


Abstract
Ipratropium bromide (ipratropium) and tiotropium bromide (tiotropium), anticholinergic agents with bronchodilating properties, are used to treat patients with chronic obstructive pulmonary disease. Because they are actively secreted into urine, the interaction of these agents with organic cation transporters (OCTs/Octs) was examined in rat kidney slices and in cultured cells expressing rat Oct (rOct) or human OCT (hOCT). Uptake of radiolabeled ipratropium in rat kidney slices was significantly inhibited by OCT/Oct substrates including cimetidine, imipramine, and quinidine, but not by organic anion transporter substrates (e.g., p-aminohippuric acid and estrone-3-sulfate). [3H]Tiotropium uptake showed similar characteristics. Reverse transcription-polymerase chain reaction showed that, in rat kidney, mRNA expression of rOct2 was the highest, followed by rOct1, but little rOct3 was detected. In vitro, rOct1 and rOct2 transported both anticholinergics, but rOct3 accepted only ipratropium. Ipratropium uptake by rat kidney slices consisted of two components with Km values of 0.114 ± 0.06 and 24.5 ± 2.21 μM. The Km value of rOct2-mediated ipratropium uptake (0.143 ± 0.03 μM) was consistent with that of the high-affinity component. The OCT/Oct inhibitor corticosterone, at a concentration of 1 μM (IC50, 1.11 ± 0.20 μM for rOct2-mediated ipratropium transport), inhibited ipratropium by 18.4%, suggesting that rOct2 is involved in renal secretion of ipratropium. In a similar manner, ipratropium and tiotropium were taken up by cultured cells expressing hOCT1 and hOCT2 but not hOCT3. We conclude that OCT2/Oct2 plays a role in renal secretion of both anticholinergics in these species. Coadministration of these anticholinergics with cationic drugs recognized by OCT2/Oct2 may decrease renal clearance, resulting in increased systemic exposure.
Introduction
Chronic obstructive pulmonary disease (COPD), also known as chronic obstructive airways disease, is a term for a group of lung diseases (including chronic bronchitis and emphysema) in which the main symptom is difficulty in breathing; it is currently one of the most common respiratory diseases (Koumis and Samuel, 2005). For the treatment of COPD, inhaled anticholinergic drugs such as ipratropium bromide (ipratropium) and tiotropium bromide (tiotropium) are effective in expanding the bronchial airways by antagonizing the action of acetylcholine at muscarinic receptor 1 to 3 (M1–3) in bronchial smooth muscle (Gross and Skorodin, 1984). Tiotropium is the first of a new generation of anticholinergics with a higher affinity for M1 and M3 than M2, resulting in long-lasting bronchodilating action (Disse et al., 1993, 1999).
Although both ipratropium and tiotropium are highly hydrophilic quaternary ammonium compounds chemically related to atropine and unlikely to cross the plasma membranes by diffusion, it is known that they possess relatively high renal clearances. Early pharmacokinetic studies indicated that renal excretion of ipratropium amounted to 58 and 55% after a single intravenous administration in rats and dogs, respectively (Forester et al., 1976). In humans, the renal clearances of both ipratropium (872 ml/min) and tiotropium (435 ml/min) are much greater than the glomerular filtration rate (Ensing et al., 1989; Türck et al., 2004). We and others have recently suggested an interaction of these anticholinergics with organic cation transporters. We have shown that both ipratropium and tiotropium are recognized by human organic cation/carnitine transporter 2 (hOCTN2) and, to lesser extent, hOCTN1 (Nakamura et al., 2010). Ipratropium was also reported to inhibit polyspecific organic cation transporter 2 (OCT2)-mediated l-methyl-4-phenylpyridinium (MPP+) transport (Zolk et al., 2009).
These transporters are expressed in tubular cells, which are potential sites of drug-drug interactions leading to retarded tubular secretion and reabsorption, which could contribute to cellular accumulation and urinary excretion of numerous compounds (Pritchard and Miller, 1993). OCTNs are thought to mediate multispecific and bidirectional transport of organic cations, as well as carnitine, at the apical membranes of proximal tubular cells (Tamai et al., 1998, 2000; Ohashi et al., 1999, 2001; Yabuuchi et al., 1999;). However, among the three subtypes, OCT2 is predominantly expressed in the basolateral membranes of proximal tubular cells in humans (Gorboulev et al., 1997) and has been suggested to play a critical role in the renal secretory processes of organic cationic drugs through proximal tubular cells (Suhre et al., 2005). To date, the secretory processes of ipratropium and tiotropium have not been clarified, although in vivo pharmacokinetics studies have suggested that they are actively secreted into urine. Therefore, we investigated the role of OCTs/Octs in renal uptake of both ipratropium and tiotropium, using rat kidney slices and HEK293 cells expressing rat or human OCTs (rOcts or hOCTs, respectively). In this study, we present the first evidence that OCTs/Octs recognize anticholinergic drugs as substrates and the role they may play in renal secretion. This study provides pharmacologically important information to predict alteration in the pharmacokinetics of these anticholinergics due to transporter-mediated drug-drug interaction during renal secretion.
Materials and Methods
Chemicals.
3H-Labeled and unlabeled ipratropium bromide (72 Ci/mmol) and tiotropium bromide (81 Ci/mmol) were provided by GlaxoSmithKline (Uxbridge, Middlesex, UK). [3H]MPP+ (80 Ci/mmol) was purchased from American Radiolabeled Chemicals (St. Louis, MO). Corticosterone, TEA+ chloride, and quinidine sulfate were obtained from Sigma-Aldrich (St. Louis, MO) and l-carnitine was obtained from Wako Pure Chemicals (Osaka, Japan), respectively. All other chemicals and reagents were commercial products of reagent grade.
Cell Culture.
HEK293 cells expressing rOct1, rOct2, rOct3, hOCT1, hOCT2, hOCT3, and plasmid vector alone were prepared in our laboratory in the similar method as described previously (Tamai et al., 2001). In brief, HEK293 cells were transfected with the respective plasmid DNA, and then selected with proper antibiotics; they were designated HEK293/rOct1, HEK293/rOct2, HEK293/rOct3, HEK293/hOCT1, HEK293/hOCT2, HEK293/hOCT3, and mock cells, respectively. All of the cell lines were grown in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA) supplemented with 10% (v/v) fetal bovine serum (Invitrogen), 100 units/ml penicillin, and 100 μg/ml streptomycin at 37°C in an atmosphere of 5% CO2.
Uptake of [3H]Ipratropium and [3H]Tiotropium by Rat Kidney Slices.
Kidneys slices (0.3-mm thick) from 8-week-old male Wistar rats were cut with a microslicer (Zero 1; Dosaka, Kyoto, Japan). Apparent uptake of [14C]inulin (0.05 μCi/ml; American Radiolabeled Chemicals) by the slices was measured to evaluate the volume of water adhering to the surface of the kidney slices, as described previously (Nakakariya et al., 2009). The saturable component of [3H]ipratropium (10 nM) uptake by the slices was determined by subtracting uptake obtained in the presence of 2 mM ipratropium from the observed uptake.
Uptake of [3H]Ipratropium and [3H]Tiotropium by HEK293 Cells.
HEK293 cells expressing a transporter gene were plated in a 24-well tissue culture plate at a density of 1 × 105 cells/cm2. Cells were cultured for 2 days and then used for uptake experiments. In general, an uptake experiment was initiated by adding 0.25 ml of transport buffer (125 mM NaCl, 4.8 mM KCl, 5.6 mM d-glucose, 1.2 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 25 mM HEPES, adjusted to pH 7.4) containing radiolabeled substrate to the cells. At the end of the uptake reaction, cells were washed with ice-cold transport buffer twice and solubilized in 0.2 ml of 1% (v/v) Triton X-100 (Wako Pure Chemicals). The radioactivity in the lysate was measured using a liquid scintillation counter (Aloka, Tokyo, Japan). Part of the lysate was used for determination of total protein amount with a protein assay kit (Bio-Rad Laboratories, Hercules, CA). Uptake of substrate by mock cells was subtracted from that by HEK293 cells expressing transporter gene to evaluate transport activity.
Quantification of mRNA Expression by Quantitative Reverse Transcription-Polymerase Chain Reaction.
mRNA expression of Oct1, Oct2, and Oct3 in rat kidney was measured by quantitative reverse transcription-polymerase chain reaction (qRT-PCR). Total RNA was prepared using Isogen reagent (Nippon Gene, Tokyo, Japan) from freshly isolated rat kidney after perfusion with phosphate-buffered saline. RT-PCR was performed using gene-specific primers as follows: 5′-TTTAACCTGGTGTGTGGAGACG-3′ (sense) and 5′-AGGAAGAAGCCCAAGTTCACAC-3′ (antisense) for rOct1, 5′-CGGTGCTATGATGATTGGCTAC-3′ (sense) and 5′-CCAGGCATAGTTGGGAGAAATC-3′ (antisense) for rOct2, and 5′-ATATCCTGTTTCGGCGTTGG-3′ (sense) and 5′-TTTCCAAACACCCCTTGCAG-3′ (antisense) for rOct3. RT-PCR was performed in the presence of Brilliant SYBR Green QPCR Master Mix (Agilent Technologies, Santa Clara, CA). The copy number of mRNA for the respective gene was determined from a standard curve based on the fluorescence of known amounts of the plasmid DNA containing cDNA of the gene. Each value obtained from qRT-PCR was normalized by mRNA expression of glyceraldehyde-3-phosphate dehydrogenase. The fold change in the OCT transporter gene, normalized to hypoxanthine-guaninephosphoribosyl-transferase (HPRT) and relative to the expression in mock cells, was analyzed by qRT-PCR and calculated using 2−ΔΔCT methods (Livak and Schmittgen, 2001). Primers for HPRT were prepared as described previously (Fischer et al., 2005).
Analytical Methods.
The cell-to-medium ratio (microliters per milligram of protein), which represents the volume of cleared extracellular transport medium, was obtained by dividing the intracellular accumulation of the test compound by its initial concentration in the transport medium applied. All data were expressed as the means ± S.E.M., and statistical analysis was performed by applying the Student's t test with the criterion of p < 0.05 for significance. The apparent kinetic parameters, Km (Michaelis constant), Vmax (maximal uptake rate), and kd (apparent nonsaturable first-order rate constant), of ipratropium uptake by rat kidney slices and HEK293 cells were calculated by nonlinear least-squares regression analysis using KaleidaGraph 4.0.2 (Synergy Software, Reading, PA) according to the Michaelis-Menten type (eqs. 1 and 2, respectively), where v and s are the uptake rates of substrate and substrate concentration, respectively.


Results
[3H]Ipratropium Uptake by Rat Kidney Slices.
We initially examined [3H]ipratropium uptake by rat kidney tissues. As shown in Fig. 1, apparent [3H]ipratropium uptake increased in a time-dependent manner and reached a plateau in 10 min. To determine whether carrier-mediated transport is involved in the renal uptake of ipratropium, various cationic and anionic compounds were tested for their inhibitory effects on ipratropium uptake (Table 1). [3H]Ipratropium uptake for 1 min was significantly decreased in the presence of an excess amount (1 mM) of unlabeled ipratropium or OCT/Oct substrates, including cimetidine, quinidine, imipramine, MPP+, and TEA+, but not anionic compounds such as p-aminohippuric acid and TEA+-3-sulfate, which are well established substrates for organic anion transporter and organic anion transporter polypeptide, respectively (Table 1). Similar observations were made for [3H]tiotropium uptake by rat kidney slices in a competitive inhibition study (Table 1). mRNA expression of Octs was further examined in rat kidney. mRNA expression of rOct2 was the highest among the three subtypes, and the level of rOct1 was 53% of that of rOct2, whereas the expression level of rOct3 mRNA was negligible (Fig. 2). Therefore, these results suggest that rOct1 and rOct2 are involved in renal uptake of both ipratropium and tiotropium.

Time course of [3H]ipratropium uptake by rat kidney slices. Rat kidney slices were incubated with [3H]ipratropium (10 nM) at 37°C and pH 7.4 for 30 min. The apparent uptake of [14C]inulin was measured for each time point to estimate the volume of water adhering to the kidney slices during the incubation. Each point represents the mean ± S.E.M. of at least four slices from three individual experiments.
Inhibitory effect of organic cations and anions on uptake of ipratropium and tiotropium by rat kidney slices
[3H]Ipratropium (10 nM) uptake and [3H]ipratropium (10 nM) uptake by rat kidney slices were determined at 37°C and pH 7.4 for 1 min in the absence (control) and presence of an indicated inhibitor at the concentration of 1 mM. Each point represents the mean ± S.E.M. of four slices.

The expression levels of Oct mRNAs in rat kidney were determined by qRT-PCR. Each expression level was normalized by the mRNA expression of glyceraldehyde-3-phosphate dehydrogenase (Gapdh), as described under Materials and Methods. Each bar represents the mean ± S.E.M. (n = 4).
Uptake of [3H]Ipratropium and [3H]Tiotropium by HEK293 Cells Expressing rOcts.
To identify the rOct subtype responsible for the renal uptake of both ipratropium and tiotropium, the uptake rates of these compounds were examined in HEK293/rOct1, HEK293/rOct2, and HEK293/rOct3cells (Fig. 3, A–C). First, the overexpression level of Oct transporter mRNA expression in HEK293/rOct1, HEK293/rOct2, and HEK293/rOct3 normalized to HPRT was calculated at 1.486 ± 0.075, 0.604 ± 0.029, and 0.790 ± 0.050 (copy/copy of HPRT, n = 8), respectively. The initial uptake rates of [3H]ipratropiumin at 1 min were 8.7-, 34-, and 5.1-fold greater in HEK293/rOct1, HEK293/rOct2, and HEK293/rOct3 cells than in mock cells, respectively. In a similar manner, [3H]tiotropium uptake was 8.4- and 3.0-fold greater in HEK293/rOct1 and HEK293/rOct2, respectively, but was unchanged in HEK293/rOct3 compared to mock cells. Ipratropium uptake by HEK293/rOct2 cells was approximately 4.5-fold greater than that of MPP+, a well established substrate for OCTs, showing that ipratropium is preferentially transported via rOct2. We further studied rOct2- and rOct1-mediated [3H]ipratropium transport and characterized them by evaluating time- and concentration-dependent uptake in HEK293/rOct1 and HEK293/rOct2cells (Fig. 4). [3H]Ipratropium uptake by HEK293/rOct1 and HEK293/rOct2 cells increased in a time-dependent manner and reached a plateau within 15 and 2 min, respectively (Fig. 4, A and B). Thus, we measured the initial uptake rate at 3 min and 30 s to examine the concentration dependence of rOct1- and rOct2-mediated ipratropium uptake, respectively. Based on the saturation kinetics of ipratropium uptake, the Km and Vmax values for rOct1 and rOct2 were estimated to be 36.9 ± 7.60 μM and 223 ± 19.5 pmol/3 min · mg protein (Fig. 4C) and 0.143 ± 0.03 μM and 7.33 ± 0.36 pmol/30 s · mg protein (Fig. 4D), respectively, based on eq. 1.

Uptake of ipratropium (Ipra) and tiotropium (Tio) by rOcts. Uptake of both [3H]ipratropium (10 nM) and [3H]tiotropium (10 nM) by HEK293/rOct1 (A, closed bars), HEK293/rOct2 (B, closed bars), HEK293/rOct3 (C, closed bars), and mock cells (open bars) was measured at 37°C and pH 7.4 for 1 min. Each bar represents the mean ± S.E.M. (n = 4).
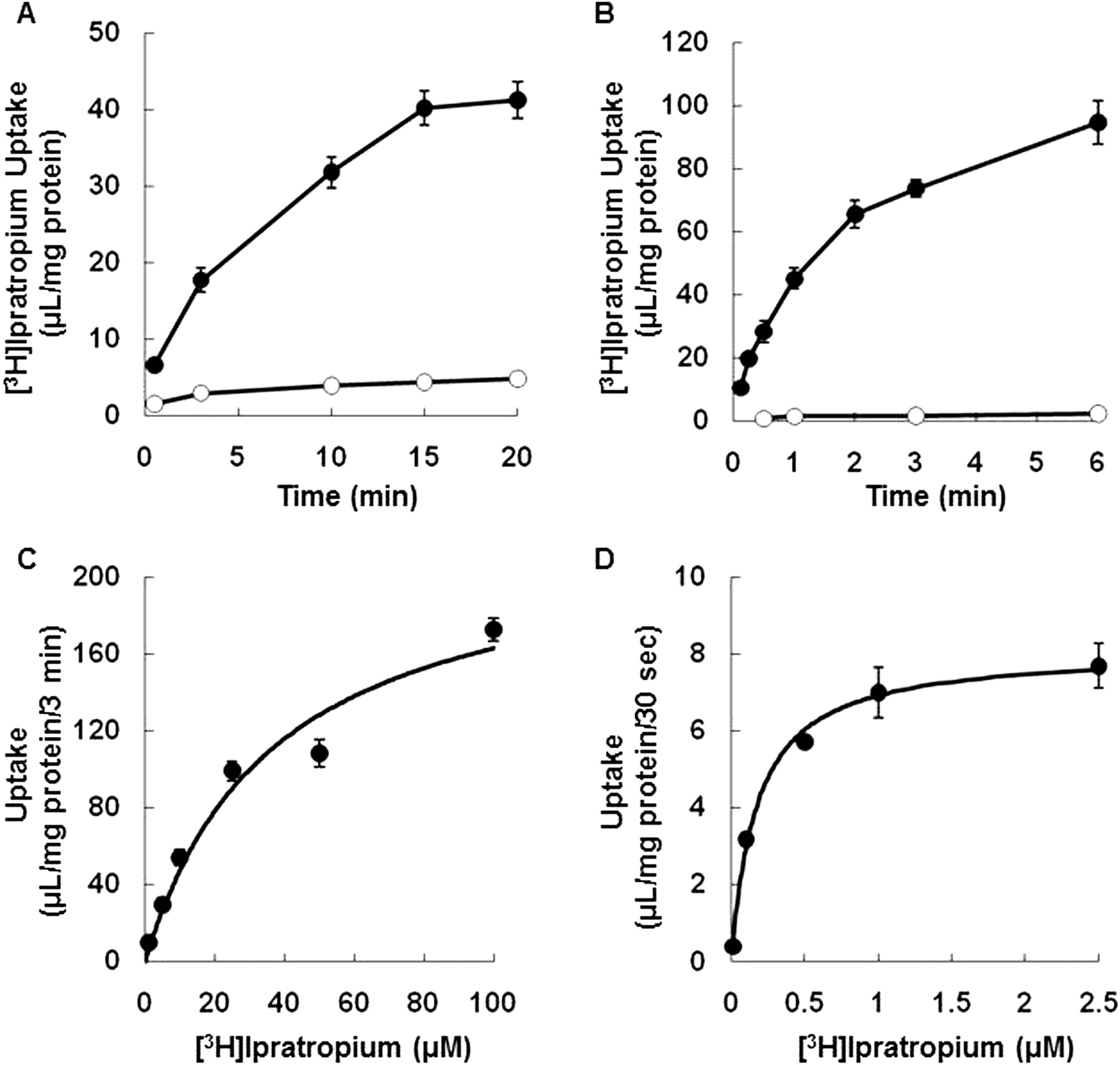
Time course and concentration dependence of ipratropium uptake by rOct1 and rOct2. Uptake of [3H]ipratropium (10 nM) by HEK293/rOct1 (A and C, closed circles), HEK293/rOct2 (B and D, closed circles), and mock (open circles) was measured at 37°C and pH 7.4 for 3 min (A and B) and 30 s (C and D). Transporter-mediated uptake of ipratropium was determined by subtraction of the uptake by mock cells. Each point represents the mean ± S.E.M. (n = 4).
Concentration Dependence of [3H]Ipratropium Uptake by Rat Kidney Slices.
To determine the subtype of Octs responsible for ipratropium uptake in rat kidney, the Km value for [3H]ipratropium uptake by kidney slices was obtained from the saturation kinetics. The concentration dependence of ipratropium uptake was measured in the range of 1 nM to 100 μM (Fig. 5, inset). The uptake rate of ipratropium was fitted to a model containing two saturable components, as described under Materials and Methods (eq. 2); the Eadie-Hofstee plot is shown in Fig. 5. Based on the fitting, the Km values for the high- and low-affinity components were 0.114 ± 0.06 and 24.5 ± 2.21 μM, respectively. The Km for the high-affinity component was almost identical to the Km for rOct2-mediated ipratropium uptake obtained from HEK293/rOct2 cells (Fig. 4D).

Eadie-Hofstee plots of the uptake of ipratropium by rat kidney slices. [3H]Ipratropium uptake by rat kidney slices was measured over the concentration range of 1 nM to 100 μM for 1 min at 37°C and pH 7.4. Transporter-mediated uptake was determined by subtracting the uptake in the presence of 2 mM ipratropium from the uptake at each concentration of ipratropium. The inset shows the concentration dependence of [3H]ipratropium uptake expressed according to the Michaelis-Menten equation. Each point represents the means ± S.E.M. (n = 4).
Inhibitory Effects of Corticosterone on rOct1- and rOct2-Mediated Uptake by HEK293 Cells.
To distinguish the contributions of rOct2 and rOct1 to ipratropium uptake, we used corticosterone, which is an inhibitor with different affinity for the two transporters. First, the affinity of corticosterone for ipratropium uptake by HEK293/rOct1 and HEK293/rOct2 cells was measured. The IC50 values of corticosterone were estimated to be 55.8 ± 7.68 and 1.11 ± 0.197 μM, respectively (Fig. 6, A and B). The data on the inhibitory effect of corticosterone are summarized in Table 2. Corticosterone at the concentration of 1 μM inhibited rOct2-mediated uptake by 47% but did not inhibit rOct1 at all, i.e., 1 μM corticosterone is a selective inhibitor of rOct2. As shown in Table 2, in the presence of 1 μM corticosterone, ipratropium uptake by rat kidney slices was inhibited by 18.4%, showing that rOct2 contributes to ipratropium uptake. Although corticosterone at the concentration of 500 μM completely inhibited ipratropium uptake by HEK293/rOct1 and HEK293/rOct2 cells, in the case of rat kidney slices, approximately 30% of the uptake remained. In accordance with this result, it appears that ipratropium is also transported by corticosterone-insensitive transporters in the slices.

Corticosterone inhibition of ipratropium uptake by HEK293/rOct1 and HEK293/rOct2 cells. Uptake of [3H]ipratropium (10 nM) by HEK293/rOct1 (A) and HEK293/rOct2 (B) cells was measured in the absence or the presence of corticosterone in the concentration range of 1 to 100 μM for 30 s at 37°C and pH 7.4. Transporter-mediated uptake was determined by subtracting the uptake by mock cells from that by rOct1- or rOct2-expressing cells, and the uptake rate was normalized by the value obtained in the absence of corticosterone in each assay. Each point represents the means ± S.E.M. (n = 4).
Inhibitory effect of corticosterone on ipratropium uptake by rat kidney slices
[3H]Ipratropium (10 nM) uptake by rat kidney slices was determined at 37°C and pH 7.4 for 1 min in the absence (control) and presence of an indicated inhibitor at the concentration of 1 mM. Each point represents the mean ± S.E.M. of four slices.
Uptake of [3H]Ipratropium and [3H]Tiotropium by HEK293 Cells Expressing Human OCTs.
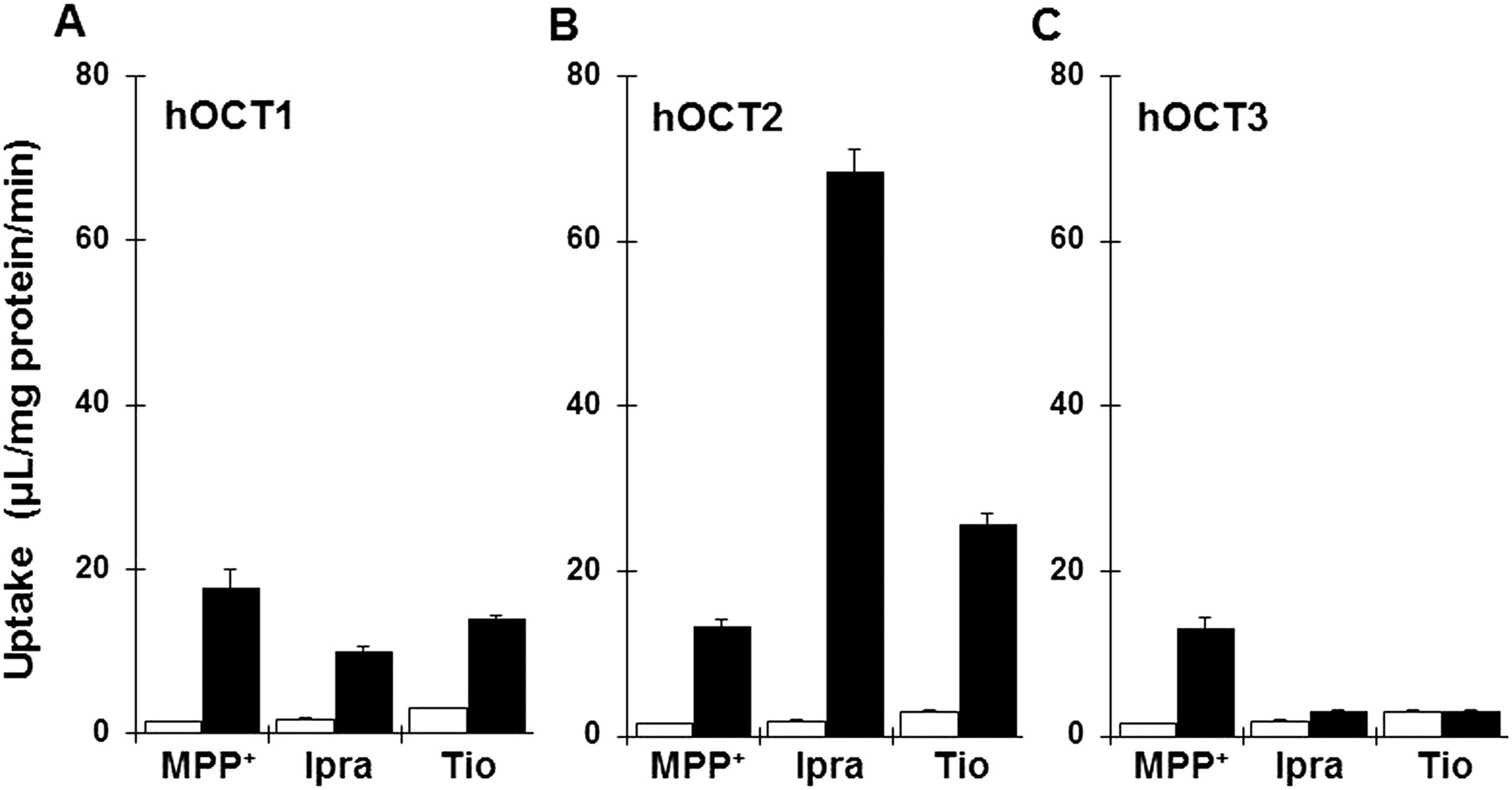
To determine whether ipratropium and tiotropium are recognized by hOCTs, we examined uptake of ipratropium and tiotropium by HEK293/hOCT1, HEK293/hOCT2, or HEK293/hOCT3 (Fig. 7, A–C). The fold change in each transporter gene expression normalized to HPRT and relative to mock cells is shown in Supplemental Fig. 1 to confirm overexpression of transporter, respectively. Uptake of [3H]ipratropium and [3H]tiotropium by HEK293 cells expressing hOCT1 and hOCT2 was 5.7- and 39-fold and 4.6- and 8.5-fold greater, respectively, than that by mock-transfected cells, whereas hOCT3 showed little transport activity for these anticholinergics.
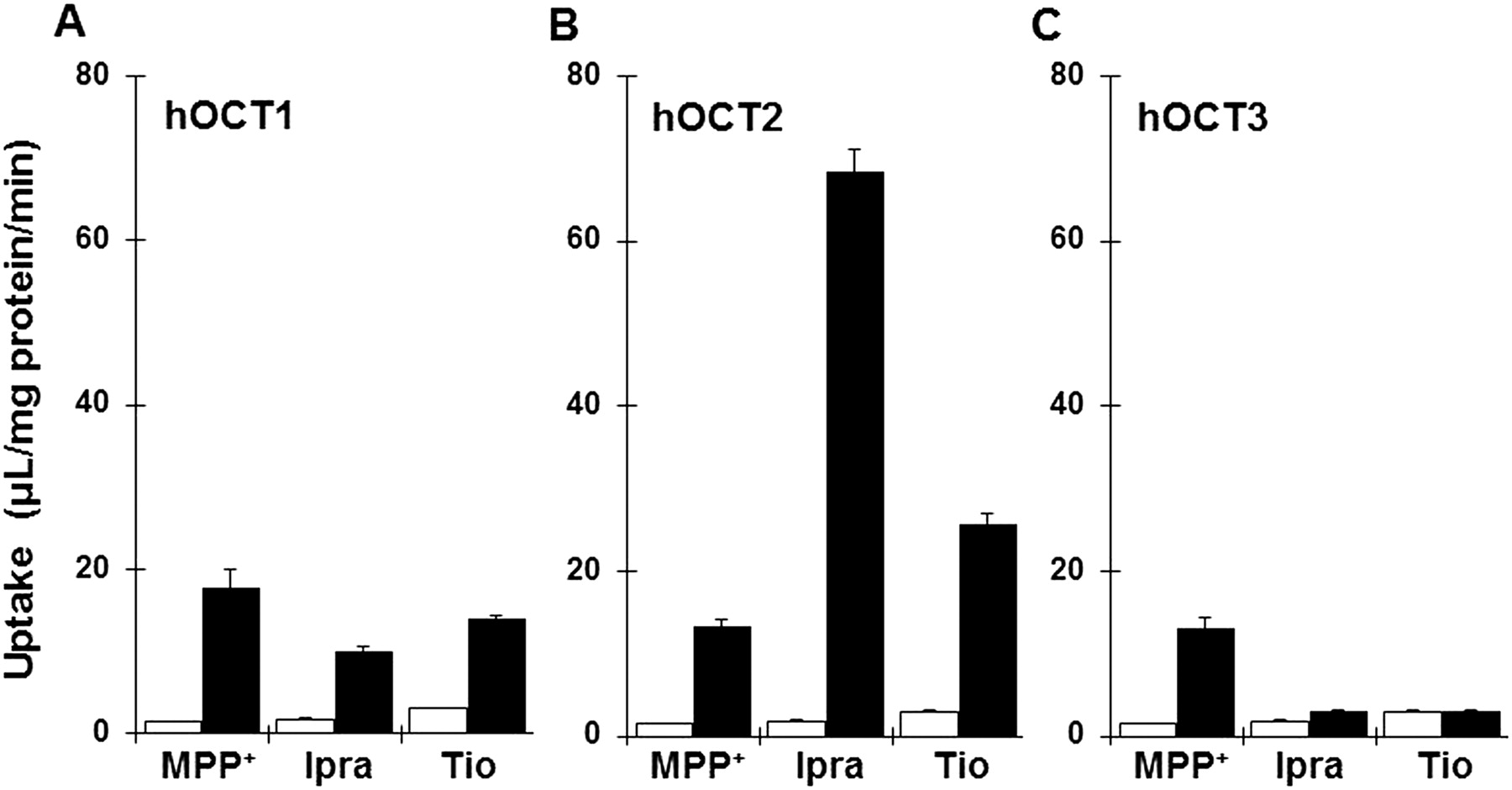
Uptake of ipratropium (Ipra) and tiotropium (Tio) by hOCTs. Uptake of both [3H]ipratropium (10 nM) and [3H]tiotropium (10 nM) uptake by HEK293/hOCT1 (A, closed bars), HEK293/hOCT2 (B, closed bars), HEK293/hOCT3 (C, closed bars), and mock cells (open bars) were measured at pH 7.4 and 37°C for 1 min. Each bar represents the mean ± S.E.M. (n = 4).
Discussion
In this study, we explored the molecular mechanism of renal secretion of two anticholinergics, ipratropium and tiotropium, showing for the first time that they are transported by human and rat OCTs/Octs. Our data provide a new rationale for the relatively high renal clearance of these anticholinergics.
Kidney slices are a well established model for evaluating absorptive transport into proximal tubular cells and have been used to characterize the interaction of various organic cations with Octs expressed in the basolateral membranes of rat tubular cells (Ishiguro et al., 2005; Dantzler, 2006). OCTs facilitate the diffusion of structurally diverse organic cations including endogenous neurotransmitters, catecholamines, and many drugs in clinical use. [3H]Ipratropium uptake by rat kidney slices was significantly decreased in the presence of other OCT/Oct substrates including cimetidine, imipramine, and quinidine, but not substrates of organic anion transporters (Table 1). Our previous report showed that both OCTN2 and, to a lesser extent, OCTN1 transport ipratropium (Nakamura et al., 2010), and mouse Octn2 is localized at the apical membranes of proximal tubular cells (Tamai et al., 2001); therefore, it is thought that rat Octn2 was not involved in the apparent uptake by kidney slices. The results of the RT-PCR study were in a good agreement with a previous report showing greater expression of rOct2 and, to a lesser extent, rOct1 compared with rOct3 (Fig. 2). In vitro uptake experiments using OCT/Oct-expressing HEK293 cells demonstrated that both ipratropium and tiotropium are substrates of both rat and human OCT1/Oct1 and OCT2/Oct2 (Figs. 3 and 7). Therefore, absorption of these anticholinergics into rat proximal tubular cells mediated by rOct1 and rOct2 is likely to be the first step in their renal secretion. We have reported that both anticholinergics are transported by OCTN2, which is expressed in the apical membranes of proximal tubular cells, and the OCTN2-mediated ipratropium occurred in an Na+-independent manner (Nakamura et al., 2010) so that ipratropium may be bidirectionally transported via OCTN2/Octn2 as shown for TEA (Ohashi et al., 2001). If so, high renal clearance of these anticholinergics could be due to concerted action of OCTs with OCTNs to mediate a vectorial movement through the cells. Multidrug and toxic compound extrusion transporters are also expressed in the apical membranes and function in excreting organic cations into urine (Otsuka et al., 2005; Hiasa et al., 2006). Interaction of ipratropium with multidrug and toxic compound extrusion transporters should be clarified to elucidate the renal secretory pathway in detail.
Because the major OCT subtype in human kidney is OCT2, we further investigated the contribution of Oct2 to ipratropium uptake by rat kidney slices. As shown in Fig. 5, the Eadie-Hofstee plot indicates that the transport consists of two components, one with low affinity and one with high affinity. The Km of the high-affinity component is 0.114 ± 0.056 μM, which is almost equal to the Km obtained for ipratropium uptake by HEK293/rOct2 cells (Fig. 4). This result indicates that rOct2 transport activity was reflected in the uptake of ipratropium into proximal tubular cells at low concentrations (up to 100 nM). Furthermore, to separately evaluate the transport activities of rOct1 and rOct2, we used uncharged corticosterone, because the affinity of corticosterone for OCTs/Octs is subtype and species dependent. The IC50 values for corticosterone inhibition of TEA uptake by rOct1 and rOct2 have been previously reported as ∼150 and ∼4 μM, respectively (Arndt et al., 2001). In the present study, an approximately 50-fold difference between the IC50 values for corticosterone inhibition of ipratropium uptake by rOct1 (55.8 μM) and rOct2 (1.11 μM) was observed (Fig. 6, A and B). Hence, we considered that corticosterone at 1 μM selectively inhibits rOct2-mediated uptake of ipratropium by kidney slices. Under this condition, ipratropium uptake by the kidney slices was reduced by 18.4% (Table 2). Assuming that 50% of Oct2 activity was inhibited under these conditions, the contribution of Oct2 to ipratropium uptake in rat kidney could be estimated to be 40% at most. Even at 500 μM corticosterone, the ipratropium uptake by kidney slices remained at approximately 30% of the control, suggesting that a corticosterone-insensitive transporter might also be involved. These results strongly suggest that rOct2 contributes, at least in part, to renal excretion of ipratropium. In humans, OCT2 is more abundantly expressed in kidney than OCT1 and OCT3 (Kekuda et al., 1998; Koepsell et al., 2007). Because we were unable to detect significant transport of ipratropium or tiotropium by hOCT3 (Fig. 7C), hOCT2 is likely to play a major role in renal secretion of both anticholinergics.
Saturation kinetics of ipratropium uptake by HEK293/rOct2 cells gave a Km value of 0.143 ± 0.03 μM (Fig. 4D). This result indicates that ipratropium has a very high affinity for rOct2, in comparison with the Km values of other cationic drugs for rOct2 (e.g., 17 μM for MPP+ and 650 μM for dopamine) (Koepsell, 1998). A previous clinical study (The American Hospital Formulary Service Drug Information; Ipratropium Bromide Inhl) suggested that the mean peak plasma concentration of ipratropium in patients with COPD reached 84 pg/ml (equivalent to approximately 0.2 nM) after administration of a higher-than-recommended dosage (4 inhalations 4 times daily, 272 μg). At this concentration, hOCT2-mediated transport of ipratropium is not likely to be saturated, assuming that ipratropium possesses a similar affinity for the rat and human transporters. According to Table 1, ipratropium uptake by rat kidney slices was inhibited by cimetidine (1 mM), suggesting that ipratropium shares the same binding site(s) as cimetidine. Considering that the mean steady-state plasma concentration on a standard 1000-mg daily dose of cimetidine was reported to be 1.0 μg/ml (equivalent to approximately 4 μM) (Somogyi and Gugler, 1983), it is conceivable that renal clearance of ipratropium could be decreased by competitive inhibition of hOCT2-mediated transport by cimetidine, resulting in an increase of the plasma concentration.
Because it was documented that these anticholinergics do not interact with cimetidine or ranitidine (Keam and Keating, 2004), it had been thought that no significant alteration in the pharmacokinetics of these anticholinergics occurs when they are coadministered with cationic drugs. However, a clinical study showed that a single intravenous administration of tiotropium with cimetidine (400 mg three times a day) resulted in a 20% increase in the area under the concentration-time curve from 0 to 4 h and a 28% decrease in the total clearance (Montalto, 2004). This result could have been due to interaction of cimetidine with anticholinergics on OCT transporters in vivo. Therefore, the possibility of such an interaction should be taken into consideration in any concomitant use of these anticholinergics with cationic compounds, and the pharmacokinetics should be carefully monitored to avoid adverse effects.
After intravenous administration of [14C]tiotropium to rats, the highest concentrations of radioactivity were detected in kidney and liver within a short period (Leusch et al., 2001). In rat liver, rOct1 is located to the sinusoidal membrane of hepatocytes (Meyer-Wentrup et al., 1998), but rOct2 is not expressed in liver (Okuda et al., 1996). A similar distribution for ipratropium has been described previously (Forester et al., 1976; Rominger, 1979). These observations suggest that hepatic uptake of both ipratropium and tiotropium is mediated by OCT1/Oct1. Therefore, coadministration of these anticholinergics with cationic drugs may result in a decrease of their hepatic clearance, as well as renal clearance.
In conclusion, the results of this study indicate that OCTs/Octs, in particular OCT2/Oct2, play a role in renal secretion of ipratropium and tiotropium in rats and humans. Competition with coadministered cationic drug for OCTs/Octs may lead a decrease of renal excretion rate and hepatic clearance, resulting in an increase of the systemic concentration of ipratropium and tiotropium.
Authorship Contributions
Participated in research design: Nakanishi, Haruta, Shirasaka, and Tamai.
Conducted experiments: Nakanishi and Haruta.
Contributed new reagents or analytic tools: Nakanishi, Haruta, and Shirasaka.
Performed data analysis: Nakanishi, Haruta, and Shirasaka.
Wrote or contributed to the writing of the manuscript: Nakanishi and Tamai.
Other: Tamai acquired funding for the research.
Acknowledgments
We thank Dr. John Keogh at the GlaxoSmithKline R&D (Ware, UK) for providing the radiolabeled ipratropium and tiotropium.
Footnotes
- Received July 9, 2010.
- Accepted October 20, 2010.
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.110.035402.
↵
 The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.
The online version of this article (available at http://dmd.aspetjournals.org) contains supplemental material.ABBREVIATIONS:
- COPD
- chronic obstructive pulmonary disease
- ipratropium
- ipratropium bromide
- tiotropium
- tiotropium bromide
- M1–3
- muscarinic receptor 1 to 3
- hOCTN2
- human organic cation/carnitine transporter 2
- OCTN
- human organic cation/carnitine transporter
- MPP+
- l-methyl-4-phenylpyridinium
- OCT
- organic cation transporter
- HEK
- human embryonic kidney
- rOCT
- rat OCT
- hOCT
- human OCT
- qRT-PCR
- quantitative reverse transcription-polymerase chain reaction
- HPRT
- hypoxanthine-guaninephosphoribosyl-transferase.

- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}