proteolysis. The binding site for FcRn on IgG is located within the interface of the CH2-CH3 domain of the IgG-Fc (Burmeister et al., 1994). The pH dependence is due to the presence of conserved histidine residues (His310 and His435) that are protonated at an acidic pH and deprotonated or neutral at physiological pH (Kim et al., 1994, 1999). Additional Fc residues that appear to be critical for Fc-FcRn binding in the mouse are Ile253, His433, and His436, whereas Ile253, Ser254, and Tyr436 have been found to be important for human Fc-FcRn binding (Kim et al., 1994; Hinton et al., 2006).
The role of FcRn in protecting IgG from catabolism has been well established. Several studies have shown that IgG clearance in β2m knockout mice (Ghetie et al., 1996; Junghans and Anderson, 1996) and FcRn-heavy chain knockout mice (Roopenian et al., 2003) is increased 10- to 15-fold, with no changes in the elimination of other Igs. It was also observed that low levels of serum IgGs and albumin in patients with familial hypercatabolic hypoproteinemia correlated with a reduction in FcRn expression resulting from the β2m deficiency (Wani et al., 2006). There is also growing evidence that the half-life of an IgG can be influenced by altering its binding affinity to FcRn. This has been achieved by altering amino acid residues in the FcRn-binding domain of the antibody (Dall'Acqua et al., 2002, 2006; Hinton et al., 2004, 2006; Petkova et al., 2006; Yeung et al., 2009). Two engineered human IgG1 variants (N434A and T307A/E380A/N434A) with enhanced binding affinity to human FcRn cleared more slowly and had approximately 2-fold longer half-lives in human FcRn-transgenic mice than in the wild-type mice (Petkova et al., 2006). Hinton et al. (2006) reported that the half-life of IgG1 and IgG2 FcRn variants (M428L and T250Q/M428L) with increased binding affinity to human and monkey FcRn at pH 6.0 but with no change in binding affinity at pH 7.5 was approximately 2-fold longer than that of the wild-type antibody in rhesus monkeys. It was also recently reported that the half-life of a N434A variant, which had 4-fold improved binding affinity at pH 6.0 over that of the wild-type (WT) to human and nonhuman primate FcRn, was 14.5 days in cynomolgus monkeys compared with that in the WT, which had a half-life of 6.2 days (Yeung et al., 2009). However, in the same study, the authors did not see an improvement in the half-life of the N434W variant. This finding was attributed to the increased binding affinity of the variant at pH 7.4, which negated the positive effect on half-life by the improved binding affinity at pH 6.0. These data are also in agreement with the previous report of Dall'Acqua et al. (2002) and in line with what would be theoretically expected given the pH dependence of the FcRn-IgG interaction. Although several studies have shown a correlation between increased binding affinity of an IgG to FcRn at pH 6.0 with no change in binding properties at pH 7.4 and increased half-life, there are some studies that did not observe this effect (Gurbaxani et al., 2006; Datta-Mannan et al., 2007a,b). Datta-Mannan et al. (2007a,b) reported no difference in the half-life of the FcRn variants (T250Q/M428L, P257I/Q311I, P257I/N434H, and D376V/N434H) compared with that of the wild-type antibody in monkeys. We were surprised to find that the pharmacokinetics of their T250Q/M428L variant in the cynomolgus monkey is in contrast to that reported by Hinton et al. (2004, 2006) for the same variant. Likewise, no differences in half-life were reported with the T307A/E380A/N434A variant in normal mice (Gurbaxani et al., 2006; Petkova et al., 2006). It has recently been suggested that FcRn may play an important role in monoclonal antibody (mAb) bioavailability after subcutaneous administration (Wang et al., 2008). The systemic bioavailability of 7E3, a monoclonal IgG1 antibody, was ∼3-fold fold higher in WT mice than in FcRn-deficient mice (Wang et al., 2008). In the light of these data, our studies were focused on evaluating the pharmacokinetics of the Asn434 variants and also on examining the impact of FcRn affinity at different pH on the antibody pharmacokinetics including bioavailability. A WT anti-TNF-α mAb IgG1 and its two FcRn variants, N434H and N434A, which showed different FcRn binding properties at pH 6.0 and pH 7.4, were evaluated in mice and monkeys. We observed that increasing the binding affinity to FcRn at pH 6.0 while maintaining the low binding affinity at pH 7.4 improved exposure. In contrast, increasing the binding affinity to FcRn at pH 7.4 decreased the exposure and also appeared to reduce the subcutaneous bioavailability.
Materials and Methods
Antibodies.
Full-length humanized anti-TNF-α mAb IgG1 and its FcRn variants (N434H and N434A) were generated at Genentech Inc. (South San Francisco, CA) and formulated in phosphate-buffered saline, pH 6.0. N434A and N434H indicate mutations of Asn434 to Ala434 and His434, respectively. Soluble murine, cynomolgus monkey, and human FcRns were expressed and purified as reported previously (Yeung et al., 2009).
Measurements of mAb Binding Affinity to FcRn.
The equilibrium dissociation constant, KD, was measured using a BIAcore 3000 instrument (GE Healthcare, Little Chalfont, Buckinghamshire, UK) as reported previously (Yeung et al., 2009). In brief, murine, cynomolgus monkey, and human FcRns were amine-coupled to CM5 sensor chips. Serial dilutions of antibody (WT or FcRn variants) were injected in phosphate-buffered saline containing 0.05% Tween 20, pH 6.0 or pH 7.4. The bound antibody was measured as response units at equilibrium, and KD was estimated by plotting the response units at equilibrium against injected antibody concentrations.
Pharmacokinetic Study in SCID Mice.
The pharmacokinetic (PK) study in SCID mice was performed following animal protocols approved by the Institutional Animal Care and Use Committee at Genentech Inc. SCID female mice received a single intravenous or subcutaneous dose of 5 mg/kg WT anti-TNF-α mAb, N434A, or N434H (n = 12/group). Blood samples were collected from the periorbital sinus or cardiac puncture in the case of terminal time points for PK at the following time points: 15 and 30 min; 1, 8, and 24 h; and 3, 7, 10, 14, 18, 21, and 28 days, and transferred into serum separator tubes. Three mice per group underwent periorbital bleeding for each time point, with 150 μl of blood collected per mouse; these collections were staggered such that blood was drawn from an individual mouse only for every fourth time point.
Pharmacokinetic Study in Cynomolgus Monkeys.
The PK study in cynomolgus monkey was approved by the Institutional Animal Care and Use Committee at Shin Nippon Biomedical Laboratories USA, Ltd. (Everett, WA) and conducted at Shin Nippon Biomedical Laboratories USA. Six cynomolgus monkeys (three males and three females) in each of three groups were given a single intravenous dose of 5 mg/kg WT anti-TNF-α mAb, N434A, and N434H, respectively. The test article was administered on day 0 intravenously into a superficial vein on the arm or leg. Blood samples (∼1.5 ml) for PK and anti-therapeutic antibody (ATA) analysis were collected from each animal by venipuncture of the saphenous, femoral, or cephalic vein and transferred into serum separator tubes. Samples were taken from all animals twice during the acclimation period (on days A4 and A11) and on days 0 (at predose, 30 min, and 6 h postdose), 1, 3, 5, 7, 14, 21, 28, 35, and 42. Additional samples were collected from animal 13 in the N434H group on days 49, 56, 63, 70, 77, and 84.
Bioanalysis of Serum Samples from Pharmacokinetic Studies.
Serum samples from SCID mice and cynomolgus monkeys were analyzed by a quantitative enzyme-linked immunosorbent assay as described previously (Loyet et al., 2009) to measure the serum concentration of total WT mAb and its two FcRn variants. First, 384-well microtiter plates (Nalge Nunc International (Rochester, NY) were coated with human TNF-α. This recombinant protein was produced at Genentech Inc. (lot N9017A2) and diluted to 0.5 μg/ml in phosphate-buffered saline, pH 7.2. Anti-TNF-α WT (PUR 11590; produced at Genentech Inc.) was used as the standard. The standard curve ranged from 0.781 to 50.0 ng/ml. Standard curves, controls, and samples were diluted in assay diluent (phosphate-buffered saline, 0.5% bovine serum albumin, 0.05% polysorbate 20, 0.2% bovine γ globulin, 0.25% CHAPS, 5 mM EDTA, 0.35 M NaCl, and 10 ppm ProClin 300). Samples were diluted a minimum of 1:100. Horseradish peroxidase-labeled goat F(ab′)2 fragment anti-human IgG Fc was obtained from Jackson ImmunoResearch Laboratories Inc. (West Grove, PA) and used as the conjugate. The signal was generated using tetramethyl benzidine (Kirkegaard and Perry Laboratories, Gaithersburg, MD) as the substrate for horseradish peroxidase. Sample results were corrected for dilution; values within the standard curve range were averaged. The assay performance is similar for anti-TNF-α WT and its two FcRn variants. The minimum quantifiable concentration for all three anti-TNF-α mAbs in undiluted cynomolgus monkey and SCID mouse serum was determined to be 78 ng/ml, based on the assay limit of 0.781 ng/ml and the minimum dilution of 1:100.
ATA Assay.
Serum samples from cynomolgus monkeys were analyzed by a bridging electrochemiluminescence assay to determine the level of ATA response against anti-TNF-α mAbs (Loyet et al., 2009). Cynomolgus monkey serum containing polyclonal antibodies directed against human IgG was obtained from Genentech Inc. (Animal CQ5003 lot 26404-46E, prepared February 4, 1997) and was used as the surrogate positive control. Controls and samples were diluted into ATA assay diluent (80 mM HEPES buffer, 0.5% fish gelatin, 0.25% polysorbate 20, 0.25% CHAPS, 0.35 M NaCl, and 0.05% ProClin300, pH 7.0) and added to 96-well, round-bottom polypropylene plates (Corning Inc., Corning, NY). The samples were diluted a minimum of 1:20. Biotinylated anti-TNF-α mAb WT and BV-tagged anti-TNF-α mAb WT (prepared according to the manufacturer's instructions; BioVeris, Gaithersburg, MD) were added to start the assays. After 1.5 to 2 h, streptavidin-coated Dynabeads M-280 (Invitrogen, Carlsbad, CA) were diluted in ATA assay diluent and added to the plates. The plates were then read in an M-series M384 analyzer (BioVeris), and the assay response was measured in electrochemiluminescence units. The cut point was set based on the value of the negative control (predose serum pool from 28 untreated cynomolgus monkeys) and was used to calculate the titer values for the samples and positive controls.
Pharmacokinetic Data Analysis.
Noncompartmental approaches (WinNonlin Pro, version 5.1.2; Pharsight, Mountain View, CA) were used to analyze antibody concentration versus time data in monkeys. Because all of the monkeys developed an ATA response, estimations of clearance and half-life were not possible because of the limited data set and an incomplete characterization of the terminal phase, except for animal 13 in the N434H group, which had a nearly complete concentration-time profile through day 42. The area under the serum concentration-time curve (AUC) from time 0 to day 7 was determined with the linear trapezoidal method, and concentrations at time 0 after an intravenous bolus dose (C0) was estimated by back-extrapolating from the first two concentration values. Pharmacokinetic parameters are reported as mean ± S.D. for n = 6 animals per group. In addition, a two-compartment model with intravenous-bolus input, first-order elimination, and micro-rate constants (model 7) (WinNonlin Pro) was fit to the serum concentration-time data until day 42 for animal 13 in N434H group.
Individual serum concentration versus time data in mice were naive pooled together and analyzed by compartmental approaches to provide an estimation of PK parameters in mice. Intravenous and subcutaneous data for each molecule were fit simultaneously to a two-compartment model to give a good estimation of absorption rate (ka) and bioavailability (F). Mean concentrations were weighted by the inverse of the concentration squared (1/Yhat2, where Yhat is the predicted concentration), and fitting was performed with the use of WinNonlin Pro.
Statistical Data Analysis.
Statistical analysis was performed using JMP (version 6.0; SAS Institute Inc., Cary, NC). PK parameters (AUCdays 0–7 and C0) were analyzed on the logarithmic scale. Tukey's procedure was applied to the log-transformed parameter values to examine the statistical evidence for differences between each group.
Results
Binding Affinity to FcRn.
The binding affinity (KD), as determined by BIAcore, of the anti-TNF-α WT antibody to mouse (mFcRn), cynomolgus monkey (cFcRn), and human FcRn (hFcRn) at pH 6.0 was 12.1, 104, and 165 nM, respectively (Table 1). Compared with that for the WT antibody, the binding affinity of the FcRn variants, N434A and N434H, was higher to mFcRn, cFcRn, and hFcRn at pH 6.0. The N434A and N434H variants showed 1.33- and 1.75-fold improvement in binding to the mFcRn compared with the WT, whereas the binding to cFcRn was ∼7.7- to 23-fold higher than that for the WT. At pH 7.4, the WT antibody had a KD of 3160 nM to mFcRn; the binding affinity to cFcRn and hFcRn could not be quantified, as the binding affinity was below the detection limit of the instrument. The N434A and N434H variants showed ∼2- and 26-fold higher binding affinity than that for the WT antibody to mFcRn at pH 7.4. Of note, the WT humanized anti-TNF-α mAb had higher binding affinity to mFcRn compared with that to hFcRn and cFcRn, which was expected and is consistent with previous reports (Petkova et al., 2006; Datta-Mannan et al., 2007b).
Anti-TNF-α WT and FcRn variant binding affinity to murine, cynomolgus monkey, and human FcRn
Pharmacokinetics in Mice.
After a single 5 mg/kg intravenous or subcutaneous dose in mice of the WT, N434A variant, or N434H variant, the serum concentration-time profiles of WT and N434A were similar (Fig. 1). Thus, there were no differences in the pharmacokinetic parameters between the WT and N434A variant (Table 2). On the other hand, the N434H variant showed faster clearance than the WT or the N434A variant. The estimated clearance (CL) for the N434H variant was approximately 16.1 ml/day/kg, which was ∼3-fold faster than that of the WT (5.07 ml/day/kg) or the N434A variant (5.92 ml/day/kg). The elimination half-life of the N434H variant was ∼2-fold faster and the volume of the central compartment (Vc) appeared slightly (∼48%) higher than that of the WT or the N434A variant. The N434H variant also had a slightly (∼12%) lower bioavailability after subcutaneous administration compared with the WT or the N434A variant (∼61.3 versus ∼73.2 and 72.4% for the WT and N434A, respectively). No difference in the absorption rate was observed among the three antibodies.

Mean (±S.D.) anti-TNF-α mAb serum concentration versus time after a single intravenous (A) or subcutaneous (B) administration of 5 mg/kg WT (●), N434A (▵), or N434H (■) to SCID mice; n = 3 mice/time point.
Two-compartment pharmacokinetic parameter estimates after a single 5 mg/kg i.v. or s.c. dose of anti-TNF-α mAb WT, N434A, and N434H to SCID mice
Pharmacokinetics in Cynomolgus Monkeys.
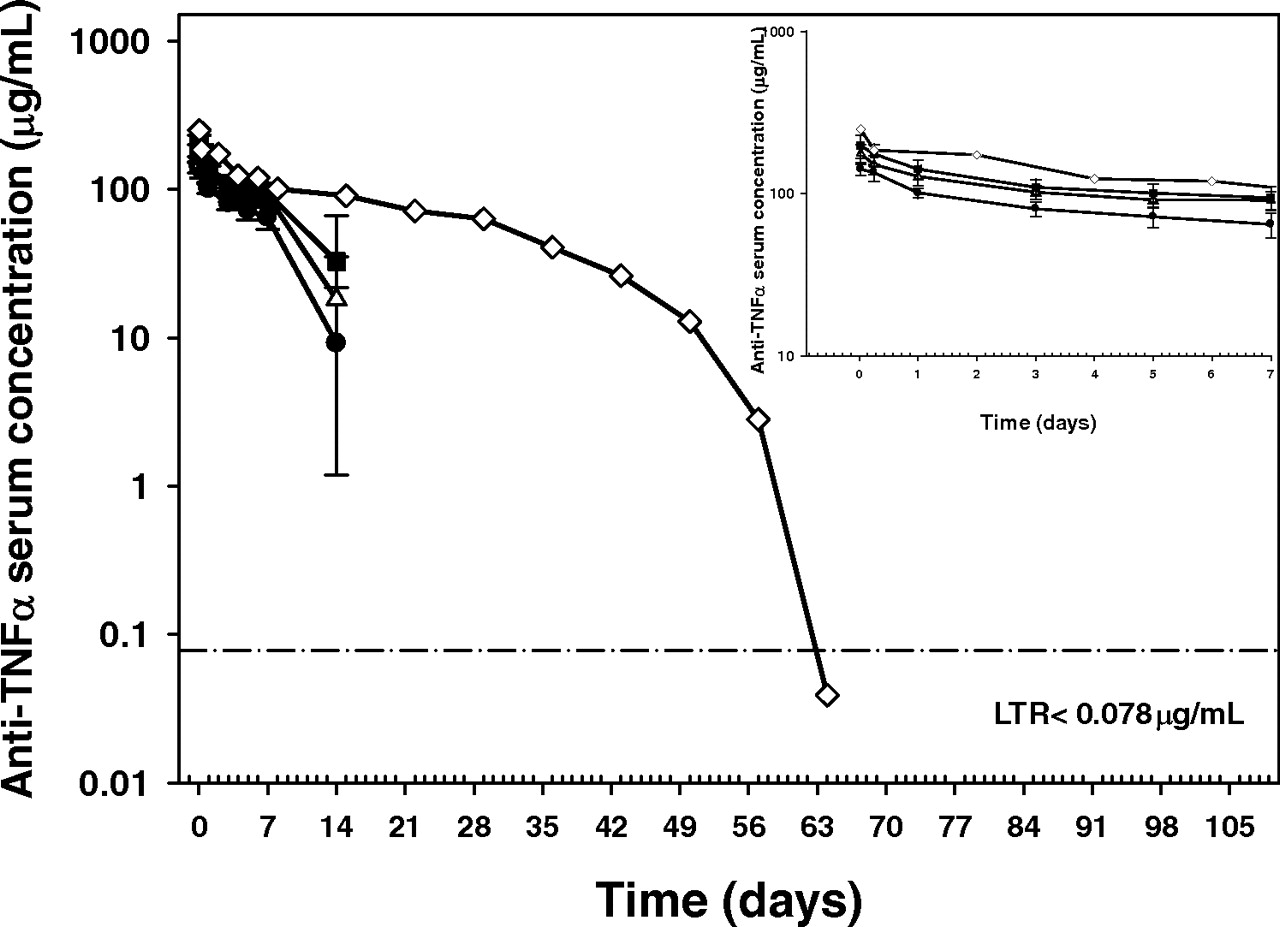
Serum anti-TNF-α mAb concentrations dropped rapidly after day 7 in all animals except animal 13 in the N434H group (Fig. 2). During the initial phase (until day 7), mean serum anti-TNF-α mAb concentrations of N434H were highest, followed by those of N434A and WT in descending order of magnitude. Mean concentration-time profiles for male and female animals were similar in each group (data not shown). All animals in the three groups developed ATAs during the course of the study with the first positive response being observed on day 14 postdose. The reduction in the serum concentrations of the drug after day 7 was probably the result of the development of ATAs. Therefore, serum anti-TNF-α concentrations gathered after day 7 were excluded for the noncompartmental analysis for all animals except for animal 13 that was dosed with N434H. In this animal, serum antibody concentrations declined rapidly after day 42; an ATA response was observed at day 63. Thus, concentration data gathered for animal 13 after day 42 were excluded from the two-compartment analysis.

Mean (±S.D.) serum concentrations of WT (●), N434A (▵), and N434H (■) and concentrations in animal 13 (♢) in the N434H group after a single intravenous dose of 5 mg/kg to cynomolgus monkeys (n = 6 animals/group). Inset, mean (±S.D.) serum concentrations of WT (●), N434A (▵), and N434H (■) and concentrations in animal 13 (♢) in the N434H group up to day 7. Except for the predose time point, less than reportable (LTR) serum concentrations were assigned a value of half the LTR value (1/2 LTR = 0.039 μg/ml) for the first occurrence, and no values for the remaining occurrences, graphic presentation, summary statistics (mean ± S.D.), and PK analysis. If drug concentrations at a PK time point in >30% of the animals in a group were LTR, group means and S.D. for that time point were not calculated. Animal 13 was included when mean data were calculated for N434H group.
The PK parameters calculated using noncompartmental analysis for all animals using data until day 7 are presented in Table 3. The mean AUCdays 0–7 values were 592, 749, and 819 μg/ml · day for WT, N434A, and N434H, respectively. The mean AUCdays 0–7 values for N434A and N434H were 1.27- and 1.38-fold that of the WT, suggesting that N434A and N434H had significantly increased exposure compared with the WT (p < 0.05). The mean estimates of C0 were also slightly higher for the variants compared with that for the WT. The concentration-time profile for animal 13 in the N434H group was described well by a two-compartment model, and the estimated half-life of the β phase (t1/2,β), CL, and AUC were 18.5 days, 1.24 ml/day/kg, and 4019 μg/ml · day, respectively.
Noncompartmental pharmacokinetic parameter estimates after a single 5 mg/kg i.v. dose of anti-TNF-α mAb WT, N434A, or N434H to cynomolgus monkeys
Data are means ± S.D. There is no significant different between the N434A group and N434H group for AUCdays 0–7 and C0.
Discussion
Variants of an anti-TNF-α humanized monoclonal IgG1 antibody, with increased binding to the FcRn, are currently being investigated as therapies with potentially improved efficacy and dosing regimens compared with the available anti-TNF-α therapies for rheumatoid arthritis. The FcRn binding affinity of an IgG appears to correlate with its circulation half-life (Roopenian and Akilesh, 2007). To improve the PK properties of therapeutic antibodies, engineering efforts have been leveraged to modulate the interaction between the Fc portion of the antibody and the FcRn. The present study was designed to evaluate the effects of the binding affinity of an IgG1 mAb to FcRn at different pH values (6.0 and 7.4) on the PK including bioavailability of the mAb. Two FcRn variants with single point substitutions (N434A and N434H) were engineered. The variants showed higher binding affinity to mFcRn, cFcRn, and hFcRn at pH 6.0 but maintained binding affinity comparable to their target, TNF-α (data not shown). The pharmacokinetics of the humanized anti-TNF-α IgG1 mAb WT and these two FcRn variants (N434A and N434H) were evaluated in SCID mice and cynomolgus monkeys.
The pharmacokinetics of the WT and the two FcRn variants in SCID mice were determined after a single intravenous dose. SCID mice were used to minimize the impact of any ATA response on the pharmacokinetic profiles. The pharmacokinetic behavior of the two FcRn variants compared with that of the WT reflected the improved binding affinity at pH 6.0 and the differential binding affinity at pH 6.0 and pH 7.4 to mFcRn. No difference in the PK profiles of the WT and the N434A variant was observed. This result could be attributed to the modest increase in binding affinity at pH 6.0 and, more importantly, to the increased binding affinity at pH 7.4 compared with that of the WT. N434A had a KD of 9.1 nM to mFcRn at pH 6.0, which is 1.33-fold of the WT. However, the KD at pH 7.4 was 1594 nM to mFcRn, which is 1.97-fold of the WT. Hence, PK benefits from the modest increases in binding affinity at pH 6.0 of these FcRn variants were compensated for by the simultaneous undesirable increase in binding affinity at pH 7.4. On the other hand, the half-life of the N434H variant was reduced to approximately half of that of the WT (6.60 days versus 11.6 days). The N434H variant had ∼1.75-fold increased binding affinity to mFcRn at pH 6.0, but it had a ∼25-fold increased binding affinity at pH 7.4 compared with the WT. Thus, the increased binding affinity at pH 7.4 probably resulted in a faster clearance of this variant compared with the WT. The high binding affinity at pH 7.4 has been suggested to impede the release of FcRn-bound IgG variant back into the circulation and also possibly to allow circulating IgG variants to bind directly to the FcRn expressed on the cell surface, effectively lowering their serum concentrations. Therefore, the more dramatic increase in FcRn binding affinity at pH 7.4 had a dominant-negative effect on IgG clearance.
In cynomolgus monkeys, after intravenous administration of the anti-TNF-α mAbs, WT, and the two FcRn variants, the serum mAb concentrations declined rapidly after day 7 in all except one animal (animal 13 in N434H group). Given the low expression levels of TNF-α in animals, target-mediated clearance was not expected to significantly contribute to overall mAb clearance at a dose of 5 mg/kg for any of the anti-TNF-α mAbs. All animals developed ATAs during the course of the study. The first time of detection of ATAs ranged from day 14 to day 28 for most animals. In general, the detection of ATAs appeared to be associated with a rapid decline in serum concentration. Therefore, the pharmacokinetics of WT, N434A, and N434H variants were compared by examining serum concentrations through day 7 to mitigate the potential effects of ATA on CL. CL could not be estimated with certainty because of the ATA response; thus, AUCdays 0–7 values were used for PK comparisons. The average AUCdays 0–7 for N434A and N434H was 1.27- and 1.38-fold that of WT (749 and 819 versus 592 μg/ml · day, respectively), indicating that N434A and N434H had significantly increased exposure from day 0 to day 7 compared with the WT (p < 0.05). Animal 13 in the N434H group had a nearly complete concentration-time profile through day 42 in the absence of an ATA response, which provided an opportunity to estimate the CL and half-life with confidence. The estimated half-life for this animal was approximately 18.5 days, which is longer by a factor of ∼2-fold than that of typical WT IgG1 in monkeys (Benincosa et al., 2000; Kakkar et al., 2007) and is consistent with our conclusion based on the AUCdays 0–7. These results are consistent with the binding affinity of these monoclonal antibodies to the cFcRn, in which the WT had the lowest binding affinity to the cFcRn at pH 6.0. No significant differences in AUCdays 0–7 were observed between the two variant groups, although N434H had ∼3-fold higher affinity at pH 6.0 than N434A. The possible explanations for this observation are as follows: 1) a 3-fold improvement in the binding affinity does not translate into improved pharmacokinetic properties; 2) the two FcRn variants would have demonstrated different PK behavior in the terminal elimination phase in the absence of ATAs; and 3) there may be a maximum achievable improvement in pharmacokinetics for variant antibodies with increased FcRn binding affinity at pH 6.0. It has been reported that some FcRn variants with ∼3 and ∼30-fold increased binding affinity (pH 6.0) have similar increases in half-life compared with the WT in monkeys (Hinton et al., 2004).
We observed increased exposure in monkeys for our FcRn variants by increasing the binding affinity at pH 6.0 while keeping the pH dependence (i.e., no or low binding at pH 7.4), which is consistent with previous reports (Dall'Acqua et al., 2002, 2006; Hinton et al., 2004, 2006; Petkova et al., 2006; Yeung et al., 2009). However, Gurbaxani et al. (2006) did not identify a single FcRn binding parameter that directly correlated with antibody elimination half-life. Likewise, Datta-Mannan et al. (2007a,b) did not observe a direct relationship between increased binding affinity to the FcRn and improved PK properties for the FcRn variants of their anti-TNF-α mAb in monkeys. The CL in monkeys for the WT molecule in the studies of Datta-Mannan et al. (2007a,b) is ∼26 ml/day/kg. This reported CL is much faster than that of a typical IgG1 antibody with low endogenous target levels in monkeys, which generally ranges from ∼5 to ∼10 ml/day/kg at doses of 0.5 to 300 mg/kg (Benincosa et al., 2000; Kakkar et al., 2007). Hence, it is possible that the IgG1 in the reports of Datta-Mannan et al. (2007a,b) may have additional clearance mechanisms compared with those of other IgG1s (e.g., proteolysis and nonspecific binding, given the low dose of 0.5 mg/kg). Therefore, modification of the binding affinity to FcRn may not yield a change in the in vivo fate of this IgG1 because it is possible that the FcRn-mediated clearance pathway may not be predominant. Thus, a number of factors need to be considered to better understand the effects of the FcRn binding affinity on the mAb PK: the difference in the absolute IgG-FcRn affinity at pH 6.0 and pH 7.4, the kinetics of the IgG/FcRn interaction at pH 6.0 as well as at pH 7.4, and the relative contribution of the FcRn-mediated clearance pathway to the total clearance of mAb compared with that of other clearance mechanisms.
An FcRn variant with stronger affinity to FcRn at pH 7.4 (N434H) had a slightly lower bioavailability (F) in mice after subcutaneous administration compared with that of the WT or the N434A variant. Bioavailability for N434H was ∼61%, whereas N434A and WT have bioavailability of ∼73%. After subcutaneous administration, IgG reaches the systemic circulation by lymphatic drainage as well as by convection and probably undergoes presystemic metabolism. Because the skin and the muscle are major expression organs of FcRn (Roopenian and Akilesh, 2007), FcRn may play a protective role for IgG in the presystemic metabolism. Thus, the decrease in the systemic availability of N434H may be due to the increase in binding affinity to FcRn at pH 7.4, which may delay the release of the antibody and accelerate the degradation rate of IgG locally. Currently, the bioavailability for mAbs after subcutaneous administration in humans is approximately 60% [64% for adalimumab (Humira; prescribing information from Abbott Laboratories, Chicago, IL, 2007) and 62% for omalizumab (Xolair; prescribing information from Genentech Inc. and Novartis, East Hanover, NJ, 2006)]. FcRn variants with higher binding affinity at pH 6.0 but not at pH 7.4 may provide a new approach to increase the bioavailability for mAb therapeutic agents. However, more investigation needs to be done to understand the role of FcRn in absorption of antibodies after subcutaneous administration.
We have shown that FcRn variants of our anti-TNF-α mAb have increased affinity to human FcRn at pH 6.0 while keeping a low binding affinity at pH 7.4. Because our WT anti-TNF-α mAb has a PK profile similar to that of adalimumab in monkeys (Loyet et al., 2009) and mice (data not shown), our FcRn variant may have improved PK profiles in humans as well (i.e., improved half-life). The trough concentration for adalimumab after subcutaneous dosing at 40 mg every other week treatment is ∼5 μg/ml (Abbott Laboratories, 2007). This concentration should be achieved by the anti-TNF-α FcRn variants described here at a 75-mg monthly treatment, with the assumptions that anti-TNF-α FcRn variants have an ∼2-fold improved half-life in humans.
In summary, we have observed significant increases in the exposure of the two anti-TNF-α FcRn variants in cynomolgus monkeys by enhancing the binding affinity of the mAb to the cFcRn at pH 6.0 while keeping the binding affinity at pH 7.4 low. However, increasing the binding affinity at pH 7.4 to mFcRn resulted in faster clearance in mice and possibly decreased the subcutaneous bioavailability in mice compared with that of the WT despite an increase in affinity at pH 6.0. Engineering mAbs with a desirable binding affinity to FcRn at pH 6.0 as well as pH 7.4 is a promising strategy to improve the pharmacokinetics of therapeutic mAbs.
Acknowledgments.
We thank Wei-ching Liang and Yan Wu for engineering the antibodies, Noore Kadri for conducting the mouse PK study, Ly Nguyen Kawaguchi for coordinating the monkey PK study, and Eric Stefanich, Lisa Damico, Jay Tibbitts, and Meina Tang for review of the manuscript.
Footnotes
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.109.031310.
-
ABBREVIATIONS:
- FcRn
- neonatal Fc receptor
- β2m
- β2-macroglobulin
- WT
- wild type
- mAb
- monoclonal antibody
- TNF
- tumor necrosis factor
- SCID
- severe combined immunodeficiency
- PK
- pharmacokinetic(s)
- ATA
- anti-therapeutic antibody
- AUC
- area under the serum concentration-time curve
- CL
- clearance
- mFcRn
- murine FcRn
- cFcRn
- cynomolgus monkey FcRn
- hFcRn
- human FcRn.
- Received November 23, 2009.
- Accepted January 13, 2010.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}