


Abstract
Rat organic anion transporting polypeptide 2 (rOatp2) is a member of the OATP family. It exhibits broad substrate specificity and accepts amphipathic organic anions, cardiac glycosides (digoxin and ouabain; a neutral compound), and organic cations (rocuronium andN-(4,4-azo-n-pentyl)-21-deoxyajamalinium). In the present study, kinetic analyses were carried out to investigate whether taurocholate (TCA), digoxin, and 17β-estradiol-d-17β−glucuronide (E217βG) share the same recognition site on rOatp2 for their transport. The transport of TCA and digoxin was mutually inhibited, and theKi values of digoxin and TCA for the transport of TCA and digoxin were 0.58 and 160 μM, respectively. TheKm and Vmaxvalues of TCA and digoxin were 190 μM and 140 pmol/min/mg of protein and 1.1 μM and 6.6 pmol/min/mg of protein, respectively. TheKm and Ki values were consistent. In addition, digoxin (1 μM) and TCA (100 μM) increased the Km values of TCA and digoxin, respectively, but they did not affect theVmax values, suggesting that their inhibition is competitive. The transport of digoxin via rOatp2 was inhibited slightly by E217βG, whereas the uptake of TCA was stimulated by E217βG in a concentration-dependent manner. These results suggest that rOatp2 has at least two substrate recognition sites, one for TCA and digoxin and the other for E217βG.
Rat organic anion transporting polypeptide 2 (rOatp2; Slc21a5) is a member of the OATP family. The OATP family consists of two groups; one includes rat Oatp1 (Slc21a1), rOatp2, rOatp3 (Slc21a7), rOat-K1 (Slc21a4), rOat-K2, and human OATP-A (SLC21A6), which exhibit 70 to 80% amino acid identity with each other, whereas the other includes rOatp4/rlst1 (Slc21a10), hLST-1/OATP-C/OATP2 (SLC21A6), hOATP-D (SLC21A11), hOATP-E (SLC21A12), hOATP8 (SLC21A8), and prostaglandin transporter (SLC21A2), which exhibit about 40% amino acid identity with the members of the first group (Jacquemin et al., 1994;Kanai et al., 1995; Kullak-Ublick et al., 1995; Saito et al., 1996; Noe et al., 1997; Abe et al., 1998, 1999; Masuda et al., 1999;Cattori et al., 2000; Konig et al., 2000; Tamai et al., 2000). Members of the OATP family accept amphipathic organic anions, except for rOat-K1, and prostaglandin transporter in which substrates are limited to folate and its derivatives and prostaglandins, respectively (Kanai et al., 1995; Saito et al., 1996). The members of the OATP family play a significant role in drug disposition.
rOatp2 is expressed in the liver and brain (Noe et al., 1997; Abe et al., 1998). It is localized to the sinusoidal membrane of the hepatocyte, both the abluminal and the luminal membrane of brain capillary endothelial cells, and the basolateral membrane of the choroid plexus (Gao et al., 1999; Kakyo et al., 1999; Reichel et al., 1999). rOatp2 is involved in the transport into and out of the brain and liver. The efflux transport of E217βG across the blood-brain barrier is partially accounted for by rOatp2 (∼40%) (Sugiyama et al., 2001).
The substrate specificity of rOatp2 is broad and includes bile acids, steroid conjugates, peptides, such as BQ-123 (an endothelin receptor antagonist) and δ-opioid receptor agonists ([d-penicilliamine2,5] enkephalin, Leu-enkephalin, and deltorpin II), cardiac glycosides (ouabain and digoxin), and organic cations (rocuronium andN-(4,4-azo-n-pentyl)-21-deoxyajamalinium) (Kakyo et al., 1999; Reichel et al., 1999). Although no significant uptake of glutathione conjugates, such as leukotriene C4and dinitrophenyl glutathione, was observed by rOatp2-expressed oocytes, intracellular glutathione derivatives and conjugates, such as ophthalmic acid, dinitrophenyl glutathione, and bromosulfophthalein glutathione, stimulated the uptake of TCA and digoxin (Li et al., 2000). It has been suggested that rOatp2 accepts glutathione conjugates only from inside the cells (Li et al., 2000). Whether a single substrate recognition site can account for the broad substrate specificity of rOatp2 in the transport remains to be clarified. In the present study, kinetic analyses were carried out to investigate whether E217βG (organic anion), TCA (organic anion), and digoxin (neutral compound) share the same recognition site on rOatp2.
Materials and Methods
Chemicals.
[3H]E217βG (44 Ci/mmol), [3H]TCA (3.0 Ci/mmol), and [3H]digoxin (19 Ci/mmol) were purchased from PerkinElmer Life Sciences (Boston, MA). [3H]E217βG and [3H]digoxin were stored at −20°C, and [3H]TCA was stored as 4°C until used. Unlabeled E217βG and TCA were purchased from Sigma Chemical Co. (St. Louis, MO). Unlabeled digoxin was purchased from Aldrich Chemical Co. (Milwaukee, WI). All other chemicals were commercially available, of reagent grade, and used without any purification.
Cell Culture.
rOatp2-expressed LLC-PK1 cells have been established previously (Sugiyama et al., 2001). Transfectants were grown in M199 (Sigma Chemical Co.) supplemented with 10% fetal bovine serum, penicillin (100 U/ml), streptomycin (100 μg/ml), and G418 sulfate (400 μg/ml) at 37°C with 5% CO2 and 95% humidity on the bottom of a dish. Cells were seeded on 12-well multiwell plates at a density of 2.4 × 105 cells/well and cultured for 3 days. Sodium-butyrate (5 mM) was added to the culture medium 24 h before starting the transport experiments to induce the expression of rOatp2 (Sugiyama et al., 2001).
Transport Study.
Uptake was initiated by adding radiolabeled ligands to the medium in the presence and absence of inhibitors after cells had been washed three times and preincubated with Krebs-Henseleit buffer at 37°C for 15 min. The Krebs-Henseleit buffer consisted of 142 mM NaCl, 23.8 mM NaHCO3, 4.83 mM KCl, 0.96 mM KH2PO4, 1.20 mM MgSO4, 12.5 mM HEPES, 5 mM glucose, and 1.53 mM CaCl2 adjusted to pH 7.4. The uptake was terminated at designed times by adding ice-cold Krebs-Henseleit buffer. Then, cells were washed twice with 1 ml of ice-cold Krebs-Henseleit buffer dissolved in 500 μl of 0.2 N NaOH and kept overnight. Aliquots (350 μl) were transferred to scintillation vials after adding 50 μl of 2 N HCl. The radioactivity associated with the cells and medium was determined in a liquid scintillation counter (LS 6000SE; Beckman Coulter, Inc., Fullerton, CA) after adding 2 ml of scintillation fluid (NACALAI TESQUE, Kyoto, Japan) to the scintillation vials. Ligand uptake is given as the cell-to-medium concentration ratio determined as the amount of ligand associated with the cells divided by the medium concentration. All inhibitors except digoxin were dissolved in Krebs-Henseleit buffer. Because of the low water solubility of digoxin, it was dissolved in dimethyl sulfoxide and subsequently diluted 1:200 in Krebs-Henseleit buffer. Preliminary experiments showed that the presence of 0.5% dimethyl sulfoxide had no effect on the uptake of substrates.
Kinetic Analyses.
Kinetic parameters were obtained using the following equation (Michaelis-Menten equation): v =VmaxS/(Km + S), wherev is the uptake rate of the substrate (picomoles per minute per milligram of protein), S is the substrate concentration in the medium (micromolar concentration),Km is the Michaelis-Menten constant (micromolar concentration), and Vmaxis the maximum uptake rate (picomoles per minute per milligram of protein). To obtain the kinetic parameters, the equation was fitted to the rOatp2-mediated uptake velocity, which was calculated by subtracting the uptake value of the substrate into vector-transfected LLC-PK1 cells from that into rOatp2-expressing cells. The experimental data were fitted to the equation by least-squares regression analysis with weighting as the reciprocal of the observed values, and the Damping Gauss-Newton method algorithm was used for fitting. Inhibition constants (Ki) for rOatp2-mediated transport were calculated assuming competitive inhibition.
Results
Time-Profiles for the Uptake of TCA and Digoxin by rOatp2-Expressed LLC-PK1 Cells.
Time-profiles of the uptake of TCA and digoxin by rOatp2-transfected LLC-PK1 cells are shown in Fig. 1. Transfection of rOatp2 to LLC-PK1 cells results in an increase in the uptake of TCA and digoxin (Fig. 1). rOatp2-mediated TCA and digoxin uptake increased linearly until 5 min. The uptake of E217βG by vector-transfected and rOatp2-expressed LLC-PK1 cells at 5 min was 4.7 ± 0.4 and 8.9 ± 0.3 μl/mg of protein, respectively, and this rOatp2-mediated uptake of E217βG increased linearly up to 5 min (Sugiyama et al., 2001).

Time-profiles of the uptake of [3H]TCA and [3H]digoxin by rOatp2-expressed LLC-PK1 cells.
The uptake of [3H]TCA (3 μM) (A) and [3H]digoxin (10 nM) (B) by rOatp2-expressed LLC-PK1 cells was examined at 37°C. Closed (▪) and open (■) squares represent the uptake by rOatp2-expressed cells and vector-transfected cells, respectively. The uptake was initiated by adding radiolabeled ligand ([3H]TCA or [3H]digoxin) and terminated at designed times by adding ice-cold buffer. Each point represents the mean ± S.E. (n = 3).
Effect of E217βG on the Uptake of TCA and Digoxin by rOatp2-Expressed LLC-PK1 Cells.
The effect of E217βG on the uptake of TCA and digoxin was examined in rOatp2-expressed LLC-PK1 cells (Fig.2). The uptake of digoxin by rOatp2 was only slightly inhibited by E217βG even at 100 μM (Fig. 2B). On the other hand, the uptake of TCA in rOatp2 was stimulated by E217βG in a concentration-dependent manner (Fig. 2A). The uptake of [3H]TCA and [3H]digoxin by vector-transfected LLC-PK1 cells at 5 min was unaffected by unlabeled E217βG, and the average value was 3.4 ± 0.2 and 9.2 ± 0.5 μl/mg of protein, respectively.
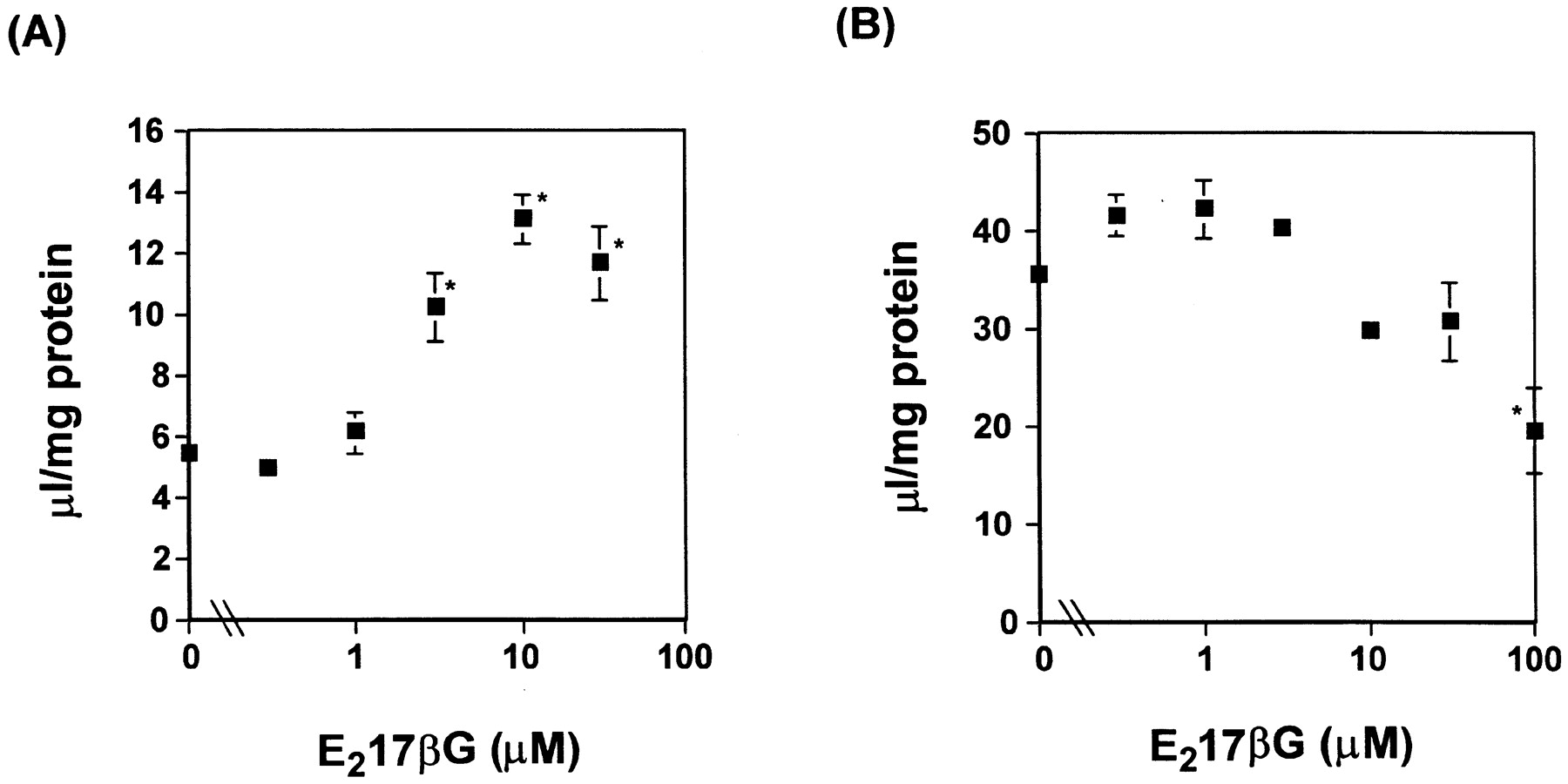
Effects of unlabeled E217βG on the uptake of [3H]TCA and [3H]digoxin into rOatp2-expressed LLC-PK1 cells.
The effects of unlabeled E217βG on the uptake of [3H]TCA (3 μM) (A) and [3H]digoxin (10 nM) (B) by rOatp2-expressed LLC-PK1 cells were examined at 37°C. The uptake was initiated by adding radiolabeled ligand ([3H]TCA or [3H]digoxin) and unlabeled E217βG and terminated at 5 min by adding ice-cold buffer. Closed (▪) squares represent the specific uptake determined at 5 min by subtracting the uptake by vector-transfected cells from that by rOatp2-expressed cells. Each point represents the mean ± S.E. (n = 3). ★, represents a statistically significant difference compared with the uptake in the absence of E217βG (P < 0.05).
Inhibitory Effect of TCA and Digoxin on rOatp2-Mediated Uptake.
The concentration-dependence of the rOatp2-mediated uptake of TCA and digoxin is shown in Fig. 3. TheKm andVmax values for the uptake of TCA and digoxin by rOatp2 were 187 ± 46 μM and 140 ± 25 pmol/min/mg of protein and 1.07 ± 0.18 μM and 6.57 ± 0.87 pmol/min/mg of protein, respectively (Table1). Digoxin inhibited the uptake of TCA via rOatp2 in a concentration-dependent manner, with aKi value of 0.58 ± 0.27 μM (Table 1). Similarly, TCA inhibited the uptake of digoxin via rOatp2 with a Ki value of 156 ± 23 μM (Table 1). These Ki values of TCA and digoxin are consistent with their Kmvalues. According to the previous study, the uptake of E217βG in rOatp2 expressed LLC-PK1 cells was inhibited by both TCA and digoxin withKi values of 39 and 0.037 μM, respectively (Table 1; Sugiyama et al., 2001). TheKi values of TCA and digoxin for the uptake of E217βG were approximately 4- and 10-fold smaller than those determined in this study.

Eadie-Hofstee plots of the uptake of [3H]TCA and [3H]digoxin in the presence and absence of unlabeled TCA and digoxin by rOatp2-expressed cells.
The rOatp2-mediated uptake of [3H]TCA (A) and [3H]digoxin (B) was measured at different substrate concentrations in the presence (■) and absence (▪) of unlabeled digoxin (1 μM) (A) and unlabeled TCA (100 μM) (B), respectively. Specific uptake was determined by subtracting the uptake by vector-transfected cells from that by rOatp2-expressed cells. Each point represents the mean ± S.E. (n = 3). The data were fitted to the Michaelis-Menten equation by nonlinear regression analysis, as described under Materials and Methods, and the solid line represents the fitted curve.
Km and Ki values of TCA, digoxin, and E217βG for Oatp2
The Km andVmax values for the transport of TCA by rOatp2 in the presence of digoxin (1 μM) were found to be 291 ± 72 μM and 148 ± 26 pmol/min/mg of protein, respectively, and those for the uptake of digoxin by rOatp2 in the presence TCA (100 μM) were 3.18 ± 1.30 μM and 8.83 ± 3.14 pmol/min/mg of protein, respectively. Since nonradiolabeled digoxin and TCA increased the Km values but did not affect theVmax values for the transport of TCA and digoxin, their inhibition is competitive.
Discussion
In this study, kinetic analyses were carried out to investigate rOatp2-mediated transport of TCA, digoxin, and E217βG. TCA, digoxin, and E217βG are known to be substrates of rOatp2 (Noe et al., 1997; Abe et al., 1998), and this was confirmed in our studies (Fig. 1; Sugiyama et al., 2001). Compared with the uptake of digoxin TCA, the transport activity of E217βG by rOatp2 is quite low, and E217βG seems to be a poor substrate for rOatp2.
The Km values of TCA, digoxin, and E217βG for rOatp2 were found to be 140, 1.07, and 17.0 μM, respectively (Table 1). In previous studies using aXenopus laevis oocyte expression system, theKm values of TCA, digoxin, and E217βG for rOatp2 were 35, 0.24, and 3.0 μM, respectively (Noe et al., 1997; Abe et al., 1998). Although the rank order of the affinity of these ligands for rOatp2 is the same as far as our own and previous results are concerned, the absoluteKm values of these ligands for rOatp2 determined in this study were about 5-fold greater than previously reported values for some unknown reason. This may be caused by a difference in the microenvironment, such as the lipid composition, between oocytes and mammalian cells.
The Km values of digoxin and TCA are comparable to their Ki values for the uptake of TCA and digoxin in rOatp2-expressed LLC-PK1 cells (Table 1). In addition, the result indicating that TCA and digoxin competitively inhibited each other's uptake suggests that TCA and digoxin share the same recognition site on rOatp2.
The uptake of E217βG by rOatp2 was saturated, with a Km value of 17 μM (Sugiyama et al., 2001). However, E217βG only slightly inhibited the transport of digoxin even at 100 μM, which is sufficient to saturate the transport E217βG by rOatp2 (Fig. 2b). In contrast, E217βG actually stimulated the uptake of TCA in a concentration-dependent manner. The previously reported Ki values of digoxin and TCA for the uptake of E217βG via rOatp2 are relatively smaller than theirKm values (Sugiyama et al., 2001). These results suggest that E217βG interacts with rOatp2 at a different site from that for TCA and digoxin. Although kinetic studies suggest that TCA and digoxin share the same site (Table1; Fig. 3), the inverse effects of E217βG on the uptake of TCA and digoxin suggest that TCA and digoxin also interact with rOatp2 at different sites. Further detailed studies, including binding assays, are required to confirm whether rOatp2 recognizes TCA, digoxin, and E217βG at different sites or not.
TCA, digoxin, and E217βG have a similar chemical structure, namely a steroid nucleus with different attached groups, taurine, digitoxose, and glucuronate, respectively. The results of this study suggest that they are not necessary recognized by rOatp2 in a similar manner. Three-dimensional structure-activity relationship analysis is helpful for investigating which structure and/or moiety plays a key role in the recognition by rOatp2 with different recognition sites and for revealing the molecular mechanism governing the broad substrate specificity of rOatp2.
Substrate-dependent kinetic parameters have been also reported for cytochrome P450 enzymes, such as CYP3A and CYP2C9 (Korzekawa et al., 1998; Wang et al., 2000), and multidrug resistant 1 (MDR1) P-glycoprotein (Ayesh et al., 1996; Orlowski et al., 1996; Garrigos et al., 1997; Pascaud et al., 1998; Buxbaum, 1999). These facts, together with this article, must be taken into consideration, especially when the possibility of drug-drug interactions is evaluated using high-throughput screening. The Ki or IC50 values for the transport of a particular drug need to be examined instead of that for the transport of typical substrates to avoid any false negative predictions. Further studies are required to confirm whether kinetic parameters are substrate-dependent in human organic anion transporters, such as hLST-1/OATP2/OATP-C and hOATP8, which are considered to play a significant role in the hepatic uptake of organic anions.
In conclusion, our present results demonstrate that the kinetic parameters for rOatp2 are substrate-dependent and suggest that there are at least two recognition sites for the uptake by rOatp2. One is for TCA and digoxin, and the other is for E217βG.
Footnotes
-
This work was supported by the Ministry of Education, Science, and Culture of Japan and CREST (Core Research for Evolutional Science and Technology) of Japan Science and Technology Corporation.
- Abbreviations used are::
- OATP
- organic anion transporting polypeptide
- E217βG
- 17β-estradiol-d-17β-glucuronide
- TCA
- taurocholate
- Received March 19, 2001.
- Accepted November 7, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}