


Abstract
A metabolite formed by incubation of human liver microsomes, etoposide, and UDP-glucuronic acid was identified as etoposide glucuronide by liquid chromatography-tandem mass spectrometry analysis. According to the derivatization with trimethylsilylimidazole (Tri-Sil-Z), it was confirmed that the glucuronic acid is linked to an alcoholic hydroxyl group of etoposide and not to a phenolic group. Among nine recombinant human UGT isoforms (UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A8, UGT1A9. UGT1A10, UGT2B7, and UGT2B15), only UGT1A1 exhibited the catalytic activity of etoposide glucuronidation. The enzyme kinetics in pooled human liver microsomes and recombinant UGT1A1 microsomes showed a typical Michaelis-Menten plot. The kinetic parameters of etoposide glucuronidation wereKm = 439.6 ± 70.7 μM andVmax = 255.6 ± 19.2 pmol/min/mg of protein in human liver microsomes andKm = 503.2 ± 110.2 μM andVmax = was 266.5 ± 28.6 pmol/min/mg of protein in recombinant UGT1A1. The etoposide glucuronidation in pooled human liver microsomes was inhibited by bilirubin (IC50 = 31.7 μM) and estradiol (IC50 = 34 μM) as typical substrates for UGT1A1. The inhibitory effects of 4-nitrophenol (IC50 = 121.0 μM) as a typical substrate for UGT1A6 and UGT1A9, imipramine (IC50 = 393.8 μM) as a typical substrate for UGT1A3 and UGT1A4, and morphine (IC50 = 109.3 μM) as a typical substrate for UGT2B7 were relatively weak. The interindividual difference in etoposide glucuronidation in 13 human liver microsomes was 78.5-fold (1.4–109.9 pmol/min/mg of protein). The etoposide glucuronidation in 10 to 13 human liver microsomes was significantly correlated with β-estradiol-3-glucuronidation (r= 0.841, p < 0.01), bilirubin glucuronidation (r = 0.935, p < 0.01), and the immunoquantified UGT1A1 protein content (r = 0.800,p < 0.01). These results demonstrate that etoposide glucuronidation in human liver microsomes is specifically catalyzed by UGT1A1.
Etoposide [4′-demethylepipodophyllotoxin-9-(4,6-O-ethylidene)-β-d-glucopyranoside] is one of the clinically important antitumor agents derived from 4′-demethylepipodophyllotoxin, which is an extract from the plantsPodophyllum peltatum and Podophyllum emodi (Clark and Slevin, 1987; Stähelin and von Wartburg, 1991). It is widely used in the treatment of testicular cancer, small cell lung cancer, and certain lymphomas (O'Dwyer et al., 1985). Etoposide causes tumor cell killing through DNA strand breakage resulting from the interaction of etoposide with the enzyme topoisomerase II and DNA (Ross et al., 1984).
In humans, the disposition of etoposide is described as a biphasic process with a distribution half-life of about 1.5 h and terminal elimination half-life ranging from 4 to 11 h (PDR, 2000). Clearance of etoposide occurs by direct renal excretion and metabolism. Roughly 35% of the administered drug is excreted into urine as a parent drug (Hande et al., 1984; Sinkule et al., 1984), but less than 3% is excreted into bile (Joel et al., 1996). Several metabolites were identified in human plasma and urine such as hydroxy acid derivatives,cis-(picro) lactone, 3′-demethyletoposide, and etoposide glucuronide (Clark and Slevin, 1987; Stewart, 1994). It has been reported that 58 and 19% of the administered drug is excreted as hydroxy acid into urine and bile, respectively (Clark and Slevin, 1987); less than 5 and 1% of the administered drug is found ascis-(picro) lactone in plasma and urine, respectively (Holthuis et al., 1986). It has been reported that 3′-demethylation of etoposide, a minor metabolite, is catalyzed by CYP3A4 (Relling et al., 1994). Etoposide glucuronide accounts for the disposition of 15 to 35% of administered etoposide (D'Incalci et al., 1986; Hande et al., 1988).
Glucuronidation is catalyzed by UDP-glucuronosyltransferase (UGT1) enzymes (Miners and Mackenzie, 1991). It is well known that there are many isoforms of mammalian UGT enzymes (Tukey and Strassburg, 2000). UGT1 and UGT2 have been shown to catalyze the glucuronidation of xenobiotics. The UGT1 and UGT2 genes seem to be structurally different in that UGT1 proteins result from alternate splicing of different first exons with five shared exons encoded by the UGT1 gene complex, whereas UGT2 proteins seem to be encoded by unique genes. In the human genome, at least 13 different first exons have been identified for the UGT1 gene (Gong et al., 2001). Currently, several common genetic polymorphisms in UGTs have been described, although their effects on the potency of glucuronidations have not been well studied. The purpose of the present study was to identify UGT isoform(s) involved in etoposide glucuronidation and to characterize the interindividual variability in etoposide glucuronidation using human liver microsomes. UGT isoforms relevant to this variability were identified by screening a panel of recombinant UGTs microsomes and by chemical inhibition of specific UGT substrates. Furthermore, correlation analyses of etoposide glucuronosyltransferase activity with glucuronosyltransferase activities of other substrates and immunoquantified UGT1A1 content were determined.
Materials and Methods
Materials.
Etoposide, bilirubin, 4-nitrophenol, and imipramine hydrochloride were purchased from Wako Pure Chemicals (Osaka, Japan). UDP-glucuronic acid, alamethicin, β-estradiol, p-nitrophenyl β-glucuronide, and α-naphthyl β-glucuronide were from Sigma-Aldrich (St. Louis, MO). Morphine hydrochloride was purchased from Takeda Chemical Industries (Osaka, Japan). Morphine-3-glucuronide was kindly provided by Dr. Kazuta Oguri (Kyusyu University, Fukuoka, Japan). Tri-Sil-Z, a mixture of trimethylsilylimidazole in dry pyridine (1.5 mEq/ml = 1 part trimethylsilylimidazole:4 parts pyridine), was purchased from Pierce Chemical (Rockford, IL). Pooled human liver microsomes (H161) and microsomes from 13 individual human livers (H003, H023, H042, H043, H056, H064, H066, H089, H093, H095, H112, HK23, and HK34) were purchased from BD Gentest (Woburn, MA). The glucuronosyltransferase activities of β-estradiol (UGT1A1), trifluoperazine (UGT1A4), and propofol (UGT1A9) in these human liver microsomes were provided by the manufacturer. The glucuronosyltransferase activities of bilirubin (UGT1A1) and immunoquantified UGT1A1 protein in these human liver microsomes except for H064, H095, and HK34 were also provided by the manufacturer. Recombinant human UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A8, UGT1A9, UGT1A10, UGT2B7, and UGT2B15 expressed in baculovirus-infected insect cells (Supersomes) were also from BD Gentest. All other chemicals and solvent were of the highest grade commercially available.
Preparation of Rat Liver Microsomes.
Male Wistar and Gunn rats (6 weeks old, n = 3) were purchased from Japan SLC (Hamamatsu, Japan). Liver microsomes from untreated rats were prepared as described previously (Guengerich, 1994). Liver microsomes were suspended in 10 mM Tris-HCl buffer (pH 7.4) containing 10 mM EDTA and 20% (v/v) glycerol. After determination of the protein concentration (Lowry et al., 1951), the suspended microsomal samples were frozen and stored at −80°C until use.
Etoposide Glucuronidation Assay.
A typical incubation mixture (200-μl total volume) contained 25 mM Tris-HCl buffer (pH 7.4), 5 mM MgCl2, 3 mM UDP-glucuronic acid, 50 μg/ml alamethicin, 1.0 mg/ml liver microsomes (1 mg/ml for recombinant UGT), and 400 μM etoposide. The reactions were initiated by the addition of UDP-glucuronic acid and were then incubated at 37°C for 60 min. The reactions were terminated by boiling at 100°C for 10 min. After removal of the protein by centrifugation at 10,000 rpm for 5 min, a 100-μl portion of the sample was subjected to high-performance liquid chromatography. Chromatography was performed using an LC-6A pump, an SPD-6 UV detector, an SIL-6B autosampler, a C-R4A integrator, and a CTO-6A column oven (all from Shimadzu, Kyoto, Japan) with a Mightysil RP-18 column (150 × 4.6-mm, 5 μm; Kanto Chemical, Tokyo, Japan). The flow rate was 1.0 ml/min and the column temperature was 35°C. The eluate was monitored at 254 nm with a noise-base clean Uni-3 (Union, Gunma, Japan). The Uni-3 can reduce the noise by integration of the output and increase the signal 3-fold by differentiation of the output and 5-fold by further amplification with an internal amplifier, resulting in a maximum 15-fold amplification of the signal. The mobile phase was 20% CH3CN/1% CH3COOH (v/v). The retention times of etoposide glucuronide and etoposide were 18.0 and 32.0 min, respectively. None of these chromatograms showed any interfering peaks with etoposide glucuronide. We present the activities for the formation of etoposide glucuronide on the basis of the chromatographic peak height using etoposide as a standard.
Identification of Etoposide Glucuronide by LC-MS/MS Analysis.
LC-MS/MS analysis was performed using an LCQDeca (Thermoquest, San Jose, CA) under electrospray ionization (ESI) conditions. The operation conditions used were capillary temperature, 350°C; capillary volt, −10 V; tube lens volt, 25 V; ion spray volt, 4.5 V; sheath gas, N2; pressure, 80 psi; auxiliary gas, N2, 20 l/min; and collision energy, 50%. Liquid chromatography was performed using an HP1100 (Agilent Technologies, Palo Alto, CA) with a Symmetry C18 column (150 × 4.6 mm, 3.5 μm; Waters, Milford, MA). The flow rate was 1.0 ml/min and the column temperature was 40°C. The mobile phase was 20% CH3CN/0.1% HCOOH (v/v). The retention times of etoposide glucuronide and etoposide were 10.9 and 30.3 min, respectively.
For the treatment with Tri-Sil-Z, etoposide glucuronide was fractionated by high-performance liquid chromatography and evaporated under N2 gas at room temperature. The sample was dissolved with 50 μl of CH3CN and 50 μl of Tri-Sil-Z was added. The mixtures were incubated for 30 min at 50°C. The resultant derivatives were measured by the method of flow injection and atmospheric pressure chemical ionization (APCI) with a positive mode.
Kinetic Analyses of Etoposide Glucuronidation.
The kinetics studies were performed using pooled human liver microsomes, recombinant human UGT1A1, and rat liver microsomes from three Wistar and Gunn rats. In determining the kinetic parameters, the etoposide concentration ranged from 50 μM to 1 mM. Kinetic parameters were estimated from the fitted curves using a computer program of KaleidaGraph (Synergy Software, Reading, PA) designed for nonlinear regression analysis. The following equations were applied for assuming a Michaelis-Menten equation: V =Vmax · S/(Km + S), whereKm is a Michaelis-Menten constant andVmax is maximum velocity.
Inhibition Analysis of Etoposide Glucuronidation in Human Liver Microsomes.
Five compounds were tested for their inhibitory effects on the etoposide glucuronosyltransferase activity in pooled human liver microsomes. Bilirubin is a typical substrate for UGT1A1 (Bosma et al., 1994; King et al., 1996). β-Estradiol is a typical substrate for UGT1A1 (and UGT1A9 as a minor enzyme) (Bosma et al., 1994; Hanioka et al., 2001a,b). 4-Nitrophenol is a substrate for UGT1A6 and UGT1A9 (Hanioka et al., 2001a,b). Imipramine is a substrate for UGT1A4 (and UGT1A3 as a minor enzyme) (Green et al., 1995). Morphine is a substrate for UGT2B7 (Coffman et al., 1997). Bilirubin and 4-nitrophenol were dissolved in dimethyl sulfoxide and ethanol, respectively. β-Estradiol was dissolved in methanol. Imipramine hydrochloride and morphine hydrochloride were dissolved in water. The final concentration of the organic solvents in the reaction mixture was <1% (v/v).
Other Glucuronidation Assay.
4-Nitrophenol, 1-naphthol, and morphine glucuronosyltransferase activities in human liver microsomes were determined as described previously (Nakajima et al., 2002; Watanabe et al., 2002).
Correlation Analyses.
Correlation analyses between etoposide glucuronidation and the other glucuronidation activities and immunoquantified UGT1A1 protein content were determined by Pearson's product moment method. A pvalue of less than 0.01 was considered statistically significant.
Results
Etoposide Glucuronidation in Human Liver Microsomes.
Etoposide glucuronide formation in the pooled human liver microsomes was investigated. The formation of the metabolite increased in a microsomal protein concentration- and time-dependent manner and was dependent on the concentration of UDP-glucuronic acid. The formation was linear at least at 2 mg/ml microsomal protein and 120-min incubation. Unless specified, the standard incubation mixture containing 1.0 mg/ml microsomal protein and 3 mM UDP-glucuronic acid was incubated at 37°C for 60 min. The etoposide glucuronosyltransferase activity in the pooled human liver microsomes was 115.5 pmol/min/mg of protein. In our preliminary study, the etoposide glucuronidation in human liver microsomes was assayed in the presence of saccharic acid 1,4-lactone, an inhibitor of β-glucuronidase, to investigate the effect of β-glucuronidase. However, the addition of 10 mM saccharic acid 1,4-lactone did not affect the etoposide glucuronosyltransferase activity.
LC-MS/MS Analyses of Etoposide Glucuronide.
The ESI mass spectrum of etoposide showed [M − H]− ion at m/z 587 and [M + CH3COOH-H]− ion atm/z 647 (Fig. 1A). The ESI mass spectrum of the peak typically formed by incubation of etoposide with human liver microsomes and UDP-glucuronic acid is shown in Fig.1B. [M − H]− ion at m/z 763 corresponding to etoposide glucuronide was observed. The product ion spectrum of the peak showed [M − H]−ions at m/z 587 corresponding to etoposide (Fig. 1D). Other product ions at m/z 381 (due to the loss of glycopyranosyl moiety of etoposide), 337, 322, and 307 (Fig. 1D) were the same as those of the product ions of etoposide (Fig. 1C). From these observations, it was confirmed that the peak formed by the incubation of etoposide with human liver microsomes and UDP-glucuronic acid was etoposide glucuronide. After the treatment of etoposide with Tri-Sil-Z, the APCI mass spectrum of the peak showed [M + H]+ ion at m/z 805 corresponding to tri-trimethysilylated etoposide (Fig. 1E). The product ion atm/z 455 corresponded to the trimethylsilylated podophyllotoxin moiety (Fig. 1E). After the treatment of the metabolite with Tri-Sil-Z, the APCI mass spectrum of the peak did not show the parent ion. However, the product ion at m/z 455 (trimethylsilylated podophyllotoxin moiety), which was the same as the product ion obtained from the treatment of etoposide was obtained (Fig.1, E and F). These results suggested that glucuronic acid is linked to the alcoholic hydroxyl group and not to the phenolic group.

ESI mass spectrum (A and B), product ion spectrum (C and D), and APCI mass spectrum after treatment with Tri-Sil-Z (E and F) of etoposide (A, C, and E) and the peak formed by the incubation of etoposide with human liver microsomes and UDP-glucuronic acid (B, D, and F).
[M − H]− ion at m/z 587 and [M + CH3COOH-H]− ion at m/z 647 corresponds to etoposide (A). [M − H]− ion atm/z 763 corresponds to etoposide glucuronide (B). Product ion spectrum showed peaks at m/z 381 corresponding to the podophyllotoxin moiety (C and D). The product ion at m/z 455 corresponded to the trimethylsilylated podophyllotoxin moiety (E and F).
Etoposide Glucuronidation in Recombinant UGT Isoforms.
Ten recombinant UGT isoforms expressed in baculovirus-infected insect cells were used to determine their etoposide glucuronosyltransferase activities. As shown in Fig. 2, only UGT1A1 exhibited etoposide glucuronosyltransferase activities (103.2 pmol/min/mg of protein).

Etoposide glucuronosyltransferase activity in recombinant human UGTs.
Etoposide glucuronosyltransferase activities in UGT enzymes were determined. Human UGTs (1 mg/ml) were incubated with 400 μM etoposide and 3 mM UDP-glucuronic acid at 37°C for 60 min. Each data point represents the mean of duplicate determinations.
Kinetics of Etoposide Glucuronidation in Human Liver Microsomes and Recombinant Human UGT1A1.
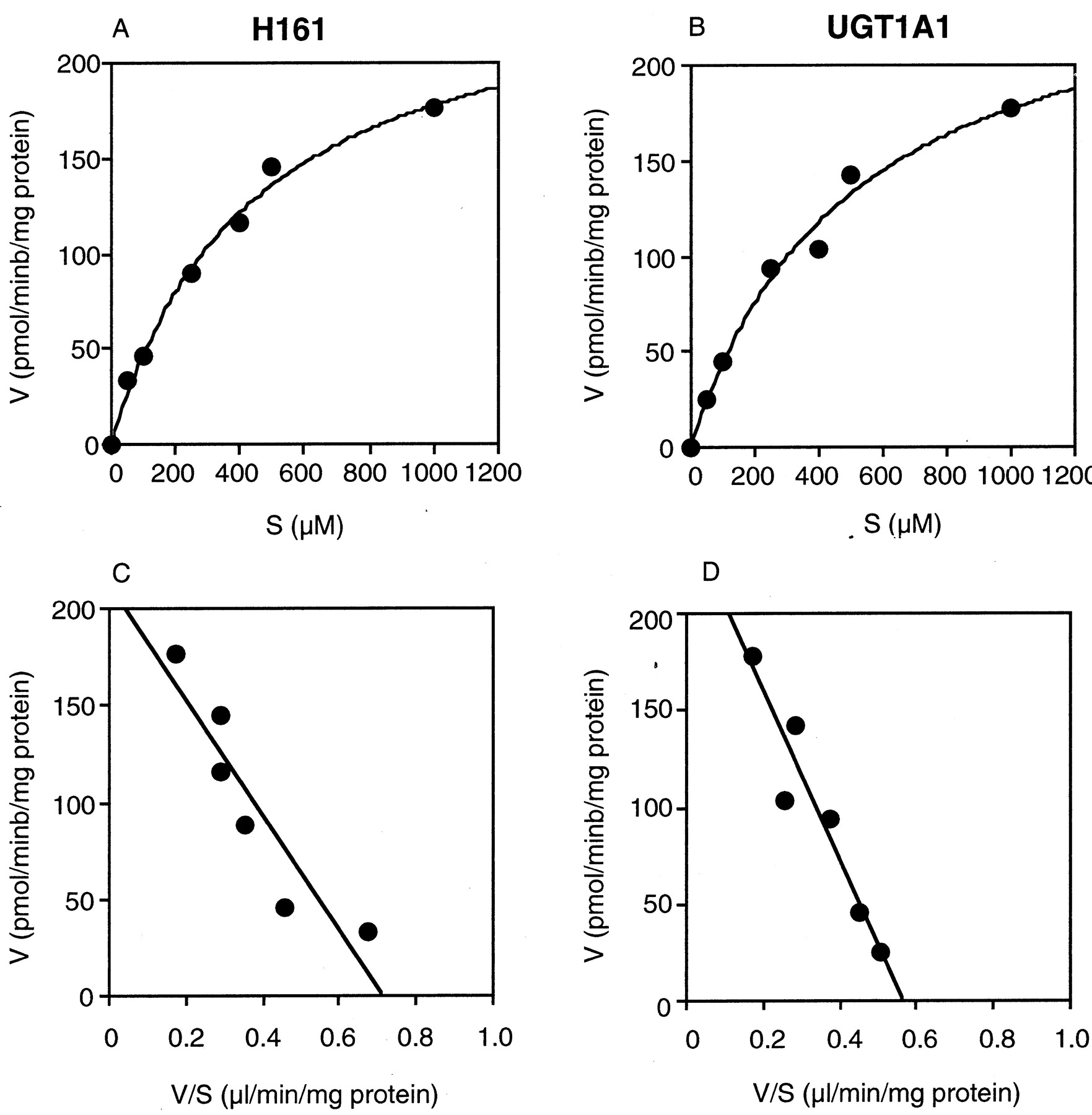
Kinetic analyses of etoposide glucuronidation in pooled human liver microsomes (H161) and recombinant human UGT1A1 were performed. The kinetics fitted the Michaelis-Menten kinetics (Fig.3, A and B) and the Eadie-Hofstee plots were monophasic (Fig. 3, C and D). When the apparent enzyme kinetic parameters were estimated, the Kmvalue was 439.6 ± 70.7 μM and theVmax value was 255.6 ± 19.2 pmol/min/mg of protein for human liver microsomes; theKm value was 503.2 ± 110.2 μM and the Vmax value was 266.5 ± 28.6 pmol/min/mg of protein for recombinant human UGT1A1.

Kinetics of etoposide glucuronidation in human liver microsomes (H161) and human UGT1A1.
The etoposide concentration ranged from 50 μM to 1 mM. The etoposide glucuronosyltransferase activities were determined as described underMaterials and Methods. Each data point represents the mean of duplicate determinations.
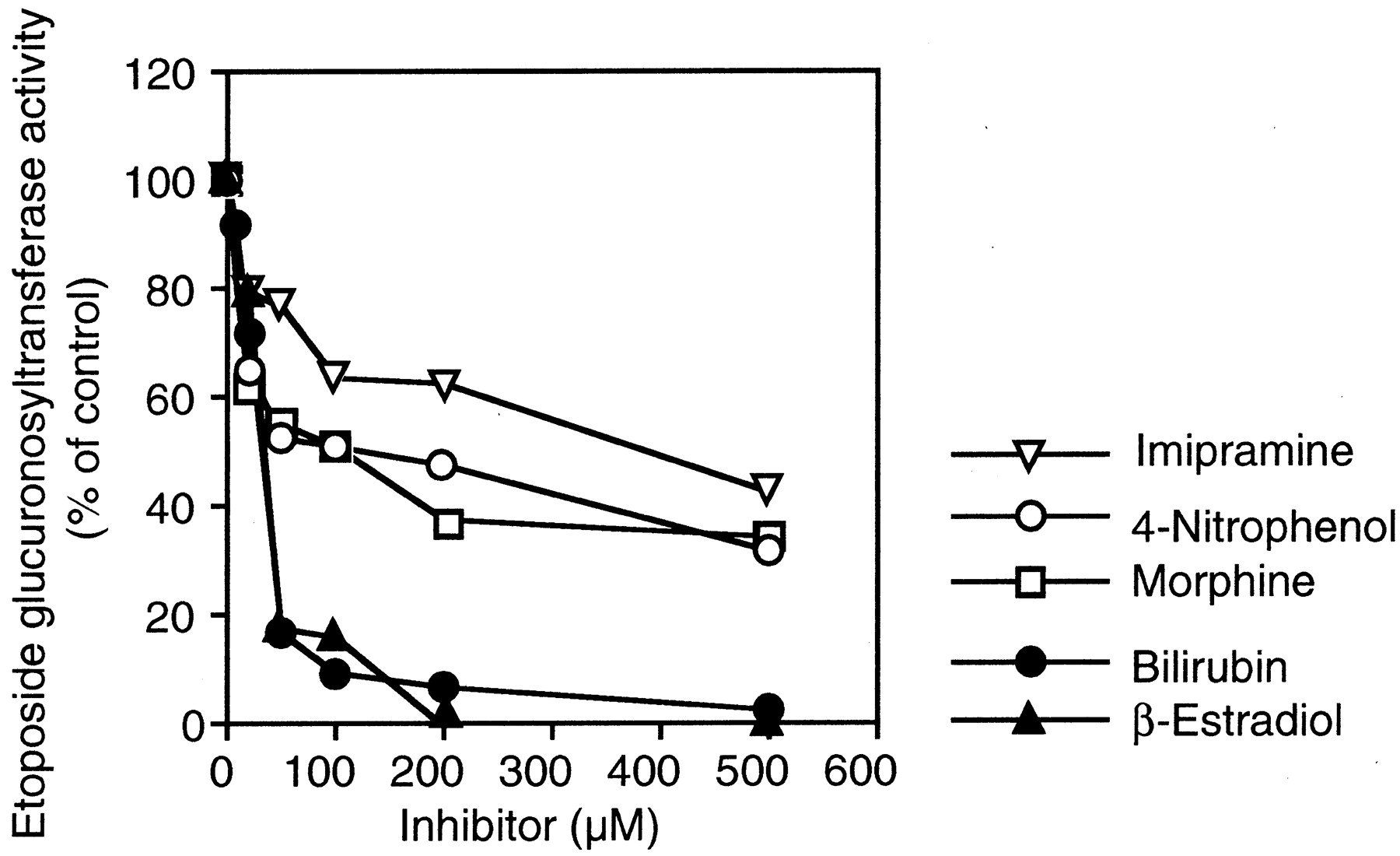
Inhibition Analyses of Etoposide Glucuronidation in Human Liver Microsomes.
The effects of bilirubin (UGT1A1), β-estradiol (UGT1A1 and UGT1A9), 4-nitrophenol (UGT1A6 and UGT1A9), imipramine (UGT1A3 and UGT1A4), and morphine (UGT2B7) on the etoposide glucuronosyltransferase activity in the pooled human liver microsomes were determined. As shown in Fig.4, the etoposide glucuronosyltransferase activity in the pooled human liver microsomes was inhibited by bilirubin (IC50 = 31.7 μM) and β-estradiol (IC50 = 34 μM). The inhibitory effects of 4-nitrophenol (IC50 = 121.0 μM), imipramine (IC50 = 393.8 μM), and morphine (IC50 = 109.3 μM) were relatively weak.
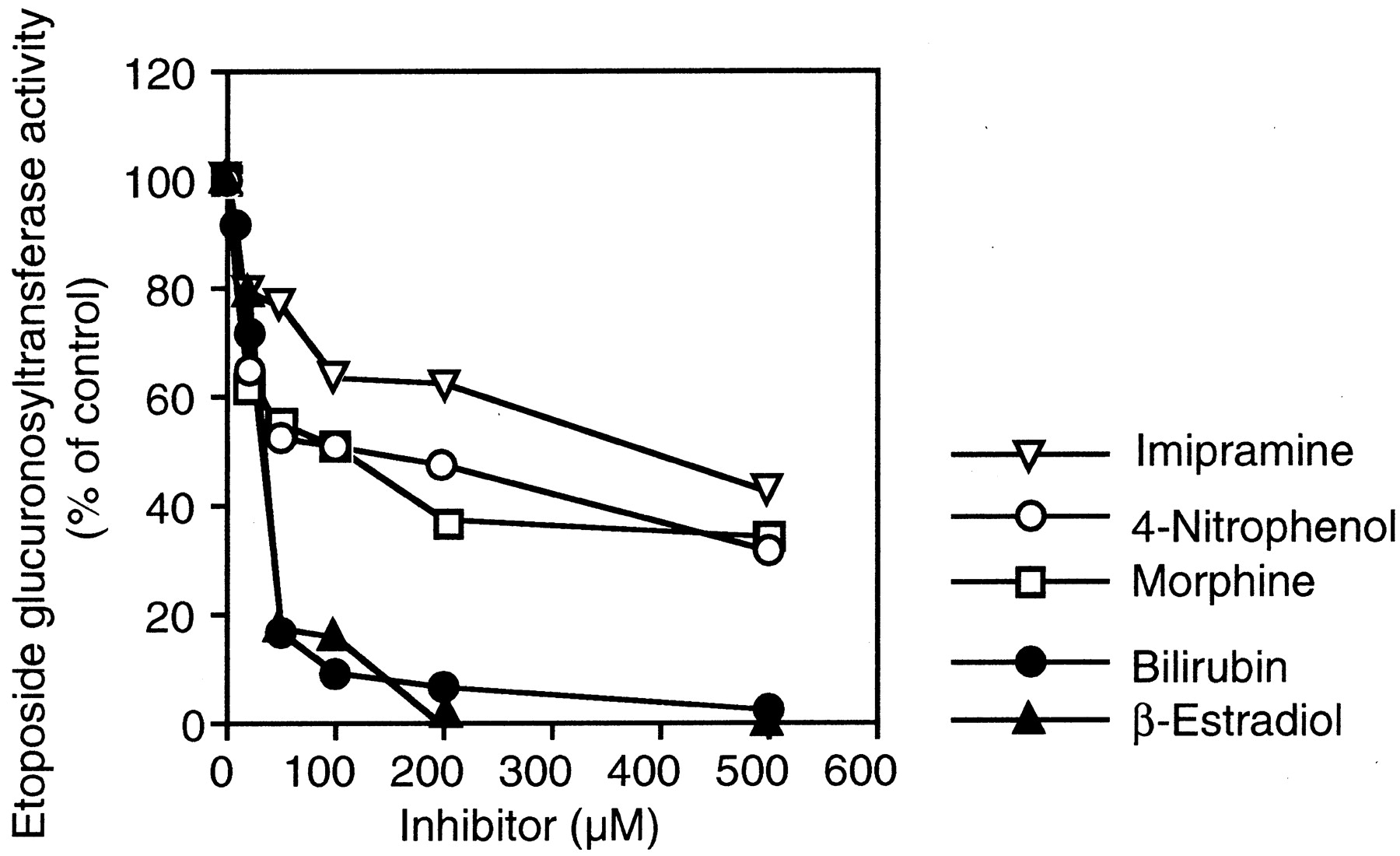
Inhibitory effects of typical substrates for UGT isoforms on etoposide glucuronosyltransferase activity in human liver microsomes.
The etoposide glucuronosyltransferase activity at 400 μM etoposide in pooled human liver microsomes (H161) was determined as described underMaterials and Methods. Bilirubin (UGT1A1), β-estradiol (UGT1A1 and UGT1A9), 4-nitrophenol (UGT1A6 and UGT1A9), imipramine (UGT1A3 and UGT1A4), and morphine (UGT2B7) are typical substrates for each UGT isoform. Etoposide glucuronosyltransferase activity in the pooled human liver microsomes in the absence of compound was 118.6 pmol/min/mg of protein. Each data point represents the mean of duplicate determinations.
Interindividual Variability of Etoposide Glucuronosyltransferase Activity in Microsomes from 13 Human Livers and Correlation with the Other Glucuronosyltransferase Activities.
Etoposide glucuronosyltransferase activities in microsomes from 13 human livers were determined at 400 μM etoposide (Fig.5A). The interindividual difference in etoposide glucuronosyltransferase activity was as large as 78.5-fold (1.4–109.9 pmol/min/mg of protein). Correlation analyses were performed between the etoposide glucuronosyltransferase activity and bilirubin (UGT1A1), β-estradiol (UGT1A1), trifluoperazine (UGT1A4), 4-nitrophenol (UGT1A6), propofol (UGT1A9), 1-naphthol (UGT1A1, UGT1A6, UGT1A8, and UGT1A9), or morphine (UGT2B7) glucuronosyltransferase activities. As shown in Fig. 5, B and C, the etoposide glucuronosyltransferase activity was significantly correlated with the bilirubin (r = 0.935, p < 0.01) and β-estradiol (r = 0.841, p < 0.01) glucuronosyltransferase activities. Furthermore, the etoposide glucuronosyltransferase activity was significantly (r = 0.800, p < 0.01) correlated with the immunoquantified UGT1A1 protein contents (Fig. 5D). In contrast, the etoposide glucuronosyltransferase activity was not correlated with the trifluoperazine (r = 0.204), propofol (r = −0.075), 4-nitrophenol (r = 0.160), 1-naphthol (r = 0.232), or morphine (r = −0.017) glucuronosyltransferase activities.
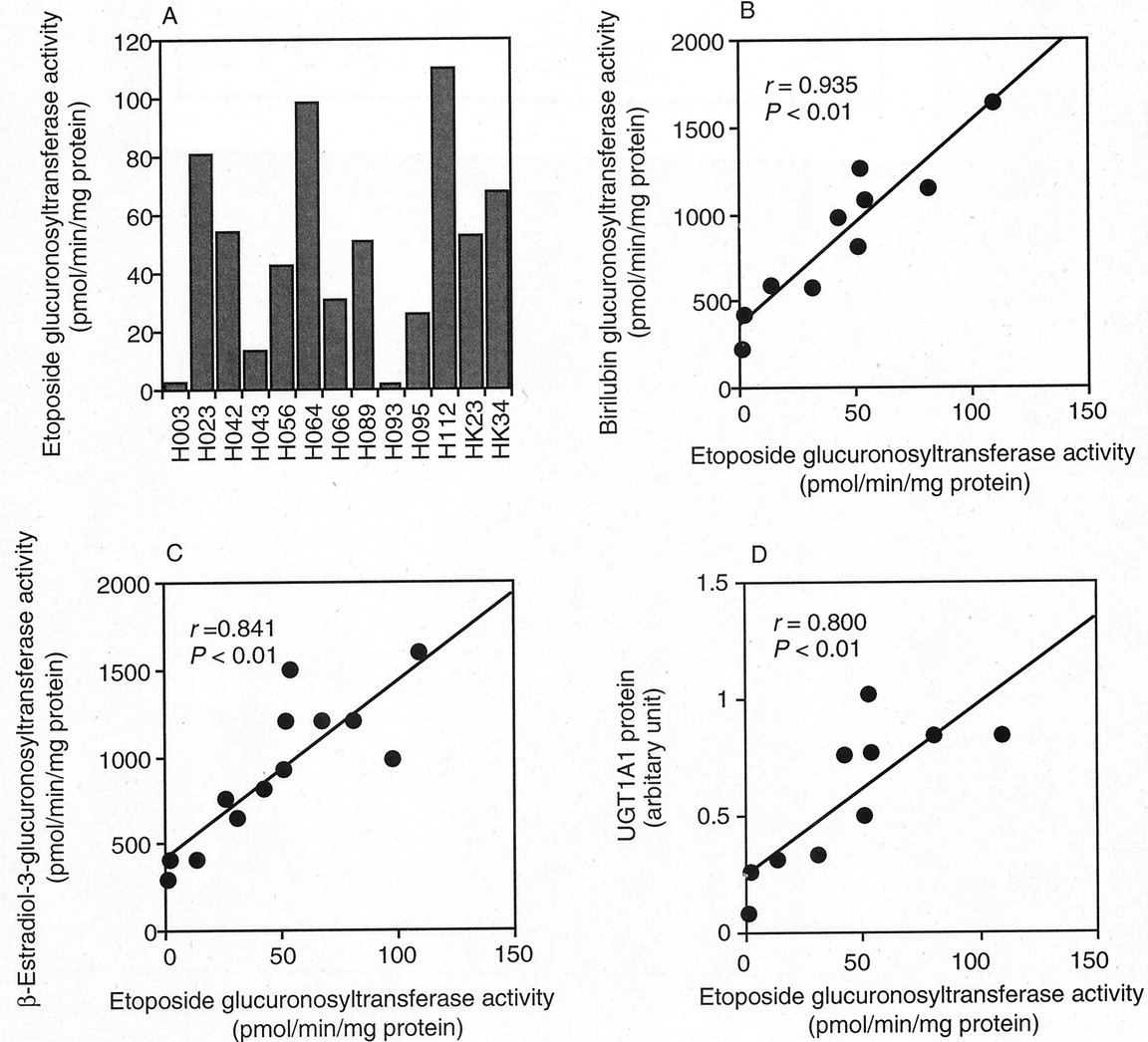
Interindividual variability in etoposide glucuronosyltransferase activity in microsomes from 13 human livers (A) and correlation analyses with enzymatic activities (B and C) and content of UGT1A1 (D).
Etoposide glucuronosyltransferase activities in microsomes from 13 human livers were determined (A). Human liver microsomes (1 mg/ml microsomal protein) were incubated with 400 μM etoposide and 3 mM UDP-glucuronic acid at 37°C for 60 min. Each data point represents the mean of duplicate determinations. Correlation analysis between the etoposide glucuronosyltransferase activity and bilirubin glucuronosyltransferase activity (B, n = 10), β-estradiol-3-glucuronosyltransferase activity (C,n = 13), and the content of UGT1A1 protein (D,n = 10) in human liver microsomes.
Kinetics of Etoposide Glucuronidation in Liver Microsomes from Wistar and Gunn Rats.
Kinetic analyses of etoposide glucuronidation in liver microsomes from Wistar (n = 3) and Gunn (n = 3) rats were performed. The etoposide glucuronide formation in liver microsomes from Gunn rats was not detected at any substrate concentration. In contrast, the kinetics of etoposide glucuronidation in liver microsomes from Wistar rats fitted the Michaelis-Menten kinetics (Fig.6). TheKm value was 565.5 μM andVmax value was 194.7 pmol/min/mg of protein.
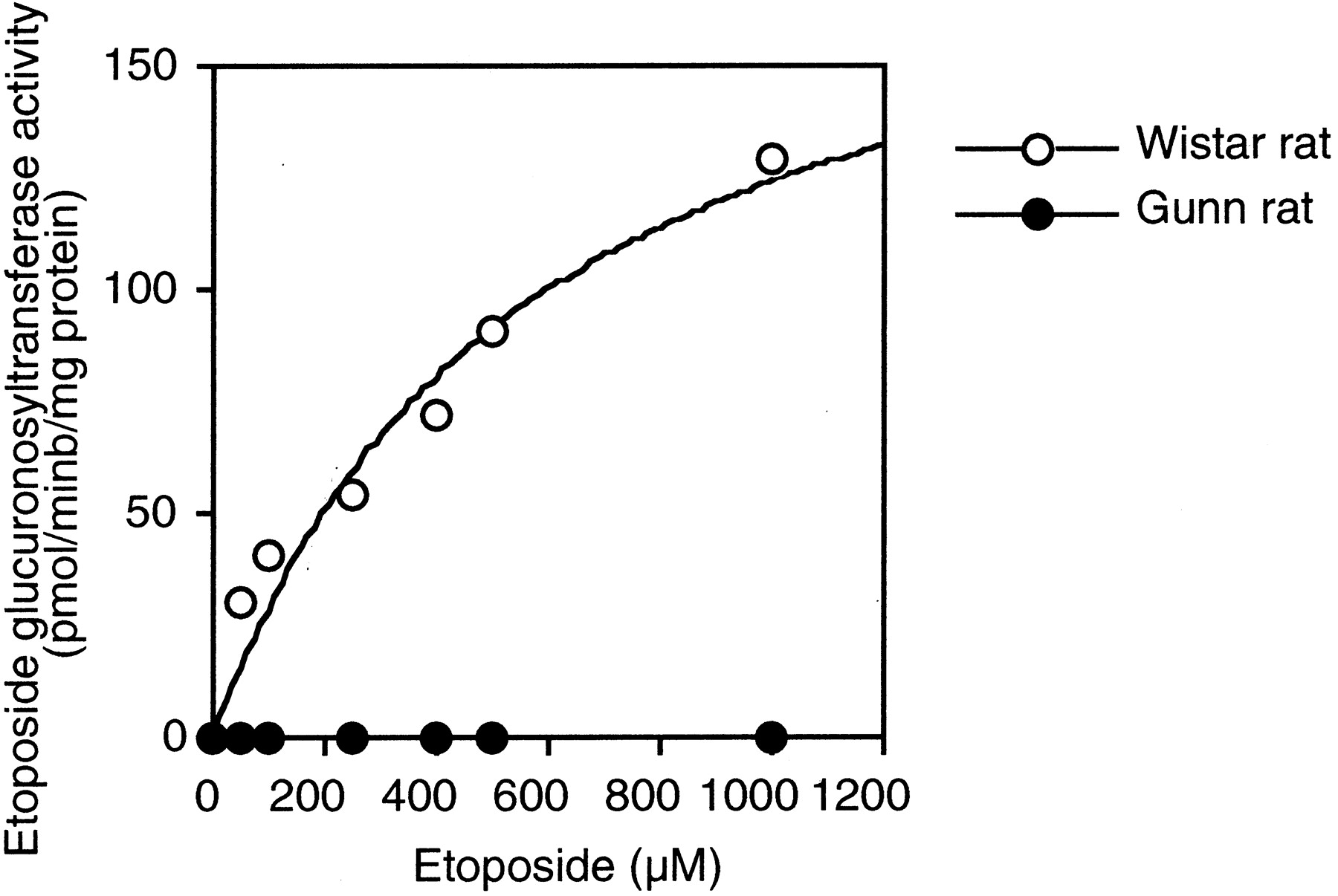
Kinetics of etoposide glucuronidation in liver microsomes from Wistar and Gunn rats.
Plots represents typical data using liver microsomes from one Wistar rat and one Gunn rat. The etoposide concentration ranged from 50 μM to 1 mM. Etoposide glucuronosyltransferase activities were determined as described under Materials and Methods. Each data point represents the mean of duplicate determinations.
Discussion
A number of approaches have been developed for the identification of human UGTs involved in the glucuronidation of endogenous and exogenous compounds in vitro (Ritter, 2000; Tukey and Strassburg, 2000). It has been reported that etoposide is metabolized to its glucuronide in rats and humans (D'Incalci et al., 1986; Hande et al., 1988). However, the UGT isoform that catalyzes the etoposide glucuronidation has not been identified. In the present study, we investigate the etoposide glucuronidation in human liver microsomes to identify the UGT isoform. We confirmed that the peak formed by the incubation of etoposide with human liver microsomes and UDP-glucuronic acid was an etoposide glucuronide by the LC-MS/MS analyses. In the structure of etoposide, there are phenolic and alcoholic hydroxyl groups that can be linked to glucuronic acid. Hande et al. (1988)suggested that glucuronic acid is linked to a phenolic hydroxyl group of etoposide, although they did not confirm the structure. In the present study, etoposide or formed metabolite was treated with Tri-Sil-Z that can silylate a hydroxy-group. As the result of the treatment with Tri-Sil-Z, the peak in the mass spectrometric spectrum at m/z of 455 corresponding to the trimethylsilylated podophyllotoxin moiety was the same in etoposide and etoposide glucuronide. Thus, it was confirmed that the formed metabolite is an etoposide glucuronide and that the glucuronic acid is linked to an alcoholic hydroxyl group. These results were consistent with a previous report by Colombo et al. (1985).
The kinetics of etoposide glucuronidation in human liver microsomes was monophasic, suggesting that a single UGT isoform is responsible for the glucuronidation. Furthermore, the Kmvalue of etoposide glucuronidation in human liver microsomes was very close to that in recombinant UGT1A1. In clinical use, the peak plasma concentration of etoposide was reported to be 27 to 114 μg/ml (46–194 μM) (Hande et al., 1984). However, it has been reported that etoposide in rat livers is 5 times higher than in blood (Nakai et al., 1986). It might be suggested that the concentration of etoposide in human liver would reach to the concentration that is near to theKm value (ca. 500 μM). Among recombinant UGT enzymes examined in the present study, only the UGT1A1 isoform exhibited etoposide glucuronosyltransferase activity. The etoposide glucuronosyltransferase activity in pooled human liver microsomes was inhibited by bilirubin and β-estradiol, which are mainly glucuronidated by UGT1A1. As shown in Fig. 4, inhibition by 4-nitrophenol, imipramine, and morphine was also observed. However, it was confirmed that the etoposide glucuronosyltransferase activity in recombinant UGT1A1 was inhibited to 58, 66, and 67% of control by 500 μM 4-nitrophenol, imipramine, and morphine, respectively. Therefore, the observed inhibitory effects of these compounds on the etoposide glucuronosyltransferase activity in human liver microsomes could be due to the inhibition of UGT1A1. The etoposide glucuronosyltransferase activities in 13 human liver microsomes were significantly correlated with the bilirubin and β-estradiol glucuronosyltransferase activities and immunoquantified UGT1A1 protein content. These results suggest that UGT1A1 would specifically catalyze the etoposide glucuronidation in human liver microsomes.
The Gunn rat has been reported to lack the UGT1A enzymes (Gunn, 1938;Iyanagi, 1991; Sato et al., 1991). In the present study, we demonstrated that the etoposide glucuronosyltransferase activity was not detected in liver microsomes from Gunn rats. In contrast, Wistar rats produced the etoposide glucuronide with aKm value that was similar to that in human liver microsomes. It has been reported that human and rat UGT1A1 enzymes have similar substrate specificity (King et al., 1996) with 89% of protein sequence similarity (Mackenzie et al., 1997). Our results suggest that etoposide glucuronidation is also catalyzed by UGT1A in rats.
We first demonstrated that there is a large interindividual difference in etoposide glucuronidation in humans (78.5-fold). UGT1A1 has been reported to be induced by phenobarbital, phenytoin, oltipraz, and 3-methylcholanthrene (Fisher et al., 2001). Human liver microsomes of H112 showed the highest etoposide glucuronosyltransferase activity (Fig. 6A). From the manufacturer's information, the donor had taken phenobarbital. Therefore, UGT1A1 in H112 might be induced by phenobarbital. Furthermore, it is known that there are genetic polymorphisms in UGT1A1 (Tukey and Strassburg, 2000). Several alleles such as UGT1A1*6 (G71R) and UGT1A1*28[(TA)7TAA] of UGT1A1 have been reported to cause a significant reduction of UGT activity toward bilirubin (Yamamoto et al., 1998) and to be associated with Gilbert's syndrome (Bosma et al., 1995; Monaghan et al., 1996). Therefore, the genetic polymorphisms in UGT1A1 might be a one of factors causing the interindividual differences in etoposide glucuronidation in humans, which will be clarified in the near future.
In conclusion, we found that etoposide glucuronidation in using human liver microsomes is specifically catalyzed by UGT1A1. Large interindividual differences in etoposide glucuronidation in human liver microsomes were observed. These results may have important implications for the clinical variability in efficacy and adverse reactions of etoposide.
Acknowledgments
We thank Dr. Kazuta Oguri for providing morphine-3-glucuronide. We acknowledge Brent Bell for reviewing the manuscript.
Footnotes
- Abbreviations used are::
- UGT
- UDP-glucuronosyltransferase
- LC-MS/MS
- liquid chromatography-tandem mass spectrometry
- ESI
- electrospray ionization
- APCI
- atmospheric pressure chemical ionization
- Received November 21, 2002.
- Accepted February 10, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}